苏姜猪A-FABP基因遗传多态性及其与肉质性状的关联分析
2017-03-29朱爱文倪黎刚周春宝朱淑斌卞桂华
朱爱文,倪黎刚,周春宝,朱淑斌,卞桂华
(江苏农牧科技职业学院,江苏泰州 225300)
苏姜猪A-FABP基因遗传多态性及其与肉质性状的关联分析
朱爱文,倪黎刚,周春宝*,朱淑斌,卞桂华
(江苏农牧科技职业学院,江苏泰州 225300)
研究采用PCR- RFLP技术分析苏姜猪、姜曲海猪、长白猪群体A-FABP基因遗传多态性,并用统计学方法分析基因座多态与肉质性状的相关性。结果表明:3个猪种A-FABP基因内含子1均具有Bsm I酶切多态性,分别检测到3种基因型(AA、AB和BB),χ2适合性检验显示,3个猪种A-FABP基因型分布处于Hardy-Weinberg平衡(P>0.05)。多态信息含量(PIC)分析显示,3个品种猪均为中度多态;基因型与肉质性状关联分析显示,不同基因型肌内脂肪含量、大理石纹差异显著,大理石纹AA、AB型显著高于BB型(P<0.05),肌内脂肪含量AA显著高于AB、BB型(P<0.05)。结论显示,苏姜猪、姜曲海猪、长白猪A -FABP基因intron 1多态性对肉质性状影响显著。
苏姜猪;A-FABP基因;PCR-RFLP;肉质性状
哺乳动物机体内脂肪细胞型脂肪酸结合蛋白(Adipocyte Fatty Acid-binding Protein,A-FABP)主要在脂肪细胞中参与甘油三酯生成,以增加肌内脂肪(Intramuscular Fat, IMF)含量。研究证实,IMF和肉质呈正相关,影响肉的嫩度、风味和多汁性,肉品2.5%~3%的肌内脂肪含量可获得理想口感[1]。利用分子生物学的手段——分子标记辅助选育(MAS)去寻找控制IMF的基因,并进行品种选育,以期提高肌内脂肪含量,是改善肉质行之有效的方法。
Brockmann等[2]研究发现,A-FABP基因位于含有影响脂肪性状QTL的猪4号染色体上,在脂肪细胞中表达,影响甘油三酯的生成与分解。Gerbens等[3]克隆、测序发现,猪A-FABP基因全长8 144 bp,核苷酸序列包括2 370 bp 的5′-上游区域和1 435 bp的3′-下游区域;含有4个外显子和3个内含子,4个外显子分别编码24、58、34、16个氨基酸,内含子的大小为2 629、840、471 bp,与人、小鼠及大鼠同源性分别为90%、83%、81%。Gerbens等[4]研究表明,杜洛克猪A-FABP多态性与含量IMF存在相关。
本研究以姜曲海猪、苏姜猪(以杜洛克猪、姜曲海猪、枫泾猪为亲本,经7个世代选育,2013年9月经国家畜禽遗传委员会审定为猪新品种)、长白猪为研究对象,采用PCR-RFLP技术对改善猪肉质性状起重要作用的A-FABP基因进行检测、分析,旨在寻找与苏姜猪肉质性状相关的遗传标记,探索改善苏姜猪肉质性状的有效途径,并为苏姜猪种质资源特性及分子标记辅助选择(MAS)研究提供科学依据。
