荻和南荻杂交种F1群体主要农艺性状的杂种优势、遗传及相关性分析
2017-02-28艾辛蒋建雄陈智勇覃静萍易自力
艾辛,蒋建雄,陈智勇,覃静萍,易自力*
(1.湖南农业大学园艺园林学院,湖南 长沙410128;2.湖南农业大学生物科学技术学院,湖南 长沙410128)
荻和南荻杂交种F1群体主要农艺性状的杂种优势、遗传及相关性分析
艾辛1,蒋建雄2,陈智勇2,覃静萍2,易自力2*
(1.湖南农业大学园艺园林学院,湖南 长沙410128;2.湖南农业大学生物科学技术学院,湖南 长沙410128)
利用主要农艺性状上具有显著差异的荻和南荻为亲本,杂交得到种间杂交种F1群体(232个单株)为试验材料,对荻和南荻杂交种茎节数、叶片长、叶宽、主茎长、花茎长、花序长、株高、基部茎径、平均单分蘖干重、最大分蘖干重、分蘖数和单株重等12个主要农艺性状的杂种优势进行了度量,采用主基因+多基因混合遗传模型对F1杂交群体的12个主要农艺性状进行遗传和相关性分析。结果表明,在F1分离群体中12个主要农艺性状呈连续的、单峰、偏态分布,说明这些性状为多基因控制的数量性状。除基部茎径外的其他11个农艺性状都有较大的中亲优势,其中单株重、平均单分蘖干重、花序长、主茎长、株高、最大分蘖干重具有显著的超高亲优势,说明杂种优势可以作为荻和南荻育种的主要方法。混合遗传分析表明,花茎长、最大茎重能检测到1对主基因,叶片宽、主茎长、株高、分蘖数、单茎均重、单株重能检测到2对主基因的存在,主基因遗传率大小顺序为叶片宽(87.76%)>单株重(81.48%)>单茎均重(65.12%)>分蘖数(59.20%)>主茎长(49.87%)>株高(48.01%)>花茎长(47.75%)>分蘖最大茎重(37.19%)。产量相关性状中的单株重、单茎均重和分蘖数具有较高的主基因遗传率,适合于早期世代选择。这12个农艺性状间存在一定的相关性,多数性状间的相关性为极显著的正相关,其中单株重与分蘖数、单茎均重、主茎长和株高的相关系数最大,在育种实践中可以利用相关性状进行间接选择。
荻;南荻;主基因+多基因;遗传分析;相关性分析
芒草(Miscanthus)是最具有开发利用潜力的纤维素类能源作物[1],中国是芒草的原产地和遗传多样性中心,其中一个最优良的种类—南荻(M.lutarioriparius)作为中国特有种具有高生物质产量、高纤维素含量及易于栽培的优良特点,在造纸、环境修复和能源利用方面的巨大价值备受关注[2],但南荻分布范围窄,对低温、干旱和盐碱的耐受性较差。荻(M.sacchariflorus)则分布范围广,具有耐寒抗冻、耐干旱盐碱胁迫等优良特性[3-4],但生物产量较低。植物远缘杂种优势的成功利用对荻和南荻的遗传改良有重要的借鉴作用[5-6],南荻与荻的重要性状具有良好的互补性,通过杂交育种的方法将优良性状聚合在一起,不仅能直接利用种间杂种优势,还可以利用杂交种后代进行进一步选择,获得具有更多优良性状的新品种。植物极为重要的经济性状(如作物产量、生育期、品质等)都是数量性状,性状的遗传受为数甚多的基因支配[7],利用主基因+多基因混合遗传模型[8-9]明确判断数量性状的类型并估计其相应的遗传参数,包括主基因对数、主基因的遗传效应、主基因的遗传方差和遗传率、多基因效应、多基因的遗传方差和遗传率等,明确其性状的遗传本质,为植物的遗传改良提供理论依据,且已在多种重要农作物得到了应用[10-24]。芒属植物为高度自交不亲和植物[14],很难得到纯系亲本,亲本的异质化程度很高,在F1代就会接近纯合亲本杂交种F2代的分离水平。本研究利用荻和南荻杂交种F1构建的232个基因型遗传群体,分析性状的分离程度、杂种优势,采用植物数量性状主基因+多基因混合遗传模型,对植株相关主要农艺性状进行混合遗传及相关性分析,探索芒属植物各性状所属的遗传模型,并在此基础上估计其遗传效应,从而为芒属植物的遗传改良和杂种优势利用提供理论指导。
1 材料与方法
1.1 试验材料
试验材料为荻和南荻,荻原产地为甘肃泾川县窑店镇(N 35.19°,E 107.23°),田间种植编号为A0405,南荻原产地为湖南长沙(N 28.12°,E 112.59°),田间编号为A0107,采集时间为2006-2007年,种植于湖南农业大学芒属植物资源圃。2010年1月从湖南农业大学芒属植物资源圃中挖取荻和南荻的地下根状茎根移栽至直径为40 cm,深度为45 cm的塑料钵中,盆栽荻出苗后进行光周期处理,进行开花期的调控(试验结果另文发表),10月中旬选择花期相遇的荻和南荻为材料进行人工授粉杂交,以荻为母本,南荻为父本。杂交时要严格隔离,保证只有父本南荻花序上的花粉落到母本荻的花序上,授粉花序需要挂标签牌进行标记,标签牌上写明父母本编号,授粉日期等。待花序梗干枯,小穗果实硬化呈褐色时采收整个花序,晒干保存。
1.2 荻和南荻杂交种的鉴定及田间性状调查
2011年3月将杂交花序上的颖果脱粒后播种在盛满育苗基质的育苗盘中进行育苗,待苗长到20 cm左右取叶片提取DNA,利用SSR标记进行杂交种真实性鉴定[15],创建荻A0405×南荻A0107杂交F1群体。鉴定真实性后的杂交种按行距2.0 m×2.0 m移栽至湖南浏阳永和镇石佳村芒草试验基地(N 28.30°,E 113.88°),试验地为山地,土质属于贫瘠的底层黄壤土,仅在栽培的第一年施有机肥2000 kg/667 m2。
