全谷物调节代谢性疾病机制研究的新视角
——肠道微生物
2017-03-08龚凌霄曹文燕张慧娟刘英丽
龚凌霄,曹文燕,王 静,张慧娟,刘英丽
(北京工商大学,北京食品营养与人类健康高精尖创新中心,食品添加剂与配料北京高校工程研究中心,北京 100048)
全谷物调节代谢性疾病机制研究的新视角
——肠道微生物
龚凌霄,曹文燕,王 静*,张慧娟,刘英丽
(北京工商大学,北京食品营养与人类健康高精尖创新中心,食品添加剂与配料北京高校工程研究中心,北京 100048)
全谷物中含有丰富的植物化学素和多种营养成分,增加全谷物的摄入能够显著降低代谢性疾病(如肥胖、Ⅱ型糖尿病)的发生率,然而也正是由于全谷物成分复杂,很多现有的体外研究模型和方法并不适用于全谷物保健机制的研究。随着肠道微生物与人体健康之间的关系逐步得到揭示,肠道微生物启发了全谷物健康保健机制的研究方向。本文将详细介绍肠道微生物与代谢性疾病、全谷物成分与肠道微生物以及全谷物与肠道微生物之间的研究现状,并分析以肠道微生物作为全谷物保健机制研究靶点的可行性,目的在于为明确健康作用的全谷物及其相关产品的综合开发提供研究思路,使得来源广泛、食用方便、功效显著的全谷物更好地服务于人类健康。
全谷物,肠道微生物,代谢性疾病,肥胖
谷物作为人类最基本的膳食组成,与人体健康有着密切关系。其中,由于全谷物中各种营养成分与植物化学素(包括酚酸、黄酮、类胡萝卜素、维生素E、γ-谷维素和β-葡聚糖、阿拉伯木聚糖等)保持原生状态、并存在协同增效作用,因此与摄入精制的谷物及提纯的植物化学素相比,增加全谷物的摄入更有利于人体健康,如降低心血管疾病、糖尿病及某些癌症等慢性疾病发生的风险等[1]。此外,新版《中国居民膳食指南(2016版)》首次推荐了每天的全谷物摄入量,强调了全谷物在我国居民健康饮食中的重要性。然而,由于全谷物成分复杂,在体内消化情况非常复杂,很多现有的体外研究模型和方法并不适用于全谷物保健机制的研究,迄今有关全谷物营养保健作用的研究主要是基于人群调查,以观察性为主,或是对全谷物中少数具体的健康因子进行健康作用的探讨,因此全谷物健康保健机制的研究一直是该领域的难点。
随着分子生物学技术的迅猛发展,作为人类“第二基因组”的肠道微生物与人体健康的关系正逐步得到揭示。当外界环境变化导致肠道微生物结构发生变化时,肠道菌群能够通过多种途径调控宿主的生理代谢。目前,越来越多的研究者认识到了肠道微生物结构的变化和功能的异常与很多代谢性疾病的发生发展密切相关。众所周知,作为含有多种营养物质和植物化学素的“全营养包”的全谷物经人体胃肠消化后进入下消化道中的部分可被肠道微生物利用,造成肠道微生物结构的变化从而对机体进行生理调节[2]。因此,肠道微生物逐渐成为全谷物对人体健康作用机制的研究靶点。本文将对全谷物介导肠道微生物预防代谢性疾病的研究现状进行综述,为利用全谷物更好地服务于人类健康提供更广阔的研究思路。
1 肠道微生物及其对宿主的影响和作用机制
定植于人类肠道中的微生物其结构和功能非常多样和复杂,包含有500~1000种菌,总数达到1013~1014,是人自身细胞总数的10倍,而其包含的基因数目是人基因组的100倍[3]。人体肠道内的菌群主要由9个门的细菌和一种古菌(Methanobrevibactersmithii)组成,9个门的细菌分别为:厚壁菌(Firmicutes)、拟杆菌(Bacteroidetes)、放线菌(Actinobacteria)、梭杆菌(Fusobacteria)、变形菌(Proteobacteria)、蓝藻菌(Cyanobacteria)、Verrucomicrobia、Spirochaeates和VadinBE97。其中98%的菌群可归入革兰氏阳性的厚壁菌(64%)和放线菌(3%)、革兰氏阴性的拟杆菌(23%)和变形菌(8%)。肠道微生物会产生很多复杂的代谢物和分子信号来影响人体健康,若肠道中有益菌占优势,则对净化肠道、维护人体健康起着重要作用;若有害菌占优势,则会导致很多疾病的发生[4]。图1总结了肠道微生物导致宿主代谢疾病发生的主要途径。