1 材料与方法
1.1 材料
1.1.1 实验材料 苏姜猪220头、姜曲海猪96头、长白猪144头,分别来自姜曲海猪保种场、常州康乐农牧公司。取以上猪种耳组织样约0.5 g/头,置于预置70%乙醇的1.5 mL Eppendorf管,-20℃冻存备用。屠宰不同基因型实验猪只,背最长肌中段最后肋与第一、二腰椎间中心部位取样,测定肉质性状指标。
1.1.2 主要试剂 Taq DNA 聚合酶、DL-2000 Marker、dNTPs、Bsm I限制性内切酶等,分别购自宝生物(大连)工程有限公司、上海生工生物技术有限公司、赛默飞世尔科技(中国)有限公司。
1.2 方法
1.2.1 DNA提取 参照《分子克隆实验指南》用酚-氯仿抽提法提取基因组DNA,溶于TE缓冲液[10 mmol/L Tris-HCl(pH 8.0),1 mmol/L EDTA(pH 8.0)],4℃保存[5]。0.8%琼脂糖凝胶电泳法检测抽提基因组DNA质量。
1.2.2 引物设计和 PCR扩增体系 PCR引物根据GenBank发布的猪A-FABP基因DNA序列(登录号:Y16039),参考朱弘焱等[6]发表的引物设计,由上海生工生物技术有限公司合成。所设计引物扩增片段大小为783 bp。引物序列见表1。
基因型频率=基因型个体数/测定群体总数。χ2适合性检验法分析基因型是否处于Hardy-Weinberg平衡状态。基因纯合度、杂合度、有效等位基因数和多态信息含量采用PopGene 32统计软件分析。基因型与肉质性状关联性采用 SPSS(16.0版本)的GLM进行最小二乘方差分析,比较不同基因型之间肉质性状差异,统计模型:
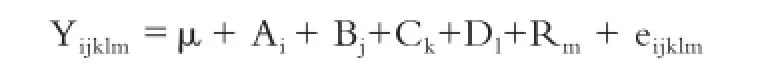
其中,Yijklm为IMF测量值,μ为群体平均值,Ai为场效应,Bj品种效应,Ck为年龄效应,Dl为性别效应,Rm
为基因型效应,eijkl为随机残差效应。
2 结果与分析
2.1 PCR扩增 所设引物用于 PCR 扩增,结果获得了特异性很好的产物(图1),用 1%琼脂糖凝胶电泳检测,片段长度与预期大小一致为783 bp,无非特异性扩增条带,可直接用于RFLP分析。

表1 PCR扩增的引物序列
PCR扩增反应总体系为20 μL:100 ng/μL DNA模板1.0 μL;10×PCR缓冲液2.0 μL;10 μmol/L引物1.0 μL; dNTPs终浓度为200 μmol/L;TaqDNA聚合酶1.0 U;其余用高压灭菌ddH2O补齐。所设计引物的 PCR扩增条件:95℃ 预变性10 min,94℃变性30 s,57℃退火30 s,72℃延伸60 s ,共32个循环,72℃后延伸10 min ,4℃ 保存。扩增产物用1%琼脂糖凝胶电泳检测。
