南四湖浮游植物多样性与群落生物量、时间序列稳定性关系
2017-02-22张化永王中玉苗明升华北电力大学工程生态学与非线性科学研究中心北京006山东大学环境科学与工程学院山东济南5000山东师范大学生命科学学院山东济南5004
田 旺,张化永*,王中玉,张 建,苗明升,赵 磊(.华北电力大学,工程生态学与非线性科学研究中心,北京 006;.山东大学环境科学与工程学院,山东 济南 5000;.山东师范大学生命科学学院,山东 济南5004)
南四湖浮游植物多样性与群落生物量、时间序列稳定性关系
田 旺1,张化永1*,王中玉1,张 建2,苗明升3,赵 磊1(1.华北电力大学,工程生态学与非线性科学研究中心,北京 102206;2.山东大学环境科学与工程学院,山东 济南 250100;3.山东师范大学生命科学学院,山东 济南250014)
基于2011到2014年对南四湖浮游植物进行的连续监测,研究浮游植物多样性(物种组成和均匀度)与生物量和群落稳定性的关系.结果表明,调查期间共鉴别出浮游植物8门78属138种.湖泊浮游植物细胞密度在5.09×105个/L到6.95×106个/L之间变化,生物量在0.44mg/L到5.46mg/L之间变化.在温度较低的春季和冬季,浮游植物生物量随着物种数的增加而增加,多样性高的群落有更高的生产力.在夏季,浮游植物生物量随着物种数的增加而降低,多样性高的群落具有相对较低的生物量.蓝藻、硅藻和总浮游植物稳定性指数与浮游植物物种数均呈单峰关系.浮游植物均匀度与各个群落稳定性指数的相关系数均不显著.这些结果表明,浮游植物多样性与群落生物量和稳定性有比较复杂的关系:物种数与生物量呈线性关系且受季节变化的影响,与稳定性呈单峰曲线关系;均匀度指数与生物量和稳定性都没有显著的关系.
浮游植物;物种组成;均匀度;群落生物量;时间序列稳定性
生物多样性与群落生产力和稳定性的关系是生态学研究的核心问题之一[1-2].近年来由于人类活动引发的湖泊富营养化造成了大量的物种灭绝和水华爆发[3-7].浮游植物是水生态系统最主要的初级生产者,并且敏感地受到环境变化的影响[8-9].研究浮游植物多样性与生物量和稳定性的关系对保护水生态系统具有重要的意义[2].
一般而言,多样性高的群落具有更高的初级生产力和稳定性[10].物种多的群落由于取样效应和生态位互补效应,能包含更多的高产物种并且能更加充分的利用资源而获得较高的生物量
[10-11].群落中不同物种在性质上的互补使得它们密度变化的异步性增强,从而使群落具有更高的稳定性[12].浮游植物具有较为特殊的生理生态特性,生态学家在研究其多样性对生物量和稳定性的影响时得到很多不同的结论:有正的影响
[13]、负的影响[14]、没有影响或者更加复杂的影响
[15-16].很多生态学家利用实验室培养的浮游植物群落证明多样性高的群落具有较高的生物量
[2,13].在复杂的自然界,Zimmerman等[17]利用北美湖泊数据验证了浮游植物多样性对群落生物量有正的影响.然而Vallina等[18]发现海洋浮游植物的多样性与群落生物量成单峰曲线关系.Steudel等[19]研究发现浮游植物多样性和群落生物量的关系随着环境因子而改变.McGrady-Steed等[13]基于室内实验发现多样性高的浮游植物群落具有更高的稳定性.然而Gonzalez等[20]利用人工培养的浮游植物群落发现多样性对稳定性没有显著影响.在自然生态系统中,生态学家同样发现了矛盾的结论.Ptacnik等[21]基于欧洲湖泊浮游植物数据得到多样性高的群落具有更高的稳定性.而 Filstrup等[22]在重度富营养化湖泊中发现浮游植物多样性对稳定性有负的作用.此外,浮游植物多样性效应还与湖泊营养状态有关,在贫营养和中营养湖泊中,浮游植物多样性高的群落一般具有较高的生物量和稳定性[13].然而在富营养化湖泊中,浮游植物往往被几种藻类所统治(主要为蓝藻或绿藻群落),其多样性较低却有极高的生物量[22-23].这些浮游植物群落失去对环境变化响应的能力而保持较高的生物量,生态系统的稳定性反而变强[22].由此可见,浮游植物多样性与群落生物量和稳定性的关系比较复杂,并且之前的研究以人为操纵的室内实验为主,基于野外测量数据的分析较少[24].