1.1 增强食欲,增加能量摄入
肠道细菌发酵不被小肠吸收的食物残渣和上皮细胞分泌的内生黏液,发酵后的代谢产物(如短链脂肪酸(Short Chain Fatty Acids,SCFAs)、葡萄糖等)是能被宿主吸收和利用的能量和底物,使宿主获得更多能量,同时为微生物自身的生长和增殖提供能量和营养物质[5];短链脂肪酸能够激活游离脂肪酸受体(Free Fatty Acid Receptors,FFARs),而FFARs能够减少控制食欲的激素酪酪肽(PYY)的分泌来增加食欲,同时促进脂肪组织中甘油三酯的存储,导致脂肪积累[6-7]。
1.2 调节宿主脂肪存储的相关基因和代谢通路,使宿主增加脂肪的积累
肠道菌群通过调控禁食诱导脂肪细胞因子(Fasting-induced adipocyte factor,Fiaf)的表达,诱导过氧化物酶体增殖激活受体辅助激活因子(Pgc-1α),降低5’-腺嘌呤核苷酸活化蛋白激酶(AMPK)活性来影响细胞能量代谢,增加脂蛋白脂肪酶(Lipoprotein Lipase,LPL)活性来影响脂肪代谢等;另一方面,短链脂肪酸直接刺激肝脏中脂肪代谢相关的酶,从而促进肝脏脂肪合成[8-9]。
1.3 引发慢性炎症反应,产生肥胖、胰岛素抵抗等代谢失调症状
革兰氏阴性菌产生的细菌脂多糖(lipopolysaccharide,LPS,又称细菌内毒素)通过破坏肠道黏膜进入血液,与机体免疫巨噬细胞的表面蛋白受体CD14/TLR4结合,通过核因子-κB(nuclear factor-κB,NF-κB)和过氧化物酶体增殖物激活受体(peroxisome proliferator-activated receptor,PPARγ)途径促进炎症因子的分泌,引发慢性炎症,从而诱导肥胖的发生[10-11]。
随着肠道菌群与代谢性疾病发生之间的关系研究的不断深入,肠道菌群逐步成为预防、缓解,甚至逆转代谢性疾病的新靶点。
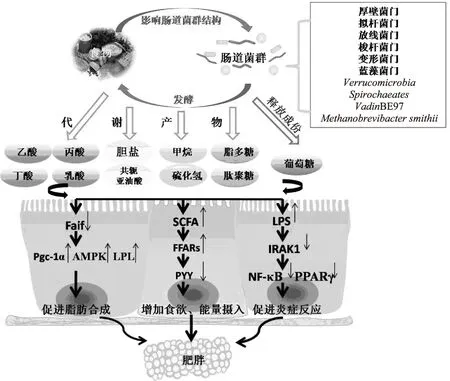
图1 肠道微生物导致肥胖和相关代谢疾病发生的主要途径Fig.1 Gut microbiota involved in the development of obesity and metabolic syndrome
2 全谷物中主要生理活性物质对肠道微生物的影响
近年来越来越多的研究证明,人体和动物直接摄入全谷物后,能够有效促进体内肠道菌群中双歧杆菌和乳酸杆菌的增殖[12-13]。目前,从谷物中已发现多种活性物质具有改善肠道微生物结构的作用。
2.1 膳食纤维
全谷物中的膳食纤维主要为细胞壁多糖(主要为纤维素、β-葡聚糖和阿拉伯木聚糖)和抗性淀粉,其中阿拉伯木聚糖通常与阿魏酸或对香豆酸等以酯键相连,因此形成另一类重要的功能性低聚糖类物质(酚酸糖酯类),上述成分均在改善肠道微生物结构中扮演着十分重要的角色。膳食纤维不被人体的消化酶水解,但可被肠道微生物中特定的菌所分泌的酶水解,膳食纤维水解产物可为特定的肠道菌提供充足的碳源而被发酵利用,最终产生能量和乙酸,丙酸和丁酸等短链脂肪酸[14]。这些短链脂肪酸既能被宿主快速吸收利用,也能被肠道双歧杆菌和乳杆菌等有益菌利用,一方面促进有益菌增殖,另一方面通过降低肠道pH抑制大肠杆菌和沙门氏菌等病原菌的生长、减少有毒发酵产物的形成,进而改善肠道菌群结构[15-16]。