1.2.3 RFLP分析 用BsmI酶切扩增产物。酶切体系20 μL:PCR扩增产物15 μL,10×Buffer 1 μL,限制性内切酶1 μL,灭菌蒸馏水3 μL,37℃ 反应6~8 h。酶切产物用1%琼脂糖凝胶电泳分析,凝胶成像系统拍照分析。
1.2.4 肉质性状测定 测定主要指标包括肉色、大理石纹、pH、失水率、剪切力和IMF含量等。用索氏抽提法提取IMF。测定方法参照相关文献[7-8]。
1.2.5 统计分析 基因频率计算公式:
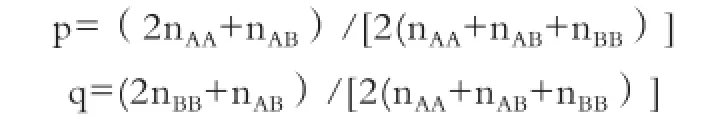
其中,p是等位基因A的基因频率,q是等位基因B的基因频率,nAA、nAB、nBB分别是群体中基因型AA、AB和BB型的个体数。
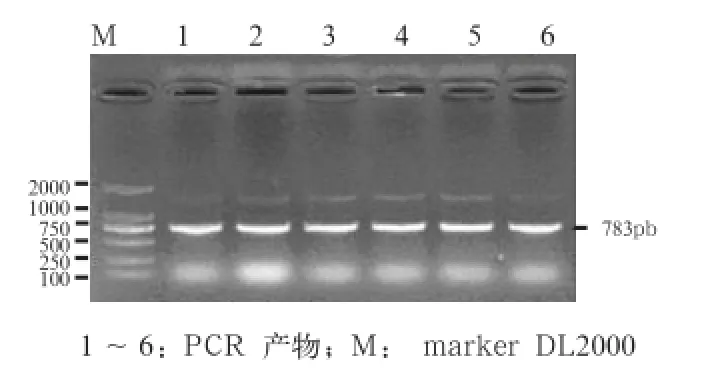
图1 猪A-FABP基因PCR 扩增结果
2.2 RFLP 检测 对扩增的PCR产物进行BsmIRFLP分析,结果发现,引物扩增的目的片段存在多态性,出现3种基因型(图2)。
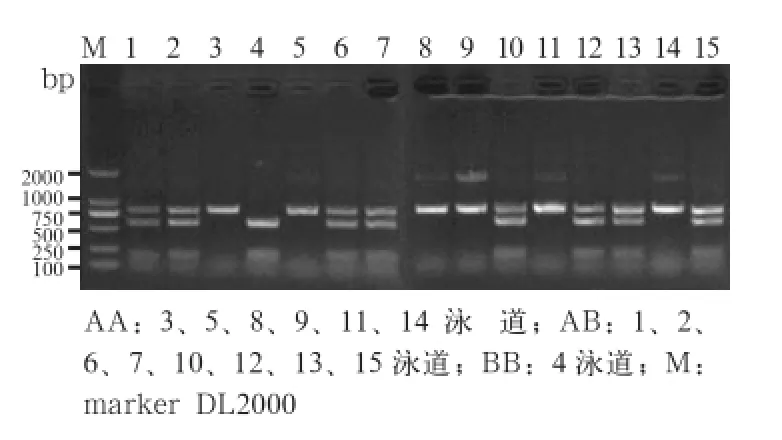
图2 猪A-FABP基因PCR 扩增产物BsmI-RFLP 分析
2.3 不同猪品种A-FABP基因遗传多态性分析
2.3.1 等位基因频率和基因型频率 对3个猪种
A-FABP基因PCR产物进行基因型检测,计算3个猪品种的等位基因频率和基因型频率。表2中χ2适合性检验结果显示,A-FABP基因PCR产物在3个猪种中基因型分布处于Hardy-Weinberg平衡(P>0.05)。
2.3.2 群体遗传杂合度、纯合度、有效等位基因数及多态信息含量分析 PIC用于对标记基因多态性的估计,是表示DNA变异程度高低的指标。PIC<0.25为低度多态,0.25<PIC<0.50为中度多态,PIC >0.50为高度多态。He是衡量群体内遗传变异的有效指标。He越大,群体内遗传变异程度越大。由表3可知,3个猪种均表现为中度多态;苏姜猪遗传多样性比姜曲海猪高。
3 讨 论
3.1 猪A-FABP基因多态性 于小燕等[9]研究发现,松辽黑猪群体存在A-FABP基因5′UTR区域位点多态性,测得3种基因型;李祯等[10]利用微卫星技术研究分析11个猪种A-FABP基因的遗传变异,结果发现微卫星基因座位点高度多态;姜延志等[11]利用PCR-RFLP标记技术检测中国地方猪种雅南猪、大河猪、大河乌猪,研究发现A-FABP基因内含子 1区域BsmI酶切位点具有多态性,发现2个等位基因,3种基因型;张保军等[12]在8个猪种的A-FABP基因内含子1的微卫星位点上,发现16个等位基因,9种基因型。