2013和2014年对每个单株及父母本的全部分蘖进行持续的观察,调查亲本和F1群体中单株的茎节数(Non)、叶片长(Lol)、叶宽(Wol)、主茎长(Lms)、花茎长(Lfs)、花序长(Loi)、株高(Ph)、基部茎径(Sd)、分蘖数(Not)、单茎均重(Wsl)、最大茎重(Wbt)、单株重(Wop)等12个农艺性状,除分蘖数和单株重外,其他植株性状重复3次,并计算2年性状调查的平均值。
农艺学性状测定方法,茎节数:地上部分的节数,不包括花茎;叶片长:目测取5片最大叶片用卷尺测定叶片长度,取平均值即代表最大叶片长度;叶片宽:用米尺测量选取的5片最大叶片最宽部位的宽度,取平均值即代表最大叶片宽度;主茎长:选取5分蘖株,测其自然高度,从地面量至花茎基部节的长度,取平均值,重复3次;花茎长:花茎基部至花序基部长度;株高:选取5株最高的茎秆用直尺测定从地上部分至花序顶端的长度;花序总长:每份材料取5个花序用直尺测量花序基部至花序顶端的长度;基部茎径:用游标卡尺测量单分蘖茎秆基部第1节的外径;分蘖数:是指每株种子苗的全部分蘖植株,分蘖株的高度要达到1.0 m以上;单株重:是指每个基因型种子苗全部分蘖的干重;单茎均重:是指每株种子苗全部分蘖干重除以分蘖数;最大茎重:是指单株全部分蘖中取单茎最高、最粗的5个单分蘖株干重的平均值。叶片的长、宽是在植株生长的旺盛期测定,其他性状的测定是在每年的12月下旬。
1.3 杂种优势分析
杂种优势分别以中亲优势、中亲优势率,超高亲优势、超高亲优势率表示[16]。中亲值(mid-parents value,MPV)为双亲性状的平均值[MPV=(P1+P2)/2],高亲值(high-parents value,HPV)为双亲中性状值最高的值。
中亲优势:Hm=F1-MPV,中亲优势率:RHm=(F1-MPV)/MPV×100%。
超高亲优势:Hh=F1-HPV,超高亲优势率:RHh=(F1-HPV)/HPV×100%。
P1,P2为亲本荻和南荻的性状值,F1为荻和南荻杂交种后代的性状值的平均数。
1.4 荻和南荻花期相关性状的混合遗传分析
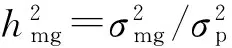
1.5 统计分析方法
采用DPS 14.10统计软件和微软Excel进行统计分析并作直方图。
2 结果与分析
2.1 荻和南荻主要农艺性状在亲本及F1杂交后代的分布
荻A0405和南荻A0107的性状差异比较大,由于2个亲本的异质化程度高,杂种F1群体232个体植株间在很多性状上的差异较大,其描述性统计结果见表1。植物学性状包括叶片,茎秆,花序性状,各性状的表型值的变异情况有较大的差异,各性状的变异系数范围为7.38%~65.24%,其中以产量相关性状的变异系数最大。各性状变异系数由大到小的顺序依次为:单株重>分蘖数>单茎均重>最大茎重>花茎长>叶片宽>主茎长>叶片长>基部茎径>花序长>株高>茎节数,总的来说,植株的株高、茎节数和基部茎径的相关性状变异较小。杂交种群体产量相关性状的变异系数普遍较高,说明产量相关性状在群体中有较大的分离,其中变异系数最大的为单株重,变异系数为65.24%,其次是分蘖数,变异系数为45.34%,单株均重的变异系数为36.23%,最大茎重的变异系数为34.57%,说明了荻和南荻种间杂交后代在产量相关性状上具有较大的选择价值,具有较高的选择效率。
将12个主要农艺性状调查结果进行频次分布的分析,绘制次数分布图(图1)。由图中可知,所有性状的分布是连续的,符合正态分布,有些性状出现不同程度的偏态分布,符合主基因+多基因的遗传特征。
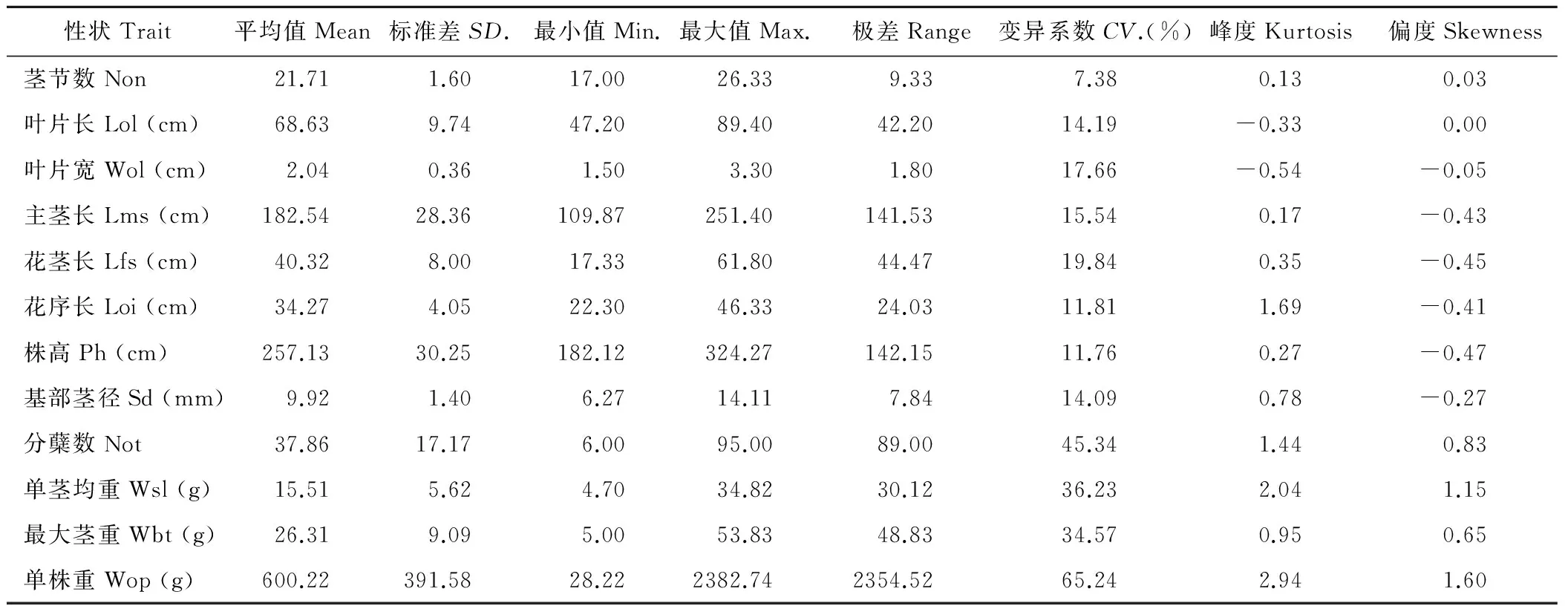
表1 荻与南荻杂交F1群体主要农艺性状表型特征值Table 1 Character values for 12 agronomical traits in F1 population between M. sacchariflorus and M. lutarioriparius
注:Non:茎节数;Lol:叶片长;Wol:叶片宽;Lms:主茎长;Lfs:花茎长;Loi:花序长;Ph:株高;Sd:基部茎径;Not:分蘖数;Wsl:单茎均重;Wbt:最大茎重;Wop:单株重。下同。
Note:Non:Numbers of stem nodes;Lol:Length of leaf;Wol:Width of leaf;Lms:Length of main stem length;Lfs:Length of flower stalk;Loi:Length of inflorescence;Ph:Plant height;Sd:Stem diameter;Not: Number of tiller;Wsl:Average dry weight of single tiller;Wbt: Dry weight of the biggest single stem;Wop: Dry weight of plant. The same below.