南四湖是我国北方重要的淡水湖泊,也是南水北调东线工程的重要输水通道和调蓄湖泊
[25-26].目前对南四湖浮游植物的研究主要集中在浮游植物群落组成、密度变化和环境因子的影响[25-28],而对浮游植物的多样性效应研究较少.本文通过在南四湖2011到2014年间的连续调查取样,研究浮游植物群落特征和动态变化,以及浮游植物物种组成和均匀度对群落生物量和稳定性的影响.该研究的目的是探索浮游植物物种多样性与群落生物量和时间序列稳定性的关系.
1 材料与方法
1.1 研究地点
南四湖位于山东省西南部(N116°34′~117°21′, E 34°27′~35°20′),由南阳湖、独山湖、昭阳湖和微山湖四个相互连通的湖泊组成.湖泊水面面积为1266km2,容量6.37×109m3,平均水深1.46m.南四湖属于暖温带季风气候区,年平均气温 13.7℃.湖区年均降水量为 550~720mm,冬、春季节较为干燥,夏季为雨季,占总降雨量的60%以上.

图1 南四湖分布与样点位置Fig.1 Distribution of sample sites in Lake Nansihu
南四湖经历了从污染到恢复的过程,在20世纪 80年代,湖泊处于健康状态,平均总氮(TN)浓度为 0.825mg/L,总磷(TP)浓度为 0.018mg/L[25].湖泊共有浮游植物 8门 116属,浮游动物 249种[27-28].从20世纪80年代末期开始,大量未经处理的工、农业污水排入以及湖泊围网养殖造成的内源释放使得南四湖污染越来越严重.在2000年,南四湖平均 TN浓度为 3.7mg/L,平均 TP浓度0.15mg/L[25].在2002年,湖泊仅有36种浮游植物、28种浮游动物[27-28].南水北调东线工程的建设给南四湖污染治理提供了机会,经过几年严格的环境管理和生态恢复,南四湖的污染状况发生了很大的改变.在 2010年,南四湖平均 TN浓度为1.01mg/L,平均TP浓度为0.09mg/L[26].该湖泊的生态恢复为深入研究水生态浮游生物提供了良好的天然实验室.
1.2 取样及分析测试
根据地表水湖泊监测布设原则[29],在南四湖设置 12个不同的取样地点(图 1).从 2011到2014每年的4月初、7月、9月和11月底进行取样,分别代表每年春、夏、秋、冬四个季节的浮游植物状况.
浮游植物样品在各样点水面下0.5m处收集1L,样品采集后加入固定剂鲁哥试剂,静置24h以上,将样品浓缩至30mL,取0.1mL浓缩液放入浮游植物计数框内进行镜检,对浮游植物进行分类鉴别并计算细胞个数[30-31],其生物量通过体积法转化获得[32].
1.3 浮游植物多样性和稳定性指数计算
浮游植物均匀度采用 Pielou’s 均匀度指数衡量[33].计算公式如下:

式中:Pi是该样点中物种 i的生物量比例;S是该样点的物种数;N是采集样品中物种个体总数;J是Pielou’s 均匀度指数.
本研究采用时间序列稳定性(变异系数)作为浮游植物的稳定性指数[14,34],计算公式如下.
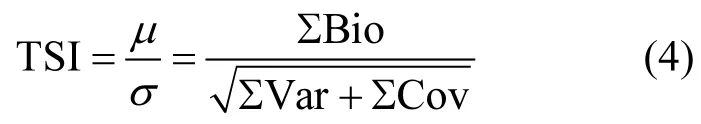
式中:TSI为稳定性指数;µ是各个物种生物量之和;σ为总生物量标准差;Var是各个物种生物量变化的方差;Cov是两两物种生物量变化的协方差;Bio是各个物种的平均生物量.其中物种生物量的方差与协方差通过各个物种在四年、四个季节测得的16个生物量变化数据计算获得.不同浮游植物群落在每个样点计算一个TSI值.在研究浮游植物物种数对 TSI的影响时,由于一个 TSI的计算所需的数据跨越四个年份,因此各个样点物种数为监测期间该样点所出现的全部物种数.不同样点的均匀度用各个物种在调查期间生物量的均值,然后由公式(1)、(2)和(3)计算获得.
1.4 统计分析
浮游植物物种数和均匀度对群落生物量生产力、稳定性的影响通过线性分析、多项式拟合获得,以F检验显著(P<0.05)且r2最大的模型为最优拟合模型.各个群落的稳定性指数差异性通过单因素方差分析获得,统计分析前对数据进行正态性(Kolmogorov-Smirnov检验)和方差齐性检验(Bartlet检验),两两之间的多重比较通过Duncan氏检验法,显著性水平为0.05.