Neyrinck[17]等观察到小麦AX可以恢复高脂饮食引起的小鼠的肠道细菌移位,增加双歧杆菌数量,尤其是乳双歧杆菌的数量。Vardakou等[18]的体外研究结果显示了小麦AX抑制类杆菌和梭杆菌生长的作用。AX的肠道益生活性与其结构密切相关,不同来源的AX显示出不同的益生效果。利用碱水解法分别从玉米、大米和小麦麸皮中制备可溶性AX,然后用从人的粪便中分离的微生物进行发酵,结果显示玉米AX在肠道中发酵后生成的SCFA产量最高[19]。Van den Abbeele等[20]比较了AX和菊粉的体外发酵情况,结果发现长链阿拉伯木聚糖能够增加长双歧杆菌的数量和丙酸盐的含量。更值得注意的是,长链阿拉伯木聚糖可以在末端结肠中被发酵,菊粉则只能在升结肠中被发酵,而末端结肠中的菌群结构对结肠健康具有重要意义。此外,AX的水解产物阿拉伯木聚寡糖(arabinoxylanoligosaccharides,AXOS)也具有益生作用,与AX相似,AXOS发酵后促进乙酸、丙酸盐、正丁酸盐的生成。Geraylou等[21]发现含有2%AXOS的饲料可以显著促进鱼肠道中的乳酸杆菌和梭状芽孢杆菌的生长。其中,较高聚合度的AXOS(DP为32)比低聚合度的AXOS(DP为3)对上述菌具有更强的增殖作用。Neyrinck等[22]发现与单独饲喂高脂饲料的小鼠相比,在高脂饲料中添加7.5%的小麦麸皮AXOS后,小鼠粪便中双歧杆菌显著增加,与双歧杆菌密切相关的抗炎症因子(IL10)的表达上调,而脂肪组织中的炎症因子IL6和F4/80的表达受抑制。与此同时,小鼠血液中抑制食欲的PPY激素和胰高血糖素样肽-1含量显著增加,小鼠的体重和脂肪质量显著减少。
β-葡聚糖具有显著的降脂减肥功效,越来越多的研究表明β-葡聚糖的生理活性与其益生元作用密切关联[23]。Metzler-Zebeli等[24]对26篇文献中涉及的小麦、燕麦、大麦、玉米和黑麦5种谷物来源的β-葡聚糖对猪消化情况进行Meta分析后得出,β-葡聚糖的摄入可以增加猪粪便中的总酸、大肠中的异丁酸盐、回肠中的丁酸盐的含量,减少粪氮排出,改善消化状况。大麦β-葡聚糖能够增加健康老年人肠道中双歧杆菌和类杆菌的数量[25]。值得注意的是,β-葡聚糖调节肠道菌群的效果与其分子大小有关。Hughes等[26]分别选用不同分子量大小的大麦β-葡聚糖(243、172、137 ku)和燕麦β-葡聚糖(230、150 ku)进行人体粪便接种体外发酵,结果显示,所有β-葡聚糖能增加发酵液中丙酸盐、丁酸盐的含量,但增加程度各有不同。分析菌群后发现,溶组织梭菌(C.histolyticum)数量明显增殖,而该菌群可生成β-葡聚糖内切酶而将β-葡聚糖作为碳源利用。Wang等[27]的临床双盲实验显示,正常人连续4周每天摄入3 g大分子量大麦β-葡聚糖(1349 ku)后,粪便中拟杆菌门数量显著增加,厚壁菌门数量显著减少,但摄入小分子量大麦β-葡聚糖(292 ku)则对肠道菌群没有显著影响。
2.2 抗性淀粉