2.2 荻和南荻杂交F1群体主要农艺性状的杂种优势
杂交群体荻与南荻的杂交种在茎节数、叶片长、主茎长、花茎长、花序长、分蘖数(茎秆密度)、最大茎重、单茎均重、单株重表现为中亲优势,中亲优势率最高的为单株重(182.90%),其次为单茎均重(59.45%)、花序长(57.82%)、分蘖数(54.53%)、主茎长(39.53%)、株高(37.81%)、最大茎重(29.40%)、花茎长(18.45%)、叶片长(9.55%)、叶片宽(5.97%)、茎节数(4.63%),基部茎径为负优势(-6.82%),偏向于低亲亲本荻。超高亲优势只有单株重、株高、主茎长、花序长、最大茎重具有优势,单株重的超高亲优势最高,达到86.64%,其次是单茎均重35.88%、花序长30.65%、主茎长30.25%、株高23.22%、最大茎重12.76%,其余为负值(茎节数-2.78%、叶片长-4.42%、叶片宽-13.19%、花茎长-4.64%、基部茎径-30.83%、分蘖数-5.35%),基部茎径的最低。多个性状尤其是产量相关性状在多种杂种优势的度量结果表明,基部茎径、分蘖数出现超亲优势是较困难的,但由于提高了平均茎秆重、最大茎重、株高和分蘖数等性状的水平,最终使荻和南荻杂交种产量获得了较大的杂种优势。本试验地为黄壤的成土母质,肥力水平极低,保水能力差,不适宜于亲本南荻的生长,因此亲本南荻的表现极差,在多项性状如在株高、基部茎径等的表现远未达到其品种特征的标准,同时,杂交种的性状表现也会受到恶劣环境的影响,也远未表现应有的杂种优势。但在这样恶劣的生长环境条件下,杂交种表现优良,对贫瘠、干旱的生态条件适应能力良好(表2)。

图1 荻和南荻杂交F1群体主要农艺性状的分布频次Fig.1 Distribution frequency of 12 agronomical traits in F1 population between M. sacchariflorus and M. lutarioriparius表2 荻和南荻杂交F1群体农艺性状的杂种优势Table 2 Heterosis of agronomical traits in F1 population between M. sacchariflorus and M. lutarioriparius

性状Trait母本A0405Femaleparent父本A0107Maleparent中亲值MPV(Mid-parentsvalue)中亲优势Hm(Mid-parentheterosis)中亲优势率RHm(RateMid-parentheterosis)(%)高亲优势Hh(High-parentheterosis)高亲优势率RHh(Rateofhigh-parentheterosis)(%)茎节数Non19.1722.3320.750.964.63-0.62-2.78叶片长Lol(cm)53.5071.8062.655.989.55-3.17-4.42叶片宽Wol(cm)1.502.351.930.125.97-0.31-13.19主茎长Lms(cm)140.15121.50130.8351.7239.5342.3930.25花茎长Lfs(cm)42.2825.8034.046.2818.45-1.96-4.64花序长Loi(cm)26.2317.2021.7212.5657.828.0430.65株高Ph(cm)208.67164.50186.5970.5537.8148.4623.22基部茎径Sd(mm)6.9514.3410.65-0.73-6.82-4.42-30.83分蘖数Not40.009.0024.5013.3654.53-2.14-5.35单茎均重Wsl(g)8.0411.419.735.7859.454.1035.88最大茎重Wbt(g)17.3323.3320.335.9829.402.9812.76单株重Wop(g)321.60102.73212.17388.05182.90278.6286.64
2.3 荻与南荻杂交F1群体主要农艺性状主效+多基因混合模型分析
2.3.1 荻与南荻杂交F1群体最适遗传模型的适合性分析 利用植物数量性状主基因+多基因混合遗传模型单个分离世代的分离分析方法,对荻A0405与南荻A0107杂交F1群体12个主要农艺性状2年表型数据均值进行混合遗传模型分析,获得了无主基因、1对主基因(A)和2对主基因(B)共计11种模型与表型资料的配伍情况,得到AIC值(表3)。根据AIC最小准则,AIC值最小者为最优模型,与最小AIC值比较接近的遗传模型作为备选模型。如单茎均重的AIC值最小为1451.48,与最小AIC比较接近的AIC值为1457.24、1457.40,分别对应的模型为2MG-ADI(B-1)、2MG-AD(B-2)和1MG-AD(A-1)。选择单茎均重的备选最适模型,利用U12、U22、U32,nW2,Dn对备选模型进行适合性检验(表4),选择统计量达到显著个数最少的模型作为最适模型,但2MG-ADI(B-1)达到了显著水平,1MG-AD(A-1)和2MG-AD(B-2)模型的统计量未达到显著水平,以AIC值最小为原则,则选择2MG-AD(B-2)模型为最适模型。