2 结果
2.1 浮游植物多样性特征
调查期间,南四湖共鉴别出浮游植物8门78属 138种,其中绿藻门 35属 60种,占总种数的43.5%.硅藻门16属33种,占总种数的23.9%.蓝藻门9属20种,占总种数的14.5%.裸藻门6属14种,比例为 10.1%.其余门类物种数较少,黄藻门、甲藻门、隐藻门、金藻门共11种.
从季节变化来看,浮游植物物种组成在相邻的两个季节都有明显的变化(图2).在每个调查的年份中,浮游植物物种数均在夏季达到最高值,冬、春季节最低(图2).各个季节间绿藻的物种组成变化最大(图2).各个不同的取样地点物种数从20到88种之间变化.
2.2 浮游植物细胞密度与生物量
研究期间湖泊浮游植物密度均值的变化范围是5.09×105个/L (2013年冬)到6.95×106个/L (2014年夏).浮游植物密度总体均值为2.79×106个/L,其中绿藻门 1.50×106个/L,占总密度的53.7%.硅藻门5.64×105个/L,占总密度的20.2%.蓝藻门 4.91×105个/L,比例为 17.6%.裸藻门1.91×105个/L,占总密度的6.86%.在温度较低的冬季,硅藻门密度占总密度的 40.3%,绿藻门和蓝藻门的比例均有所下降,分别为 38.4%和13.9%.在秋季,绿藻门密度占总密度的70.4%,达到最大值,而硅藻平均密度的比例达到最小值,为8.17%.

图2 2011到2014年南四湖浮游植物物种组成、生物量、细胞密度季节变化Fig.2 Seasonal variations in phytoplankton species richness, biomass and abundance from 2011 to 2014