抗性淀粉(Rsistant starch,RS)因其不能在健康人体小肠中被消化吸收而能被肠道菌群发酵利用,因此具有与膳食纤维相似的益生功能[28-29]。方建东[30]的研究显示,高直链玉米抗性淀粉能够降低成年KM小鼠结肠内pH环境,增加乙酸、丙酸、丁酸含量,同时明显促进乳酸杆菌和双歧杆菌增殖,抑制大肠杆菌和肠球菌生长,而且上述调整效果呈现一定的剂量依赖性。但是由于高含量RS使结肠内pH过低,会出现正常菌群的增殖被抑制的现象。Dahl等[31]研究显示连续6周分别饮用含30 g不同RS4型的土豆抗性淀粉的饮料后,健康成人粪便中的双歧杆菌数量并没有变化,但粪便中厚壁菌门数量、厚壁菌门与拟杆菌比值减少,说明抗性淀粉造成健康成人肠道内的菌群漂移。Walker等[32]的人体内研究也表明,RS的摄入能够显著提高布氏瘤胃球菌属(Ruminococcusbromii)、颤杆菌克属(Oscillibacter)和直肠真杆菌属(Eubacteriumrectale)数量。
2.3 酚类物质
谷物中含有多种酚类化合物,如阿魏酸、香草酸、咖啡酸、丁香酸、对香豆酸、花青素、奎宁、黄酮、黄酮醇、黄烷酮类和氨基酚类化合物[33]。谷物中的酚类物质以游离态、可溶性结合型和不溶性结合型三种形式存在,80%以上存在于麸皮和胚乳中[34]。游离态酚类物质主要在小肠中被吸收,而结合态酚类物质则在结肠中被肠道微生物分泌的多酚代谢酶水解、转化后才被人体吸收代谢,因此酚类物质与肠道菌群之间有复杂的相互关系[35-36]。体外研究表明[37],花青素、绿原酸、咖啡酸、芦丁和槲皮素等都能促进人粪便培养液中双歧杆菌等益生菌的生长,增加短链脂肪酸的含量,抑制大肠杆菌、鼠李糖乳杆菌、鼠伤感沙门氏菌等有害菌的生长,减少厚壁菌门和拟杆菌门的比例[38-40]。酚类物质可通过破坏特定菌细胞膜的正常功能、改变微生物代谢酶构成和活性等途径来影响肠道菌群结构,但尚不清楚引起这种改变是由于酚类物质本身还是酚类物质的代谢产物。另外,阿魏酸是谷物酚类物质的主要成分,但99%以上的阿魏酸与阿拉伯糖以阿魏酰糖酯的形式存在,结肠中微生物所产生的阿魏酰酯酶能够水解获得游离的阿魏酸和阿魏酰低聚糖。阿魏酰低聚糖被证明具有增殖双歧杆菌、改善肠道微生物菌群的作用,其效果优于低聚木糖[41]
3 全谷物对肠道微生物的影响
全谷物与精制谷物相比,其对益生菌的增殖作用关键在于它的麸皮。谷物麸皮中富集的阿拉伯木聚糖、β-葡聚糖和纤维素等膳食纤维以及多种植物化学素。Benno等[42]检测了食用糙米和大米的志愿者的肠道中梭状芽胞杆菌数量变化情况,与食用大米的志愿者相比,食用糙米的志愿者肠道梭状芽胞杆菌数量显著降低。Vitaglione等[43]研究显示,健康成人连续8周食用全麦产品后,血清中二氢阿魏酸和粪便中阿魏酸含量分别增加了4倍和2倍,而食用精制小麦产品的健康成人则没有变化。同时,增加全麦的摄入使得血浆炎症因子TNF-α减少,IL-10增加,使粪便中拟杆菌和厚壁菌门数量增加,而梭菌属数量减少。通过相关性分析显示,粪便阿魏酸含量与类杆菌数量之间呈明显正相关,而TNF-α含量与类杆菌和乳酸杆菌数量则呈负相关。结果说明,肠道菌群中的厚壁菌和拟杆菌共同作用使全麦中结合阿魏酸释放,增加阿魏酸的生物利用率而改善宿主的炎症水平。Lappi等[44]让芬兰的51位年龄在54~66岁的代谢综合征患者连续12周分别摄入全燕麦面包和精制小麦面包,对干预前后各位受试者粪便中的菌群进行分析后研究发现,虽然两组受试者肠道中的微生物组成基本没有变化。但是,摄入全燕麦面包的受试者的粪便中拟杆菌门数量减少37%,另外Bacteroidsvulgatus、B.plebeius和Prevotellatannerae减少,而梭菌Ⅳ群和Ⅺ群增加。赵兰涛等[45]人用5种不同的全谷物和一种精制谷物研究对小鼠肠道菌群的影响,研究发现全谷物对肠道菌群有益生调节作用,摄入全谷物的小鼠肠道双歧杆菌和乳杆菌数量显著上升,而摄入精米和基础饲料则没有变化,表明全谷物对改善肠道环境和控制体重方面有作用。