选择12个性状的备选最适模型,利用U12、U22、U32,nW2,Dn对备选模型进行适合性检验(表4),选择统计量达到显著个数最少的模型作为最适模型,多数性状能检测出主效基因的存在。茎节数性状的最适模型分别是0MG模型,没有检测到主效基因的存在。叶片宽性状的最适模型都是2MG-A,都有两对基因控制,主基因表现为加性-显性-上位性。株高相关性状主茎长、花茎长、花序长和株高4个性状的最适模型分别是2MG-EEAD,1MG-AED,0MG,2MG-EEAD模型,花序长检测不到主效基因的存在。基部茎径性状的最适模型是0MG,检测不出主效基因的存在。单茎均重、最大茎重、单株重3个产量相关性状的最适模型分别是2MG-ADI,1MG-AD,2MG-AD,都能检测到主效基因的存在。分蘖数的最适模型为2MG-AD。
2.3.2 荻与南荻杂交F1群体主要农艺性状遗传参数估计 根据性状遗传模型参数的最大似然估计值,估计最适遗传模型的遗传参数(表5)。由表5可知,茎节数、花序长、基部茎径没有找到主基因,其他性状都能找到主基因。控制叶片长的遗传模型为无主基因模型(0MG),控制叶片宽的遗传模型为加性遗传模型(2MG-A),但2对主效基因都比较小(0.26、0.59),叶片宽的主基因方差大于表型方差,遗传率87.76%,为高度遗传力性状。控制茎节数性状未检出主效基因的存在,花序长未能检出主效基因。控制主茎长性状的遗传模型2对主效基因为等显性模型(2MG-EEAD),显性效应相等并于加性效应相等,没有上位性效应,加性效应为16.21,遗传率为49.87%,属中等遗传力性状。花茎长可检测出2对主基因,符合完全显性遗传模型(2MG-AED),显性效应为6.82,

表3 荻与南荻杂交F1群体不同遗传模型下的AIC值Table 3 Akaike’s information criterion value (AIC) under different genetic model for different traits of F1 population between M. sacchariflorus and M. lutarioriparius
下划线表示的AIC值最小。The underlined value refers to the minimum AIC value.表中A-0、A-1、A-2、A-3、A-4、B-1、B-2、B-3、B-4、B-5、B-6分别对应0MG、1MG-AD、1MG-A、1MG-EAD、1MG-AEND、2MG-ADI、2MG-AD、2MG-A、2MG-EA、2MG-AED、2MG-EEAD遗传模型。A-0,A-1,A-2,A-3,A-4,B-1,B-2,B-3,B-4,B-5,B-6 respectively corresponding to 0MG,1MG-AD,1MG-A,1MG-EAD,1MG-AEND,2MG-ADI,2MG-AD,2MG-A,2MG-EA,2MG-AED,2MG-EEAD genetic model.
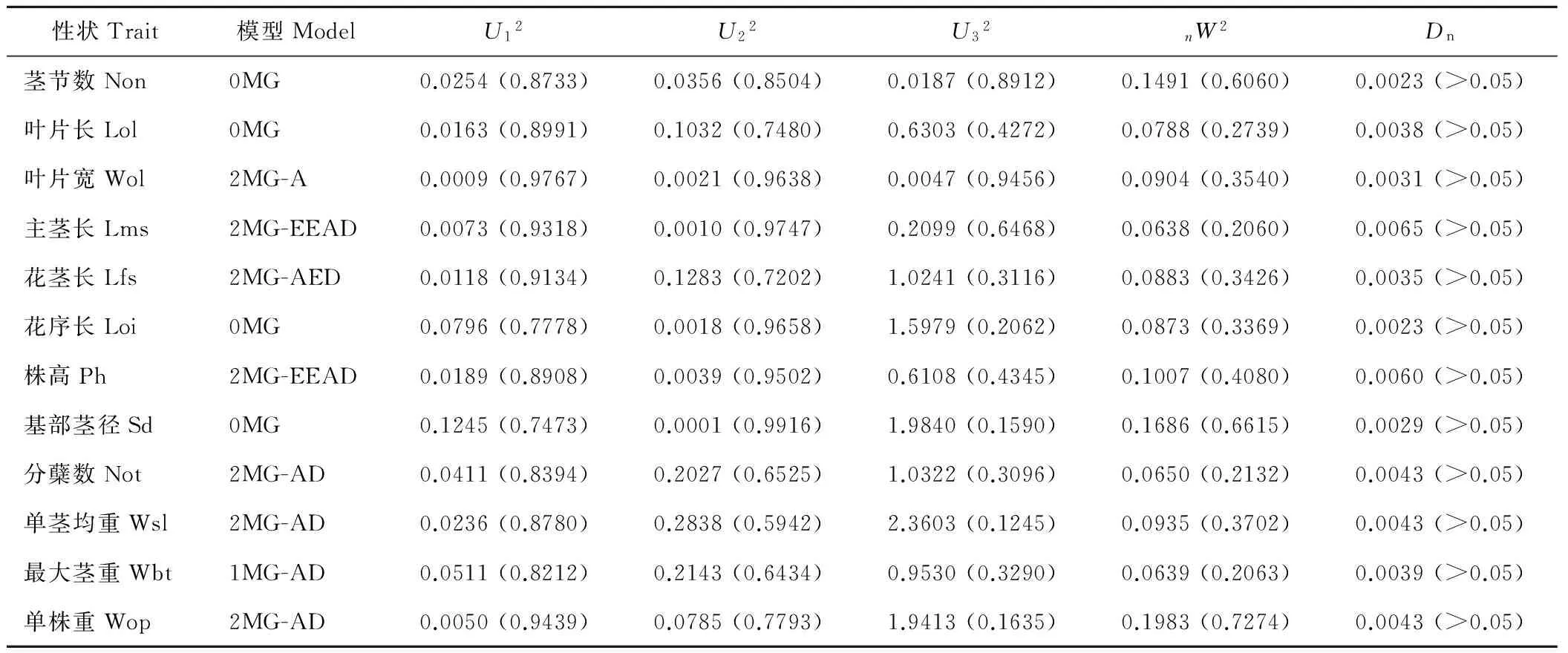
表4 荻与南荻杂交F1群体农艺性状入选模型的适合性检验Table 4 Fitness test of the selected genetic model of 12 agronomical traits of F1 population betweenM. sacchariflorus and M. lutarioriparius
U12、U22、U32、nW2和Dn是适合性检验的5个统计量,其中U12、U22、U32为均匀性检验,nW2为Smirnov检验,Dn为Kolmogorov检验。括号内数值为概率值。U12,U22,U32,nW2,Dnare five statistic values for fitness test;U12,U22,U32are the values of homogeneity testing;nW2is the value of Smirnov test;Dnis the value of Kolmogorov test. The values in the hraeket mean probability.