湖泊浮游植物生物量均值的变化范围是0.44mg/L(2013年冬)到5.46mg/L(2011年夏).浮游植物总生物量均值为 2.19mg/L,其中绿藻门0.94mg/L,占总生物量的 43.0%.硅藻门 0.45mg/L,占总生物量的 20.7%.裸藻门 0.43mg/L,比例为19.4%.蓝藻门由于个体较小,其均值为0.20mg/L,比例为8.92%.在温度较低的冬季,硅藻门生物量占总生物量的比例达最大值,为41.9%,绿藻门和蓝藻门的比例均有所下降,分别为32.6%和7.11%.在秋季,绿藻门生物量占总生物量的59.2%,达到最大值,而硅藻平均生物量的比例达到最小值,为8.82%.
2.3 浮游植物物种多样性与生物量的关系
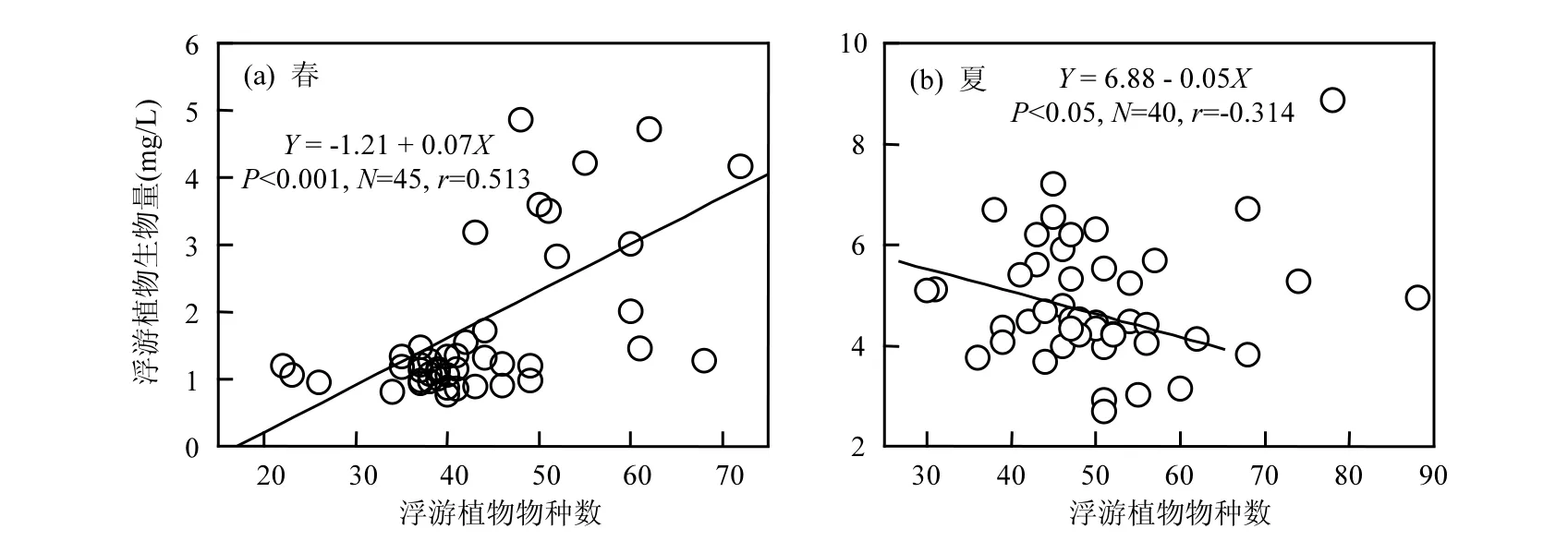
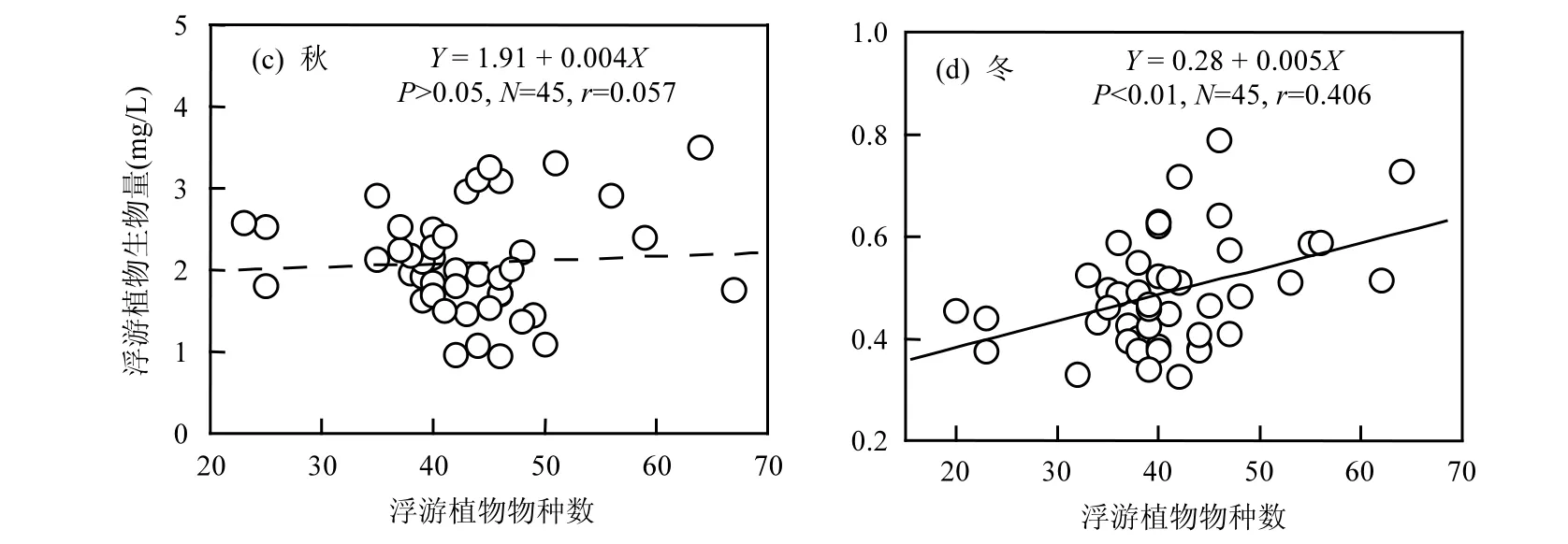
图3 浮游植物物种数对生物量的影响Fig.3 Effect of species richness on phytoplankton biomass
图3和表1可以明显的观测到,在温度较低的冬季和春季,浮游植物生物量随着物种数的增加而增加(P<0.001,r=0.513).浮游植物每增加一个物种,其生物量平均增加0.07mg/L.在温度最低的冬季,浮游植物生物量和物种数的回归斜率为0.005(表 1).在温度最高的夏季,浮游植物生物量和物种数的相关系数不显著(r=0.106).但是进一步的分析显示,在浮游植物物种数低于65的样点,浮游植物生物量随着物种数的增加而降低(P<0.05,r=-0.314).浮游植物每增加一个物种,其生物量平均降低0.05mg/L.在秋季,浮游植物生物量与物种数没有显著的相关性(r=0.057).