虽然全谷物的益生作用与其麸皮密切相关,但研究却显示全谷物比麸皮具有更好的益生作用。Costabile等[46]让31名健康成人连续3周每天连续摄入48 g全小麦早餐谷物或小麦麸皮早餐谷物后发现,摄入全小麦早餐谷物组的粪便中双歧杆菌和乳酸菌数量较摄入小麦麸皮组明显增加,表明全谷物比麸皮的益生效果更好。
另外,不同谷物的益生效果也具有明显的差异。Yang等[47]首先将五种谷物(小麦、黑麦、玉米、稻米和燕麦)进行体外消化,然后再将消化液透析后的膳食纤维分别在肥胖成人和正常体重成人的粪便中进行体外接种发酵,结果显示,燕麦发酵后产生的丙酸盐含量最低而丁酸盐含量最高,这与燕麦中水溶性β-葡聚糖含量比其他谷物高相关,表明燕麦具有较好的减肥效果。在猪的研究中表明,在促进结肠中乳酸菌增殖的能力方面,燕麦>大麦>小麦,而且燕麦促进双歧杆菌增殖、减少肠杆菌的能力也较强[48]。Zhong等[49]研究发现,食用全大麦后,低脂和高脂饲养Wistar大鼠粪便中的SCFA含量均显著增加,而且饮食中膳食纤维含量的高低对效果具有显著影响。两种全大麦的摄入降低了血浆中LPS、MCP-1的含量,增加了粪便中乳酸菌(Lactobaillus)数量,减少了粪便中脆弱类杆菌(Bacteroidesfragilis)数量,但是发现仅低脂饲养的大鼠粪便中双歧杆菌属数量增加了,说明全大麦通过改善肠道菌群结构缓解了高脂饮食诱导大鼠的炎症反应。
以上结果均表明,全谷物对肠道菌群的变化和宿主代谢都产生了影响,全谷物对身体有益的机制可能是通过肠道菌群来实现的。但是这种调节作用的机制目前有待更深入的研究。
4 结语
全谷物富含膳食纤维(戊聚糖、β-葡聚糖和纤维素等)和多种活性成分,这些物质不能被小肠消化吸收利用,能够在大肠处被肠道菌群利用使其增殖并产生代谢物,从而改善有益菌的生长环境,促进肠道内有益菌的增殖、改善肠道菌群结构、保护肠屏障功能,促进肠蠕动,减少肠道间有害物质的停留从而对慢性疾病的发生起到预防作用。但是目前已开展的工作主要集中在全谷物对肠道菌群组成和数量的影响上,而针对全谷物和肠道菌群代谢产物及其代谢性疾病发生的分子机制方面的研究几乎没有。肠道微生物作为人体的“第二器官”与人体的健康密切相关,由于肠道微生物可以被饮食所调节,因此可以预防饮食不平衡而导致的代谢性疾病,可介导肠道微生物进行预防和治疗。我国谷物种类丰富,分布广,产量大,是谷物生产优势最强大的国家,全谷物类产品加工程度比较低,保留了谷物中大量的天然成分,但是全谷物产品的口感粗糙,我国对谷物的消费很大还停留在追求精白面粉、大米的优良口感和风味上,对全谷物产品的开发较西方国家滞后,但随着人们对全谷物食品营养价值的认识逐渐加深,全谷物产业会成为我国粮食加工和食品工业的发展方向,全谷物食品的开发具有广阔的应用前景。而将调节肠道微生物以改善和预防代谢性疾病的发生作为全谷物产品开发的新视角,可以促进全谷物食品开发新技术、新产品的研制,益于对我国特色谷物资源的充分挖掘和利用,在进一步推动我国全谷物行业的发展的同时,为改善国民健康提供经济、便捷的饮食保障措施。
[1]Okarter N,Liu R H. Health benefits of whole grain phytochemicals[J]. Critical Reviews in Food Science and Nutrition,2010,50(3):193-208.