遗传率为47.75%,属于中度遗传力性状。株高2对主效基因为等显性模型(2MG-EEAD),显性效应相等并于加性效应相等,没有上位性效应,加性效应为16.95,遗传率为48.01%,属中等遗传力性状。基部茎径性状是芒属植物最重要的农艺性状之一,在本试验中却未能检测出主基因的存在,需要进一步的研究。分蘖数有2对主效基因存在,符合加性-显性模型(2MG-AD),第1对主效基因和第2对主效基因的加性效应分别为10.30、12.53,第2对主效基因的加性效应略大于第1对主效基因,2对主效基因的显性效应为负效应,分别为-14.49、-9.10,第1对主效基因的显性效应大于第2对的显性效应,2对主效基因间不存在互作关系。最大茎重为1对主效基因,主效基因为加性-显性效应(1MG-AD),加性效应(6.62)和显性效应(-6.62)数值几乎相等,但加性效应为正效应,显性效应为负效应。单茎均重的2对主效基因的遗传为加性-显性模型(2MG-AD),2对主效基因的加性效应分别为4.57、3.72,2对主效基因的显性效应都为负效应,分别为-6.43、-2.19,以第1对主效基因为主。单株重检测出2对主效基因,符合加性-显性模型(2MG-AD),第1对主效基因的加性效应分别为434.02,第2对主效基因的加性效应分别为230.26,第1对主效基因的加性效应比第2对主效基因的加性效应大约1倍,2个性状的显性效应都为负效应,第1对主效基因的显性效应分别为-610.84,第2对主效基因的显性效应分别为-85.59,明显的以第1对主效基因的显性效应为主,单株重的2对主效基因间没有互作效应。分蘖数、单茎均重、最大茎重、单株重的遗传率分别为59.20%、65.12%、37.19%、81.48%,说明单茎均重、单株重和分蘖数为较高遗传力性状,最大茎重低遗传力性状。

表5 农艺性状在入选模型下的遗传参数估计Table 5 Genetic parameter estimate of agronomical traits under the selected model
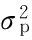
2.4 主要农艺性状的相关性分析
应用DPS Statistics 14.10软件相关性统计分析方法,利用Pearson方法对植株与产量相关的12个主要农艺性状开展相关性分析(表6),结果表明,多数性状间存在显著或极显著相关性,除花序和花茎长性状出现负相关外,其他性状都为正相关,说明对这些具有相关性的性状选择都有正效应。茎节数与叶片宽、主茎长、花茎长、株高、基部茎径、分蘖数、最大茎重、单株重存在极显著相关,与单茎均重存在显著性相关,除与花序长、花茎长为负相关外,其他都为正相关。主茎长和株高与多数性状都为显著或极显著正相关,说明茎高相关性状在性状选择上的重要性。与单株重相关的性状有茎节数、主茎长、株高、基部茎径、分蘖数、单茎均重、最大茎重,都达到了极显著正相关,相关系数分别为0.209**、0.563**、0.507**、0.347**、0.775**、0.628**、0.369**,其中相关性最大的是分蘖数和单茎均重,其次是主茎长和株高。

表6 荻与南荻杂交F1群体12个主要农艺性状的表型相关分析Table 6 Correlation analysis among 12 agronomic traits in F1 population between M. sacchariflorus and M. lutarioriparius
3 讨论
3.1 荻和南荻种间杂交种的杂种优势表现
远缘杂交可以实现不同物种间优良性状的转移,在培育抗病和抗逆品种,提高生态适应能力和改善品质等方面得到了广泛的应用[18-20],尤其很多物种的远缘杂种在生物产量上具有较大的杂种优势[21]。杂种优势是植物界普遍存在的现象,是植物杂交育种的主要途径之一。植物杂种优势的大小主要来源于亲本的遗传异质性,有差异的基因位点数越多、差异越大,其配组的杂交后代出现杂种优势的可能性越大[13-22],选择遗传差异大的亲本进行杂交组合的配制,是杂种优势利用亲本选择选配的最基本的重要原则之一。本研究中选择的荻和南荻不仅地理远缘,而且还是种属远缘的2个亲本,它们的遗传差异,包括生物质产量、生态和生境适应性、品质等方面都有较大的差异,尤其是在生物质产量上的差异极大。本试验中的南荻株高可以达到4 m以上,基部茎径可达2.0 cm以上,而荻一般在原产地株高在3 m左右,基部茎径在1.2 cm左右,由于栽培试验地为黄壤的成土母质,非常贫瘠,加之没有进行肥水管理,其亲本及其杂交群体特性没有得到充分表达。从本试验结果来看,在这种极其恶劣的生长条件下,荻和南荻杂交种仍然具有极强的杂种优势。在芒属植物的4个主要种类中,荻和南荻的性状相似性较高,缺乏互补性状,其杂种优势的表现远小于荻和五节芒(M.floridulus)、南荻和五节芒和南荻和芒(M.sinensis)的杂种优势(另文发表),因为它们具有较多的互补性状,表现出了极高的杂种优势,如分蘖数、单株生物质产量也表现出较大的优势。杂种优势较大的几个重要农艺性状其变异系数也较大,说明这些性状在群体的变异也较大,选育出杂种优势明显的优良单株的可能性也很大,选择效率高,这为荻和南荻的遗传改良提供了多种可能性。由于荻和南荻杂交种的茎节具有发育良好的腋芽,可以进行高效的无性繁殖而培育无性系杂种一代新品种[23]。
3.2 荻和南荻杂交种主要农艺性状的遗传效应
本研究利用主基因+多基因混合遗传模型和相关软件对荻和南荻种间杂交种F1群体主要农艺性状的遗传行为进行了研究,有利于揭示各性状的遗传行为并确定合理选择方法,该方法在多种作物和多种性状的选择上得到了较好的研究结果[10-11,24]。该研究的分析结果表明杂交种的12个主要农艺性状中叶片长宽、主茎长、花茎长、株高、分蘖数、单茎均重、最大茎重、单株重8个性状能检测到1对或2对主基因的存在,其余性状检测不到主基因的存在。8个性状主基因遗传率大小的顺序依次为叶片宽>单株重>单茎均重>分蘖数>主茎长>株高>花茎长>最大单茎重,其中叶片宽、单株重、单茎均重、分蘖数4个性状的主基因的遗传率大于50%,属于高遗传力性状,受环境影响较小,可以根据表现型在早期世代进行选择[25]。本研究中出现的主基因遗传率高的性状,不仅变异系数大,且杂种优势也强,说明利用杂交技术创新种质及选育新品种具有特别重要的意义。对于基部茎径等无主基因且变异系数较小、没有杂种优势的性状,或在不同群体中主基因有无不确定的性状,建议采用轮回选择或回交育种的方法强化这些性状的表现[26]。由于荻和南荻的遗传研究刚刚起步,缺少更多可参考的研究数据,本试验结果能否合理解释荻和南荻主要农艺性状遗传实质,还需要通过建立更多的杂交群体进行深入研究。