图4 浮游植物均匀度对生物量的影响Fig.4 Effect of species evenness on phytoplankton biomass

表1 浮游植物生物量与物种组成、均匀度的回归分析Table 1 Regression analysis for phytoplankton biomass and species richness or evenness
图4和表1表明,在春季浮游植物生物量随着均匀度的增加而降低(P<0.01,r=-0.453).由图4可以清晰的观测到,在均匀度相对较低的区间(0.65~0.82),浮游植物的生物量值变化较大且分布较为散乱(0.81~7.64mg/L),在均匀度相对较高的群落(0.82~0.92),浮游植物生物量相对较低且集中(0.77~1.72mg/L).在夏、秋、冬三个季节,浮游植物生物量与均匀度之间的相关系数都不显著(夏:r =-0.103,秋:r=-0.287,冬:r=-0.171).
2.4 浮游植物物种多样性与群落稳定性的关系
浮游植物物种组成和主要群落稳定性关系如图5所示.从该图可以看出,蓝藻群落的稳定性与浮游植物物种数呈单峰关系(P<0.01,r2=0.740).随着浮游植物物种数的增加,蓝藻群落稳定性指数先增加后降低,在物种数为100时达到最大值(图5).绿藻群落稳定性指数与浮游植物物种数相关系数不显著(r=-0.019).硅藻群落的稳定性与浮游植物物种数的关系与蓝藻类似,也呈单峰关系(P<0.01,r2=0.571).硅藻稳定性指数在浮游植物物种数为93时达到最大值,之后有下降趋势.整个浮游植物群落稳定性指数与物种数关系也呈单峰关系(P<0.05,r2=0.498).

图5 浮游植物物种数对稳定性指数的影响Fig.5 Effect of species richness on phytoplankton stability index

图6 浮游植物均匀度对稳定性指数的影响Fig.6 Effect of evenness on phytoplankton stability index
浮游植物均匀度与主要群落稳定性指数关系如图6所示.蓝藻、绿藻、硅藻和浮游植物的稳定性指数与浮游植物均匀度之间的相关系数分别为:r=-0.401、r=0.206、r=-0.381和r=-0.236,这些相关系数均不显著(P>0.05).从图 6中可以看出,随着浮游植物的稳定性的增加,蓝藻、绿藻、硅藻和浮游植物的稳定性指数没有明显的增加或降低的趋势.

图7 不同浮游植物群落稳定性指数比较,柱状图为均值±标准差Fig.7 Compare of stability index of different phytoplankton communities, values are expressed as mean ± SD
图7表明,南四湖不同浮游植物群落的稳定性指数有显著差异(F(4,55)=49.5,P<0.001).单因素方差分析显示,蓝藻与硅藻群落稳定性指数没有显著差别,说明这两个群落对环境变化响应类似.绿藻与浮游植物群落稳定性指数没有显著差别,这与绿藻占浮游植物生物量的比重最大有关.其余群落的两两比较显示稳定性指数均有显著差异,其关系为:浮游植物=绿藻>蓝藻=硅藻>裸藻(稳定性指数的比较).
3 讨论
南四湖经历了从健康到污染再到恢复的过程,其浮游植物从1980s的116属,到2002年减为36种,再到2010年恢复到117种[27-28].在该研究中,2011年到2014年间共鉴别出浮游植物78属138种.与2002年和2010年相比,南四湖浮游植物物种数有较大的增加,但与 1980s相比还有一定的差距[27-28].
浮游植物多样性与群落生物量关系比较复杂,大部分研究认为物种多的群落具有更高的初级生产力[11-12,14,35].也有生态学家发现随着浮游植物多样性的增加,生产力逐渐降低,或者两者呈单峰曲线关系[15-16,22].在该研究中,发现浮游植物物种数与生物量的关系随季节变化:在冬、春季节呈正相关;在夏季呈负相关;在秋季两者无显著关系.在冬季和春季水温较低,浮游植物物种多的群落能产生相对高的生物量来维持湖泊的生产力;在夏季水温较高,适合藻类的生长甚至爆发,物种数高的群落具有相对较低的生物量.这说明,多样性越高,浮游植物生物量的季节波动越小,群落抵抗外界干扰(温度波动)的能力也越强.Steudel等[19]研究发现生物多样性与群落生物量关系受环境变化的影响.王海东等[34]研究发现在高寒草甸,刈割(放牧或捕食)、施肥(营养盐)对物种多样性与生产力的关系有较大的影响.这些研究结果表明,在同一个生态系统中,生物多样性与群落生物量的关系可能会随着时间或者环境的变化而改变.