[2]Lamsal B P,Faubion J M. The beneficial use of cereal and cereal components in probiotic foods[J]. Food Reviews International,2009,25(2):103-114.
[3]Nicholson J K,Holmes E,Lindon J C,et al. The challenges of modeling mammalian biocomplexity[J]. Nature Biotechnology,2004,22(10):1268-1274.
[4]陈卫. 肠道菌群:膳食与健康研究的新视角[J]. 食品科学技术学报,2015,33(6):1-6.
[5]Roberfroid M B,Bornet F,Bouley C,et al. Colonic microflora:nutrition and health. Summary and conclusions of an International Life Sciences Institute(ILSI)[Europe]workshop held in Barcelona,Spain[J]. Nutrition Reviews,1995,53(5):127-130.
[6]Samuel B S,Shaito A,Motoike T,et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor,Gpr41[J]. Proceedings of the National Academy of Sciences,2008,105(43):16767-16772.
[7]Dewulf E M,Cani P D,Neyrinck A M,et al. Inulin-type fructans with prebiotic properties counteract GPR43 overexpression and PPARγ-related adipogenesis in the white adipose tissue of high-fat diet-fed mice[J]. The Journal of Nutritional Biochemistry,2011,22(8):712-722.
[8]Bäckhed F,Ding H,Wang T,et al. The gut microbiota as an environmental factor that regulates fat storage[J]. Proceedings of the National Academy of Sciences of the United States of America,2004,101(44):15718-15723.
[9]Bäckhed F,Manchester J K,Semenkovich C F,et al. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(3):979-984.
[10]Cani P D,Amar J,Iglesias M A,et al. Metabolic endotoxemia initiates obesity and insulin resistance[J]. Diabetes,2007,56(7):1761-1772.
[11]Cerf-Bensussan N,Gaboriau-Routhiau V. The immune system and the gut microbiota:friends or foes?[J]. Nature Reviews Immunology,2010,10(10):735-744.
[12]赵兰涛,程李琳,张晖,等. 不同谷物对小鼠肠道菌群的影响[J]. 粮食与食品工业,2013,20(5):51-55.
[13]孙元琳,陕方,赵立平. 谷物膳食纤维——戊聚糖与肠道菌群调节研究进展[J]. 食品科学,2012,33(9):326-330.
[14]Tremaroli V,Bäckhed F. Functional interactions between the gut microbiota and host metabolism[J]. Nature,2012,489(7415):242-249.
[15]Hughes S A,Shewry P R,Li L,et al.Invitrofermentation by human fecal microflora of wheat arabinoxylans[J]. Journal of Agricultural and Food Chemistry,2007,55(11):4589-4595.
[16]Cani P D,Possemiers S,Van de Wiele T,et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability[J]. Gut,2009,58(8):1091-1103.
[17]Neyrinck A M,Possemiers S,Druart C,et al. Prebiotic effects of wheat arabinoxylan related to the increase in bifidobacteria,Roseburia and Bacteroides/Prevotella in diet-induced obese mice[J]. PLoS One,2011,6(6):e20944.
[18]Vardakou M,Palop C N,Gasson M,et al.Invitrothree-stage continuous fermentation of wheat arabinoxylan fractions and induction of hydrolase activity by the gut microflora[J]. International Journal of Biological Macromolecules,2007,41(5):584-589.