3.3 荻和南荻杂交种主要农艺性状的相关性
芒属植物作为能源植物进行品种选育才刚刚起步,对性状间的复杂关系缺乏系统研究,不能为育种提供有效参考。农艺性状是构成作物品种的主要特性,群体内株间的同一性状存在多样性,且各性状对相同生境因子影响的反应不同,存在一定的相关性,育种实践中可以利用这种性状间的相关性选择合适的标准对性状进行间接选择,提高选择效率。在主要农艺性状的相关性方面,与单株重相关性较大的性状有分蘖数、单茎均重、主茎长、株高、最大茎重、基部茎径、茎节数,都达到了极显著正相关,其中相关性最大的是分蘖数和单茎均重,其次是主茎长和株高,这与芒[27]、水稻(Oryzasativa)[28]、小麦(Triticumaestivum)[29]、扁穗雀麦(Bromuscatharticus)[30]的研究结果有差异。出现这种结果的原因与荻和南荻及其杂交种的分蘖散生,分蘖密度较低有关,分蘖间没有形成竞争性关系,如果分蘖密度增加,基部茎径和单茎均重等性状就会因为植株的生长空间竞争而与分蘖密度呈负相关。叶片性状与杂交种产量没有相关性,这是由于荻和南荻杂交种产量构成性状的特点,植株在生长过程中叶片逐渐脱落,荻和南荻杂交种的产量主要来源于茎秆,这与蒋林峰等[31]对鸭茅(Dactylisglomerata)的性状分析结果有一定差异。12个农艺性状的相关性分析表明,荻和南荻杂交后代的大多数农艺性状间呈显著或极显著的正相关关系,尤其是单株重与分蘖数、单茎均重、主茎长和株高有极显著的正相关关系,因此对这些性状的正向选择有利于提高选择效果,但这一结论只是利用本试验所用的F1群体得出的,在今后的研究中还有待于利用更多的杂交群体对结果进行进一步的检验。
References:
[1] Yan J, Chen W L, Luo F,etal. Variability and adaptability ofMiscanthusspecies evaluated for energy crop domestication. Global Change Biology Bioenergy, 2012, 4(1): 49-60.
[2] Liu L, Zhu M, Zhu T P. Exploitation and utilization ofMicanthusandTriiarrhena. Journal of Natural Resources, 2001, 6(16): 562-563. 刘亮, 朱明, 朱太平. 芒荻类植物资源的开发和利用. 自然资源学报, 2001, 16(6): 562-563.
[3] Zong J Q, Chen J B, Nie D Y,etal. Preliminary evaluation on salinity tolerance ofMiscanthussinensisandM.sacchariflorus(Maxim.) Benth. of China. Acta Agrestia Sinica, 2011, 19(5): 803-807. 宗俊勤, 陈静波, 聂东阳, 等. 我国不同地区芒和荻种质资源抗盐性的初步评价. 草地学报, 2011, 19(5): 803-807.
[4] Li Q, Li J P, He M,etal. Study on the adaptability ofMisconstruessacchariflorusbiological components to soil water stress. Pratacultural Science, 2013, 30(6): 893-897. 李强, 李加鹏, 何淼. 荻实生苗生物构件对土壤水分胁迫的响应. 草业科学, 2013, 30(6): 893-897.
[5] Zhan Q W, Qian Z Q. Heterosis utilization of hybrid between Sorghum [Sorghumbicolor(L.) Moench] and Sudangrass [Sorghumsudanense(Piper) Stapf]. Acta Agron Sinica, 2004, 30(1): 73-77. 詹秋文, 钱章强. 高粱与苏丹草杂种优势利用的研究. 作物学报, 2004, 30(1): 73-77.
[6] Xu W H, Wang K J, Wang Y J,etal. Comparison of yield and nutritive value inSorghum, sudangrass andSorghum-sudangrass Hybrid. Acta Agron Sinica, 2006, 32(8): 1218-1222. 徐文华, 王空军, 王永军, 等. 高粱、苏丹草及高粱-苏丹草杂交种产量和饲用品质的比较. 作物学报, 2006, 32(8): 1218-1222.
[7] He D H, Zhang X L. Advance in quantitative traits from phenotypic variation to gene discovery. Hereditas, 2006, 28(12): 1613-1618. 贺道华, 张献龙. 数量性状由表型变异到基因发现的研究进展. 遗传, 2006, 28(12): 1613-1618.
[8] Gai J Y, Zhang Y M, Wang J K. Genetic System of Quantitative Traits in Plants[M]. Beijing: Science Press, 2003: 96-166. 盖钧镒, 章元明, 王建康. 植物数量性状遗传体系[M]. 北京: 科学出版社, 2003: 96-166.