Filstrup等[22]发现浮游植物均匀度指数与群落生物量呈显著负相关.在南四湖,春季浮游植物生物量随着其均匀度的增加而降低,而在其它季节两者的关系不显著(图 3).进一步的比较显示,Filstrup等[22]的研究在重度富营养化湖泊中,浮游植物群落主要被少数蓝藻物种统治,其均匀度指数较低,主要分布在0~0.7之间,但是群落生物量较高,因此得到的结果是均匀度越低浮游植物生物量越高[22].南四湖属于中富营养化湖泊,浮游植物群落主要被绿藻、硅藻、蓝藻和隐藻共同占据,其均匀度指数相对较高,在 0.64~0.93之间变化,均匀度指数与生物量没有明显的关系(图 4).这些研究结果表明,浮游植物均匀度和群落生物量的关系与湖泊营养状态有关.
生态学家发现生物多样性与群落稳定性的关系同样复杂[36].大部分研究表明物种数多的群落稳定性高[36],也有研究认为群落稳定性随着多样性的增加而降低,或者两者没有明显的关系[37].在本文中,浮游植物物种数与蓝藻、硅藻和总浮游植物的时间序列稳定性都呈单峰关系,而与绿藻群落稳定性的关系不显著(P>0.05).很多实验表明,在局域尺度上,生态系统功能随着物种多样性的增加而呈现出饱和型上升的趋势[38].结合这些研究结果,可以得到,在群落物种多样性较低时,稳定性随着多样性的增加而增加;当物种数达到一定程度时,稳定性不再增加甚至有所回落[38].在南四湖,绿藻门是浮游植物最主要的群落,其细胞密度和生物量分别占总浮游植物的 53.7%和43.0%.然而其稳定性与浮游植物物种数没有显著的相关性,这说明多样性与稳定性关系还与组成群落的物种性质有关.
Isbell等[36]在草地上进行实验发现植物时间序列稳定性与均匀度之间没有明显的关系. Wittebolle等[39]在微宇宙实验中发现在严格的条件下,微生物群落的均匀度与生态系统稳定性呈正相关.在该研究中,发现南四湖浮游植物均匀度与各个群落时间序列稳定性没有显著的关系(图 6),这与 Isbell等[36]研究一致,而与Wittebolle等[39]的结果不同.这说明自然生态系统中物种均匀度与稳定性的关系比较复杂并需要进一步探索.
南四湖是南水北调东线工程的重要的输水通道和调蓄湖泊,其水质安全和生态健康关系到调水是否成功.本文研究结果证实了浮游植物多样性在维持湖泊生态系统生物量和稳定性的重要作用.因此对南四湖,以及我国很多类似湖泊的保护[5,23,40-42],需要在限制其富营养化的同时加强对浮游植物多样性的保护和恢复,发挥它们在生态系统中的作用.
4 结论
4.1 2011到 2014年在南四湖共鉴别出浮游植物8门78属138种,其中绿藻门60种,硅藻门33种,蓝藻门20种,裸藻门14种;各个季节的浮游植物细胞密度均值从5.09×105个/L到6.95×106个/L之间变化,生物量从0.44mg/L到5.46mg/L之间变化,绿藻门是最主要的群落,其次为硅藻门;
4.2 在冬季和春季浮游植物生物量随着物种数的增加而增加,在夏季浮游植物生物量随着物种数的增加而降低,浮游植物多样性高的群落在温度低时有更高的生产力,在温度高时有较低的生物量;
4.3 南四湖总浮游植物和绿藻的稳定性最高,其次为蓝藻和硅藻,裸藻的稳定性最低,浮游植物物种数与蓝藻、硅藻和总浮游植物稳定性指数均呈单峰关系,然而与绿藻群落稳定性没有显著的相关关系;
4.4 在春季浮游植物均匀度与生物量成反比,在其它季节两者关系不显著,浮游植物均匀度对各个群落稳定性指数的影响也都不显著.
该研究对了解南四湖浮游植物多样性对群落生物量和稳定性的影响有重要意义.
[1] 张全国,张大勇.生物多样性与生态系统功能:最新的进展与动向 [J]. 生物多样性, 2003,11(5):351-363.
[2] Giller P S, Hillebrand H, Berninger U G, et al. Biodiversity effects on ecosystem functioning: emerging issues and their experimental test in aquatic environments [J]. Oikos, 2004,104(3):423-436.
[3] 曾庆飞,谷孝鸿,毛志刚,等.固城湖及上下游河道富营养化和浮游藻类现状 [J]. 中国环境科学, 2012,32(8):1487-1494.