郭瑞等[13]研究猪A-FABP基因的多态性及其与背膘厚的关系,发现民猪、长白猪等群体T65A位点的基因型和基因频率分布存在显著差异,突变位点对背膘厚存在显著的影响,背膘厚度AA型猪显著低于AB型。朱淑斌等[14]研究发现,地方猪种姜曲海猪在BsmI-RFLP位点上存在多态性。本研究在苏姜猪、姜曲海猪、长白猪3个猪种均发现BsmI-RFLP多态性,测得2个等位基因,3种基因型,与姜延志等[11]、朱淑斌等[14]结果相同。
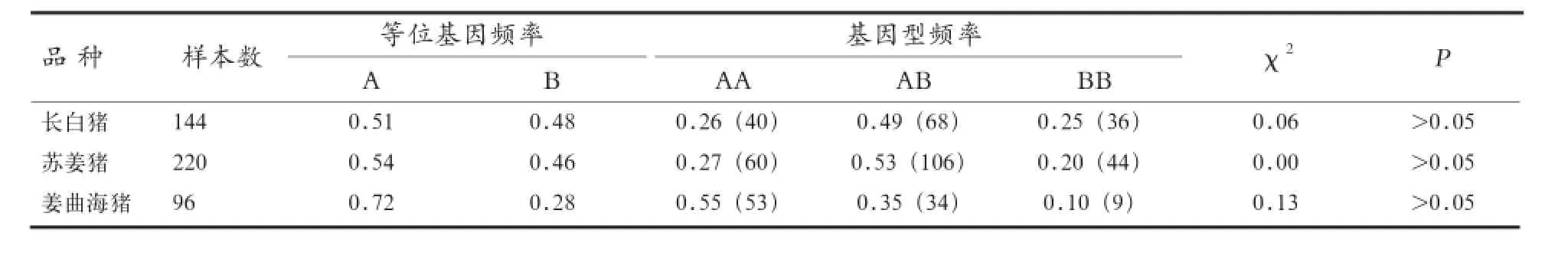
表2 A-FABP基因在 3个猪品种中的等位基因频率和基因型频率

表3 A-FABP基因多态位点纯合度、杂合度、有效等位基因数和多态信息含量
2.4 不同猪种A-FABP基因酶切不同基因型对肉质性状的影响 由表4可知,苏姜猪、姜曲海猪和长白猪A-FABP基因内含子1BsmI酶切后,3种基因型对大理石纹和IMF含量均有显著影响(P<0.05),大理石纹为AA型和AB型均显著高于BB型(P<0.05),AA型和AB型之间差异不显著(P>0.05);IMF含量为AA型显著高于AB型和BB型(P<0.05)),AB型和BB型差异不显著(P>0.05);其他肉质性状的不同基因型之间差异均不显著(P>0.05)。
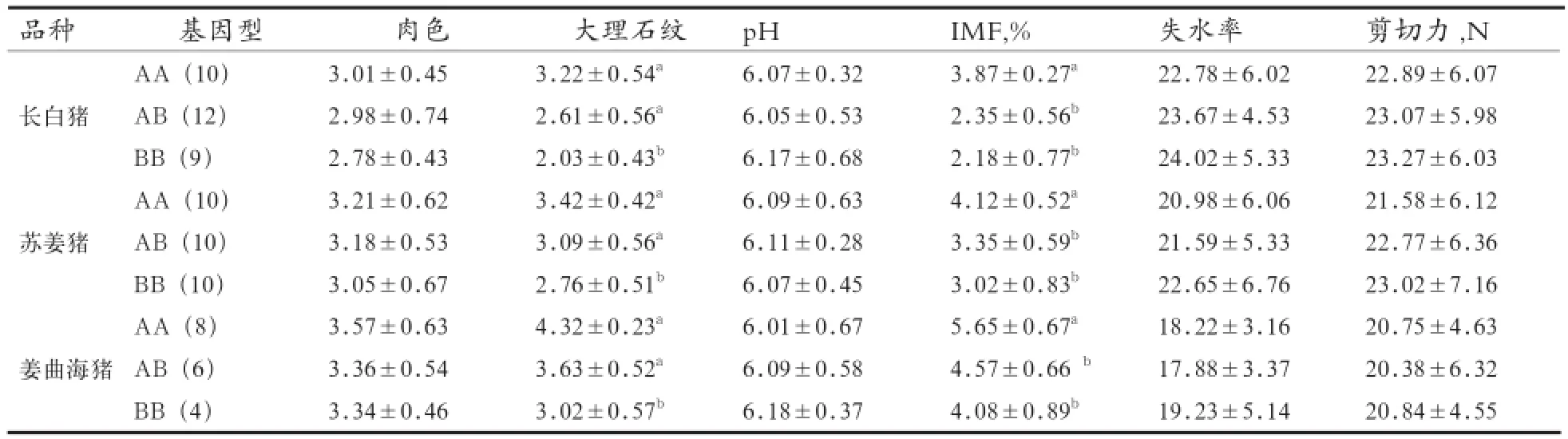
表4 A-FABP基因不同酶切基因型肉质性状比较
3.2A-FABP基因与猪肉质性状的关系 许多学者对猪A-FABP基因与肉质性状的关联进行了分析研究。Andersson等[15]认为在野猪与大白猪杂交F2代群体中第4号染色体上存在影响脂肪沉积的QTL,与Walling等[16]对梅山猪与大白猪杂交F2代群体研究结论相似。姜延志等[9]研究发现,地方猪种大河猪、雅南猪等猪种A-FABP基因内含子1区域存在BsmI位点多态性,IMF含量AA基因型个体极显著高于AB、BB型。Gerbens等[4]在猪A-FABP基因内含子1区内检测到一个(CA)n微卫星序列,杜洛克群体分析发现6种基因型,且AlA3较AlAl基因型IMF含量显著提高。王存芳等[17]研究表明,杜洛克、汉普夏、长白猪、大约克和莱芜猪A-FABP基因微卫星(CA)n位点与IMF含量密切相关。于小燕等[7]在松辽黑猪群体中发现A-FABP基因内含子1位点多态性,不同基因型对大理石纹、IMF含量有极显著影响,而对其他肉质性状影响不显著。本研究中,3 个猪种不同A-FABP基因型在大理石纹和IMF含量2个肉质性状指标中存在显著差异,最小二乘均值关系为AA > AB> BB,试验结果与姜延志等[11]、于小燕等[9]相似。