[19]Rose D J,Patterson J A,Hamaker B R. Structural differences among alkali-soluble arabinoxylans from maize(Zea mays),rice(Oryza sativa),and wheat(Triticum aestivum)brans influence human fecal fermentation profiles[J]. Journal of Agricultural and Food Chemistry,2009,58(1):493-499.
[20]Van den Abbeele P,Venema K,Van de Wiele T,et al. Different human gut models reveal the distinct fermentation patterns of arabinoxylan versus inulin[J]. Journal of Agricultural and Food Chemistry,2013,61(41):9819-9827.
[21]Geraylou Z,Souffreau C,Rurangwa E,et al. Effects of arabinoxylan-oligosaccharides(AXOS)on juvenile Siberian sturgeon(Acipenser baerii)performance,immune responses and gastrointestinal microbial community[J]. Fish & Shellfish Immunology,2012,33(4):718-724.
[22]Neyrinck A M,Van Hee V F,Piront N,et al. Wheat-derived arabinoxylan oligosaccharides with prebiotic effect increase satietogenic gut peptides and reduce metabolic endotoxemia in diet-induced obese mice[J]. Nutrition & Diabetes,2012,2(1):e28.
[23]申瑞玲,朱莹莹,李林,等. 燕麦β-葡聚糖调节肠道菌群与降脂减肥作用的研究进展[J]. 食品工业科技,2014,35(8):364-366.
[24]Metzler-Zebeli B U,Zebeli Q. Cerealβ-glucan alters nutrient digestibility and microbial activity in the intestinal tract of pigs,and lower manure ammonia emission:A meta-analysis[J]. Journal of Animal Science,2013,91(7):3188-3199.
[25]Mitsou E K,Panopoulou N,Turunen K,et al. Prebiotic potential of barley derivedβ-glucan at low intake levels:a randomised,double-blinded,placebo-controlled clinical study[J]. Food Research International,2010,43(4):1086-1092.
[26]Hughes S A,Shewry P R,Gibson G R,et al.Invitrofermentation of oat and barley derivedβ-glucans by human faecal microbiota[J]. FEMS Microbiology Ecology,2008,64(3):482-493.
[27]Wang Y,Ames N P,Tun H M,et al. High molecular weight barleyβ-glucan alters gut microbiota toward reduced cardiovascular disease risk[J]. Frontiers in Microbiology,2016,7(e28472).
[28]Nugent A P. Health properties of resistant starch[J]. Nutrition Bulletin,2005,30(1):27-54.
[29]Topping D L,Fukushima M,Bird A R. Resistant starch as a prebiotic and synbiotic:state of the art[C]//Proceedings-nutrition Society of London. CABI Publishing,1999,62(1):171-176.
[30]方建东.抗性淀粉对小鼠肠道菌群的影响极以及作用机制的研究[D].杭州:浙江工商大学,2014.
[31]Dahl W J,Ford A L,Ukhanova M,et al. Resistant potato starches(type 4 RS)exhibit varying effects on laxation with and without phylum level changes in microbiota:A randomised trial in young adults[J]. Journal of Functional Foods,2016,23:1-11.
[32]Walker A W,Ince J,Duncan S H,et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota[J]. The ISME Journal,2011,5(2):220-230.
[33]Liu R H. Whole grain phytochemicals and health[J]. Journal of Cereal Science,2007,46(3):207-219.
[34]Adom K K,Liu R H. Antioxidant activity of grains[J]. Journal of Agricultural and Food Chemistry,2002,50(21):6182-6187.
[35]Valdés L,Cuervo A,Salazar N,et al. The relationship between phenolic compounds from diet and microbiota:impact on human health[J]. Food & Function,2015,6(8):2424-2439.
[36]杨华,叶发银,赵国华. 膳食多酚与肠道微生物相互作用研究进展[J]. 食品科学,2015(3):223-227.
[37]Zhang X,Yang Y,Wu Z,et al. The Modulatory Effect of Anthocyanins from Purple Sweet Potato on Human Intestinal Microbiotainvitro[J]. Journal of Agricultural and Food Chemistry,2016,64(12):2582-2590.
[38]Parkar S G,Trower T M,Stevenson D E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota[J]. Anaerobe,2013,23:12-19.