[4] 秦伯强,高 光,朱广伟,等.湖泊富营养化及其生态系统响应[J]. 科学通报, 2013,58(10):855-864.
[5] 王 华,杨树平,房晟忠,等.滇池浮游植物群落特征及与环境因子的典范对应分析 [J]. 中国环境科学, 2016,36(2):544-552.
[6] 李俊龙,郑丙辉,张铃松,等.中国主要河口海湾富营养化特征及差异分析 [J]. 中国环境科学, 2016,36(2):506-516.
[7] 李佐琛,段洪涛,张玉超,等.藻源型湖泛发生过程水色变化规律[J]. 中国环境科学, 2015,35(2):524-532.
[8] Weyhenmeyer G A, Peter H, Willen E. Shifts in phytoplankton species richness and biomass along a latitudinal gradientconsequences for relationships between biodiversity and ecosystem functioning [J]. Freshwater Biology, 2013,58(3):612-623.
[9] 吴雅丽,许 海,杨桂军,等.太湖春季藻类生长的磷营养盐阈值研究 [J]. 中国环境科学, 2013,33(9):1622-1629.
[10] Tilman D, Wedin D, Knops J. Productivity and sustainability influenced by biodiversity in grassland ecosystems [J]. Nature, 1996,379(6567):718-720.
[11] Loreau M, Naeem S, Inchausti P, et al. Biodiversity and ecosystem functioning: current knowledge and future challenges [J]. Science, 2001,294(5543):804-808.
[12] Thibaut L M, Connolly S R. Understanding diversity-stability relationships: towards a unified model of portfolio effects [J]. Ecology Letters, 2013,16(2):140-150.
[13] McGrady-Steed J, Harris P M, Morin P J. Biodiversity regulates ecosystem predictability [J]. Nature, 1997,390(6656):162-165.
[14] Schmidtke A, Gaedke U, Weithoff G. A mechanistic basis for underyielding in phytoplankton communities [J]. Ecology, 2010, 91(1):212-221.
[15] Gamfeldt L, Hillebrand H, Jonsson P R. Species richness changes across two trophic levels simultaneously affect prey and consumer biomass [J]. Ecology Letters, 2005,8(7):696-703.
[16] Weis J J, Cardinale B J, Forshay K J, et al. Effects of species diversity on community biomass production change over the course of succession [J]. Ecology, 2007,88(4):929-939.
[17] Zimmerman E K, Cardinale B J. Is the relationship between algal diversity and biomass in North American lakes consistent with biodiversity experiments [J]. Oikos, 2014,123(3):267-278.
[18] Vallina S M, Follows M J, Dutkiewicz S, et al. Global relationship between phytoplankton diversity and productivity in the ocean [J]. Nature Communications, 2014,5(4299):1-10.
[19] Steudel B, Hector A, Friedl T, et al. Biodiversity effects on ecosystem functioning change along environmental stress gradients [J]. Ecology Letters, 2012,15(12):1397-1405.
[20] Gonzalez A, Descamps-Julien B. Population and community variability in randomly fluctuating environments [J]. Oikos 2004, 106(1):105-116.
[21] Ptacnik R, Solimini A G, Andersen T, et al. Diversity predicts stability and resource use efficiency in natural phytoplankton communities [J]. Proceedings of the National Academy of Science, 2008,105(13):5134-5138.
[22] Filstrup C T, Hillebrand H, Heathcote A J, et al. Cyanobacteria dominance influences resource use efficiency and community turnover in phytoplankton and zooplankton communities [J].Ecology Letters, 2014,17(4):464-474.
[23] 钱奎梅,刘 霞,段 明,等.鄱阳湖蓝藻分布及其影响因素分析[J]. 中国环境科学, 2016,36(1):261-267.
[24] Ptacnik R, Moorthi S D, Hillebrand H. Hutchinson reversed, or why there need to be so many species [J]. Advances in Ecological Research, 2010,43:1-43.
[25] 舒凤月,刘玉配,赵 颖,等.南四湖水体氮、磷营养盐时空分布特征及营养状态评价 [J]. 环境科学, 2012,33(11):3748-3752.
[26] 张祖陆,辛良杰,梁春玲.近50年来南四湖湿地水文特征及其生态系统的演化过程分析 [J]. 地理研究, 2007,26(5):957-966.
[27] 巩俊霞,段登选,王志忠,等.南四湖浮游生物调查分析 [J]. 长江大学学报自然科学版:农学卷, 2010,7(1):39-42.
[28] 武周虎,张 可,金玲仁,等.南四湖水质空间分布特征分析与改善效果评估 [J]. 水资源保护, 2012,28(6):1-7.