3.3A-FABP基因内含子1区域BsmI位点AA基因型遗传力 经过对7窝89头仔猪的实验室检测,并结合实验猪场生产数据资料,发现苏姜猪A-FABP基因内含子1区域BsmI多态位点AA基因型公猪和AA基因型母猪交配,后代仔猪均为AA型,暂未出现分离或突变现象,该结果可能与检测样品数量偏少有关,是否会出现分离或变异需进一步检测、研究。
4 结 论
本研究结果表明,苏姜猪、姜曲海猪、长白猪3个猪种A-FABP基因内含子1区域存在BsmI位点多态性,检测到2个等位基因,3种基因型(AA、AB、BB),A为优势等位基因。不同基因型与肉质性状关联分析发现,不同基因型大理石纹和IMF含量差异显著,大理石纹最小二乘均值关系为AA、AB型显著高于BB型,IMF含量最小二乘均值关系为AA型显著高于AB、BB型,其他肉质性状的不同基因型差异均不显著。笔者认为,该突变位点值得引起关注,最终定论还需要扩大样本数、增加猪品种数、对标记与肉质性能关联作进一步深入的研究。
[1] Wood J D, Richardson R Ⅰ, Nute G R,et al. Ef f ects of fatty acids on meat quality:a review[J]. Meat Sci, 2003, 66: 21-32.
[2] Brockmann G,Timtchenko D, Das P,et al. Detection of QTL for body weight and body fat content in mice using genetic markers [J]. Anim Breed Genet, 1996, 113:373-379.
[3] Gerbens F, Harders F L,Groenen MA,et al. A dimorphic microsatellite in the porcine H-FABP gene at chromosome 6 [J].Anim Genet,1998, 29(5):398-413.
[4] Gerbens F,Jansen A,van Erp A P,et al. The adipocyte fatty-acid-binding-protein locus:Characterization, and association with intramuscular fat content in pig[J]. Mamm Genome, 1998, 9(12):1022-1026.
[5] 萨姆布鲁克J, 弗里奇E F, 曼尼阿蒂斯. 分子克隆实验指南[M]. 北京:科学出版社, 1999.
[6] 朱弘焱,苏玉虹,宋衡元. 辽宁种猪H-FABP和A-FABP基因位点多态性研究[J]. 畜牧与兽医, 2010, (8):15-18.