[39]Lee H C,Jenner A M,Low C S,et al. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota[J]. Research in Microbiology,2006,157(9):876-884.
[40]Parkar S G,Stevenson D E,Skinner M A. The potential influence of fruit polyphenols on colonic microflora and human gut health[J]. International Journal of Food Microbiology,2008,124(3):295-298.
[41]Yuan X,Wang J,Yao H. Feruloyl oligosaccharides stimulate the growth of Bifidobacterium bifidum[J]. Anaerobe,2005,11(4):225-229.
[42]Benno Y,Endo K,Miyoshi H,et al. Effect of rice fiber on human fecal microflora[J]. Microbiology and Immunology,1989,33(5):435-440.
[43]Vitaglione P,Mennella I,Ferracane R,et al. Whole-grain wheat consumption reduces inflammation in a randomized controlled trial on overweight and obese subjects with unhealthy dietary and lifestyle behaviors:role of polyphenols bound to cereal dietary fiber[J]. The American Journal of Clinical Nutrition,2015,101(2):251-261.
[44]Lappi J,Salojärvi J,Kolehmainen M,et al. Intake of whole-grain and fiber-rich rye bread versus refined wheat bread does not differentiate intestinal microbiota composition in Finnish adults with metabolic syndrome[J]. The Journal of Nutrition,2013,143(5):648-655.
[45]赵兰涛. 全谷物对肠道菌群益生作用的研究[D]. 无锡:江南大学,2013.
[46]Costabile A,Klinder A,Fava F,et al. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota:a double-blind,placebo-controlled,crossover study[J]. British Journal of Nutrition,2008,99(1):110-120.
[47]Yang J,Keshavarzian A,Rose D J. Impact of dietary fiber fermentation from cereal grains on metabolite production by the fecal microbiota from normal weight and obese individuals[J]. Journal of Medicinal Food,2013,16(9):862-867.
[48]Aumiller T,Mosenthin R,Weiss E. Potential of cereal grains and grain legumes in modulating pigs’ intestinal microbiota-A review[J]. Livestock Science,2015,172:16-32.
[49]Zhong Y,Marungruang N,Fk F,et al. Effects of two whole-grain barley varieties on caecal SCFA,gut microbiota and plasma inflammatory markers in rats consuming low-and high-fat diets[J]. British Journal of Nutrition,2015,113(10):1558-1570.
The gut microbiota-targeted whole cereal grain intake and its mechanism in ameliorating metabolic diseases
GONG Ling-xiao,CAO Wen-yan,WANG Jing*,ZHANG Hui-juan,LIU Ying-li
(Beijing Advanced Innovation Center for Food Nutrition and Human Health,Beijing Higher Institution Engineering Research Center of Food Additives and Ingredients,Beijing Technology & Business University(BTBU),Beijing 100048,China)
A large body of scientific researches has shown the inverse relationship between consumption of whole cereal grains and occurrences of metabolic diseases. Due to the complicated ingredients,which may show synergistic action to mediate the protective effects,the conventional methods are not appropriate to clarify the mechanisms for whole grains on health promotion. Meanwhile,gut microbiota is closely related to human health,which is probably modulated by diet intervention. Thus,research has been increasingly focusing on the potential of ingredients present in whole cereal grains to steer gut microbiota activity and composition in a beneficial way. In this review,special interest will be directed to gut microbiota,whole cereal grains and metabolic disease. The results may bring a new way to elucidate the mechanism of whole cereal grain in ameliorating metabolic diseases. As results,the gut microbiota-targeted whole cereal grain intake will serve for human health better.
whole cereal grain;gut microbiota;metabolic disease;obesity
2016-06-28
龚凌霄(1981-),女,博士,讲师,研究方向:天然产物与功能性食品,E-mail:gonglingxiao@btbu.edu.cn。
*通讯作者:王静(1976-),女,博士,教授,研究方向:功能性食品配料,E-mail:wangjing@th.btbu.edu.cn。
国家自然科学基金项目(31501480,31571940,31271976)。
TS201.4
A
1002-0306(2017)02-0364-06
10.13386/j.issn1002-0306.2017.02.062