[9] Zhang Y M, Gai J Y, Wang J K. Identification of two major genes plus polygenes mixed inheritance model of quantitative traits in B1 and B2, and F2. Journal of Biomathematics, 2000, 15(3): 358-366. 章元明, 盖钧镒, 王建康. 利用B1和B2或F2群体鉴定数量性状主基因+多基因混合遗传模型并估计其遗传效应. 生物数学学报, 2000, 15(3): 358-366.
[10] Wen Y, Lu X P, Mi F G. Main gene+polygene mixed model and its effect on individual yield of Sorghum×Sudan grass. International Journal of Genetics, 2010, 33(4): 199-203. 温莹, 逯晓萍, 米福贵. 高丹草单株产量主基因+多基因混合遗传模型及效应分析. 国际遗传学杂志, 2010, 33(4): 199-203.
[11] Lv L J, Guo Y S, Du L J,etal. Major genes plus polygenes mixed inheritance model for starch contents in barley seed. Journal of Triticeae Crops, 2014, 34(1): 13-22. 吕亮杰, 郭元世, 杜丽杰, 等. 大麦籽粒淀粉含量的主基因+多基因遗传模型分析. 麦类作物学报, 2014, 34(1): 13-22.
[12] Xu L L, Chen F D, Chen S M,etal. Genetic analysis of salt tolerance of F1progenies between chrysanthemum and the intergeneric hybrid of chrysanthemum and crossostephium. Acta Ecologica Sinica, 2013, 33(16): 4902-4908. 许莉莉, 陈发棣, 陈素梅, 等. 栽培菊花与菊属-近缘属属间杂种杂交后代耐盐性的遗传分析. 生态学报, 2013, 33(16): 4902-4908.
[13] Peng H, Chen F D, Fang W M,etal. Heterosis and mixed genetic analysis of branch traits of cut chrysanthemum. Acta Horticulture Sinica, 2013, 40(7): 1327-1336. 彭辉, 陈发棣, 房伟民, 等. 切花小菊分枝性状杂种优势表现与遗传分析. 园艺学报, 2013, 40(7): 1327-1336.
[14] Hu B. Study on Self-incompatibility ofMiscanthus[D]. Changsha: Hunan Agricultural University, 2012. 胡彬. 芒属植物自交不亲和性初步研究[D]. 长沙: 湖南农业大学, 2012.
[15] Wen H, Ai X, Jiang J X,etal. Identification of F1hybrid progenies betweenMiscanthussacchariflorusandM.lutarioripariusby SSR markers. Grassland and Turf, 2013, 33(4): 7-11, 22. 文浩, 艾辛, 蒋建雄, 等. 采用SSR分子标记鉴定荻与南荻F1杂交种的真实性. 草原与草坪, 2013, 33(4): 7-11, 22.
[16] Li B, Wu R. Heterosis and genotype×environment interactions of juvenile aspens in two contrasting sites. Canadian Journal of Forestry Research, 1997, 7(10): 1525-1537.
[17] Cao X W, Liu B, Zhang Y M. SEA: A software package of segregation analysis of quantitative traits in plants. Journal of Nanjing Agricultural University, 2013, 36(6): 1-6. 曹锡文, 刘兵, 章元明. 植物数量性状分离分析Windows软件包SEA的研制. 南京农业大学学报, 2013, 36(6): 1-6.
[18] He Q, Deng H F, Shu F,etal. Present status on utilization of distant hybridization and heterosisin rice. Journal of Agricultural Science and Technology, 2007, 9(5): 24-29. 何强, 邓华凤, 舒服, 等. 水稻远缘杂交及杂种优势利用现状. 中国农业科技导报, 2007, 9(5): 24-29.
[19] Xie F T, Chou X Q, Zhang H J,etal. F1heterosis of soybeans crossed from varieties released in different geographic places and decades. Soybean Science, 2007, 26(6): 857-861. 谢甫绨, 丑晓奇, 张惠君, 等. 大豆品种远缘杂交F1代的杂种优势分析. 大豆科学, 2007, 26(6): 857-861.
[20] Li H M, Zhao F W, Li A G,etal. Study on seed set and heterosis of distant cross of Upland Rice (OryzasativaL.)×Long-Awn-Barnyard-Grass (Echinochloacaudata). Acta Agriculture Nucleatae Sinica, 2003, 17(1): 11-15. 李慧敏, 赵凤梧, 李爱国, 等. 旱稻(OryzasativaL.)×长芒稗(Echinochloacaudata)远缘杂交后代结实率及杂种优势分析. 核农学报, 2003, 17(1): 11-15.
[21] Zhang B Q, Zhou S, Yang C F,etal. Biomass and root characters of intergeneric hybridErianthusarudinaceus×Saccharumspontaneumand its progeny. Journal of China Agricultural University, 2016, 21(4): 18-25. 张保青, 周珊, 杨翠芳, 等. 斑割复合体及其杂交后代的生物量及根系性状. 中国农业大学学报, 2016, 21(4): 18-25.
[22] Chen S L, Li Y R, Cheng Z S,etal. Heterosis and genetic analysis of oil content in peanut using mixed model of major gene and polygene. Scientia Agricultura Sinica, 2009, 42(9): 3048-3057. 陈四龙, 李玉荣, 程增书, 等. 花生含油量杂种优势表现及主基因+多基因遗传效应分析. 中国农业科学, 2009, 42(9): 3048-3057.
[23] Wei J, Xiao L, Yang S,etal. Effects of rooting agent on the cutting and seedling growth ofMiscanthuslutarioriparia. Acta Agrestia Sinica, 2015, 23(5): 1002-1006. 魏娟, 肖亮, 杨塞, 等. 不同浓度生根粉浸泡南荻插茎对其存活及幼苗生长的影响. 草地学报, 2015, 23(5): 1002-1006.
[24] Huang Y X, Guo T, Li W,etal. Analysis of percentage of productive tiller in rice by the genetic model “major gene plus polygene”. Journal of South China Agricultural University, 2013, 34(3): 300-303. 黄永相, 郭涛, 李伟, 等. 用“主基因+多基因”遗传模型分析水稻茎蘖成穗率. 华南农业大学学报, 2013, 34(3): 300-303.