[7] 张伟力,曾勇庆. 猪肉肌内脂肪测定方法及其误差分析[J].猪业科学,2008 , (7) :102-103.
[8] 周贵,王立克,黄瑞华,等. 畜禽生产学实验教程[M]. 北京:中国农业大学出版社,2006:117-180.
[9] 于小燕. 松辽黑猪A-FABP基因SNPs位点及其与肉质性状的相关分析[D]. 长春:吉林大学, 2008.
[10] 李祯,储明星,曹红鹤,等. 中外11个猪种A-FABP基因微卫星遗传变异的研究[J]. 遗传,2004, 26(4): 473-477.
[11] 姜延志,李学伟. 猪脂肪细胞型脂肪酸结合蛋白基因BsmⅠ位点多态性与肌内脂肪含量的相关研究[J].中国畜牧杂志, 2006, 42(7):1-3.
[12] 张保军. 猪LPL基因和A-FABP基因遗传变异研究[D].杨凌:西北农林科技大学, 2004.
[13] 郭瑞. 民猪,长白,大白H-FABP和A-FABP基因多态性的研究[D]. 哈尔滨:东北农业大学,2006.
[14] 朱淑斌,赵旭庭,周春宝,等. 姜曲海猪A -FABP基因内含子1多态性与肉质性状的相关分析[J].中国畜牧杂志,2013,49(13):14-15.
[15] Andersson L, Haley C S, Ellegren H,et al. Genetic mapping of quantitative trait loci for growth and fatness in pigs[J]. Science, 1994, 263:1771-1774.
[16] Walling G A,Archibald A L, Cattermole J A,et al. Mapping of quantitative trait loci on porcine chromosome 4 [J]. Anim Genet, 1998, 29(6): 415-424.
[17] 王存芳. 猪H-FABP和A-FABP基因的多态性及其与肌内脂肪性状关系的研究[D]. 泰安:山东农业大学, 2002.
Genetic Polymorphism of A-FABP Gene and Correlation Analysis with Meat Quality Traits in Sujiang Swine
ZHU Ai-wen, NⅠ Li-gang, ZHOU Chun-bao*, ZHU Shu-bin, BⅠAN Gui-hua
(Jiangsu Agri-animal Husbandry vocation College, Jiangsu Taizhou 225300, China)
Aesearch by PCR-RFLP technical analyzed Sujiang swine, Jingquhai swine and landrace groups’A-FABPgenetic polymorphisms, and analyzed the correlation between loci polymorphism and meat quality traits. Results showed that the three swine-breeds’ A - FABP gene intron 1 haveBsmⅠenzyme polymorphisms, three genotypes were detected (AA, AB, and BB). The Chi-square test results showed that three swine-breeds’A-FABPgenotype distribution in Hardy-Weinberg equilibrium (P>0.05) .Polymorphism information content (PⅠC), which were moderately polymorphic in three varieties of swines.According to the association analysis of different genotypes of marbling and intramuscular fat content(ⅠMF), signif i cant dif f erence was found in marbling, genotype AA, AB was signif i cantly higher than BB (P<0.05), intramuscular fat content genotype AA was signif i cantly higher than AB, BB (P<0.05). Conclusion showed that Sujiang swine, Jiangquhai swine and landrace groups’A-FABPgene intron 1 polymorphism had signif i cant ef f ects on meat quality traits.
Sujiang swine;A-FABPgene; PCR-RFLP; Meat quality traits
S828.2
A
10.19556/j.0258-7033.2017-03-029
2016-07-25;
2016-08-08
江苏省科技项目(BY2014124);江苏省农业三新工程项目(SXGC[2014]271);江苏高校品牌专业建设工程资助项目(PPZY2015A083)
朱爱文(1972-),男,硕士,讲师,主要从事动物遗传育种研究,E-mail:ZAW_0@163.com
* 通讯作者:周春宝(1973-),男,博士,教授,主要从事动物遗传育种研究,E-mial:zhou_tz@ 163.com