[29] HJ/T 91-2002 地表水和污水监测技术规范 [S].
[30] 胡鸿钧,魏印心.中国淡水藻类系统、分类及生态 [M]. 北京:科学出版社, 2006.
[31] Arhonditsis G B, Winder M, Brett M T, et al. Patterns and mechanisms of phytoplankton variability in Lake Washington (USA) [J]. Water Research, 2004,38(18):4013-4027.
[32] 孙 军,刘东艳,钱树本.浮游植物生物量研究Ⅰ.浮游植物生物量细胞体积转化法 [J]. 海洋学报, 1999,22(2):75-85.
[33] Pielou E C. The measurement of diversity in different types of biological collections [J]. Journal of Theoretical Biology, 1966, 13(1):131-144.
[34] 王海东,张璐璐,朱志红.刈割、施肥对高寒草甸物种多样性与生态系统功能关系的影响及群落稳定性机制 [J]. 植物生态学报, 2013,37(4):279-295.
[35] 张全国,张大勇.生产力、可靠度与物种多样性:微宇宙实验研究[J]. 生物多样性, 2002,10(2):135-142.
[36] Isbell F I, Polley H W, Wilsey B J. Biodiversity, productivity and the temporal stability of productivity: patterns and processes [J]. Ecology Letters, 2009,12(5):443-451.
[37] Donohue I, Petchey O L, Montoya J M, et al. On the dimensionality of ecological stability [J]. Ecology Letters, 2013, 16(4):421-429.
[38] Bond E M, Chase J M. Biodiversity and ecosystem functioning at local and regional spatial scales [J]. Ecology Letters, 2002,5(4): 467-470.
[39] Wittebolle L, Marzorati M, Clement L, et al. Initial community evenness favours functionality under selective stress [J]. Nature, 2009,458:623-626.
[40] 许 海,秦伯强,朱广伟.太湖不同湖区夏季蓝藻生长的营养盐限制研究 [J]. 中国环境科学, 2012,32(12):2230-2236.
[41] 田 丰,钱 新,陈 众.调水对巢湖浮游植物群落演替模式的影响 [J]. 中国环境科学, 2012,32(12):2224-2229.
[42] 叶琳琳,史小丽,张 民,等.巢湖夏季水华期间水体中溶解性碳水化合物的研究 [J]. 中国环境科学, 2012,32(2):318-323.
Phytoplankton diversity effects on community biomass and temporal stability in Lake Nansihu.
TIAN Wang1, ZHANG Hua-yong1*, WANG Zhong-yu1, ZHANG Jian2, MIAO Ming-sheng3, ZHAO Lei1
(1.Research Center for Engineering Ecology and Nonlinear Science, North China Electric Power University, Beijing 102206, China;2.School of Environmental Science and Engineering, Shandong University, Jinan 250100, China;3.College of Life Science, Shandong Normal University, Jinan 250014, China). China Environmental Science, 2017,37(1):319~327
The phytoplankton community of Lake Nansihu was investigated seasonally from 2011 to 2014. The effects of phytoplankton diversity (species richness and evenness) on both community biomass and temporal stability were analyzed. A total of 138 phytoplankton species belonging to 78 genera and 8 phyla were identified in the lake. Phytoplankton abundance ranged between 5.09×105/L and 6.95×106/L and its biomass varied from 0.44mg/L to 5.46mg/L in different seasons. In spring and winter when the temperature was low, phytoplankton biomass increased with increasing species richness, phytoplankton community with larger number of species was more productive. In summer when the temperature was high, there was a strong negative relationship between phytoplankton biomass and species richness, phytoplankton community with higher diversity had relatively lower biomass. The stability indices of Cyanophyta, Bacillariophyta and total phytoplankton were all unimodally related to phytoplankton species richness. However, the effects of phytoplankton evenness on the Cyanophyta, Chlorophyta, Bacillariophyta or total phytoplankton stability index were not significant (P>0.05). Results in this research indicated that the effects of phytoplankton diversity on community biomass and temporal stability were complex: species richness had a linear relationship with community biomass and influenced by seasonal variations, a unimodal relationship with temporal stability; evenness had no relationship with either community biomass or temporal stability.
phytoplankton;species richness;evenness;community biomass;temporal stability
X171
A
1000-6923(2017)01-0319-09
田 旺(1987-),男,山东滨州人,华北电力大学博士研究生,主要从事淡水生态学研究.发表论文8篇.
2016-05-10
国家水体污染控制与治理科技重大专项(2009ZX07210-009,2015ZX07203-011,2015ZX07204-007)
* 责任作者, 教授, bjecology@gmail.com