[25] Zhan Q W, Lin P, Li D,etal. Genetic analysis of main agronomic characters ofSorghumbicolor×Sorghumsudanense. Journal of Nanjing Agricultural University, 2010, 33(3): 26-30. 詹秋文, 林平, 李娣, 等. 高丹草重要农艺性状的遗传分析. 南京农业大学学报, 2010, 33(3): 26-30.
[26] Guo H L, Gao Y D, Xue D D,etal. Genetic analysis of cold tolerance of Zoysia grass. Acta Prataculturae Sinica, 2009, 18(3): 53-58. 郭海林, 高雅丹, 薛丹丹, 等. 结缕草属植物抗寒性的遗传分析. 草业学报, 2009, 18(3): 53-58.
[27] Nie G, Zhang X Q, Huang L K,etal. Phenotypic variation of wildMiscanthussinensispopulations from southwestern China. Acta Prataculturae Sinica, 2013, 22(5): 52-61. 聂刚, 张新全, 黄琳凯, 等. 中国西南区野生芒居群表型变异研究. 草业学报, 2013, 22(5): 52-61.
[28] Liu G F, Xu H M, Yang J,etal. Genetic analysis on tiler number and plant height per plant in rice (OryzasativaL.). Journal of Zhejiang University: Agric. & Life Sci., 2006, 32(5): 527-534. 刘桂富, 徐海明, 杨剑, 等. 水稻(OryzasativaL.)分蘖数和株高的遗传分析(英文). 浙江大学学报: 农业与生命科学版, 2006, 32(5): 527-534.
[29] Zhang J, Zhang D Y, Wang J A,etal. The dependence study of the effective tillers per plant and agronomic characters in Wheat. Journal of Shanxi Agriculture Science, 2009, 37(6): 17-19, 26. 张晶, 张定一, 王姣爱, 等. 小麦单株有效分蘖数与农艺性状的相关性研究. 山西农业科学, 2009, 37(6): 17-19, 26.
[30] Ma X, Zhou C J, Zhang C L,etal. Patterns of morphological variation and agronomic traits in a worldwide sample of prairie grass germplasm. Acta Agrestia Sinica, 2015, 23(5): 1048-1056. 马啸, 周朝杰, 张成林, 等. 扁穗雀麦种质资源形态和农艺性状变异的初步分析. 草地学报, 2015, 23(5): 1048-1056.
[31] Jiang L F, Zhang X Q, Fu Y F,etal. Agronomic trait variation of some main orchard grass (Dactylisglomerata) cultivars in China. Acta Prataculturae Sinica, 2015, 24(3): 142-154. 蒋林峰, 张新全, 付玉凤, 等. 中国主要鸭茅品种农艺性状变异研究. 草业学报, 2015, 24(3): 142-154.
Heterosis, genetic and correlation analysis of main agronomic traits in F1population derived from crossing betweenMiscanthussacchariflorusandM.lutarioriparius
AI Xin1, JIANG Jian-Xiong2, CHEN Zhi-Yong2, QIN Jing-Ping2, YI Zi-Li2*
1.CollegeofHorticulture&Landscape,HunanAgriculturalUniversity,Changsha410128,China; 2.CollegeofBioscience&Biotechnology,HunanAgriculturalUniversity,Changsha410128,China
A F1population with 232 individuals was developed from a cross betweenMiscanthussacchariflorusandM.lutarioriparius, twoMiscanthusspecies significantly different in the main agronomic traits. A total of 12 traits including the number of nod (Non) , the length of leaf (Lol), the width of leaf (Wol), the length of main stem (Lms), the length of flower stalk (Lfs), the length of inflorescence (Loi), the plant height (Ph), the basal stem diameter (Sd), the number of tillers (Not), the average dry weight of single tiller (Wsl), the average dry weight of the biggest single tiller (Wbt), the weight of individual plant (Wop) were determined, and then subjected to genetic and correlation analysis using a mixed major gene plus polygene inheritance model. The results showed that each of trait produced a continuous, single peak and skewed distribution pattern, suggesting that they are quantitative traits. Except for the stem diameter (Sd), the other 11 traits exhibited a mid-parent heterosis, especially in which the Wop, Wsl, Loi, Not, Lms, Ph and Wbt even exhibited a heterobeltiosis, which indicates that the hybrid heterosis would be a major breeding method forMiscanthus. In addition, the mixed factors genetic analysis indicated that the performance of Lfs and Wbt might be controlled by a pair of major genes, and those of Wol, Lms, Ph, Not, Wsl and Wop by 2 pairs of major genes. The orders of the heritability of major genes for each trait are as follows: Wol (87.76%)>Wsl (81.48%)>Wop (65.12%)>Not (59.20%)>Lms (49.87%)>Ph (48.01%)>Lfs (47.75%) >Wbt (37.19%). Some yield-related traits comprising ‘Wop’, ‘Wsl’ and ‘Not’ are suitable to be selected in early generation because they are with a higher major gene effect. Correlation analysis among traits revealed that Wop is positively correlated with Non, Lms, Ph, Sd, Not, Wsl, and Wbt. Of those, Not, Wsl, Lms and Ph gave the highest correlation with Wop, and thus these traits could be selected indirectly according to their related trait performance in breeding practice.
Miscanthussacchariflorus;M.lutarioriparius; major gene plus polygene; genetic analysis; correlation analysis
10.11686/cyxb2016370
http://cyxb.lzu.edu.cn
2016-10-08;改回日期:2016-12-09
国家自然科学基金项目(芒属植物光周期敏感和衰老性状的遗传特性与分子标记研究,31271788)资助。
艾辛(1963-),男,湖南桃源人,副教授,博士。E-mail:hnaixin@126.com
*通信作者Corresponding author. E-mail:yizili998@163.com
艾辛, 蒋建雄, 陈智勇, 覃静萍, 易自力. 荻和南荻杂交种F1群体主要农艺性状的杂种优势、遗传及相关性分析. 草业学报, 2017, 26(2): 111-122.
AI Xin, JIANG Jian-Xiong, CHEN Zhi-Yong, QIN Jing-Ping, YI Zi-Li. Heterosis, genetic and correlation analysis of main agronomic traits in F1population derived from crossing betweenMiscanthussacchariflorusandM.lutarioriparius. Acta Prataculturae Sinica, 2017, 26(2): 111-122.
