运动技能水平与躯体感觉输入对运动表象的影响*
2017-02-01张兰兰沈诚朱桦李雪佩戴雯吴殷2张
张兰兰沈 诚朱 桦李雪佩戴 雯吴 殷2张 剑
(1上海体育学院运动科学学院) (2上海体育学院经济与管理学院, 上海 200438)
1 前言
运动表象是指在无外部动作输出的情况下对简单或复杂动作进行心理排练或模仿的神经心理过程(Lotze & Cohen, 2006)。作为一种学习和提高技能水平的有效方法, 运动表象被广泛运用于运动员和音乐家的技能训练(Cocks, Moulton, Luu, & Cil,2014; Keller, 2012), 及脑卒中患者的运动康复中(de Vries, Tepper, Otten, & Mulder, 2011)。
运动表象分为视觉表象和动觉表象两种。视觉表象指以外部的、第三视角进行心理模仿形成肢体运动的视觉表征过程(Milton, Small, & Solodkin,2008), 参与情景运动记忆的提取(Greenberg &Knowlton, 2014); 动觉表象主要指以内部的、第一视角对动作进行心理模仿, 侧重于体会动作执行的动觉感受过程(Solodkin, Hlustik, Chen, & Small,2004), 依赖已有的运动经验。已有证据表明, 运动表象与实际执行之间的时间一致性可以反映运动表象的质量, 即运动表象与实际执行用时越接近,运动表象质量越高(Papaxanthis, Pozzo, Skoura, &Schieppati, 2002)。运动表象与运动执行的神经基础具有重叠性(Dechent, Merboldt, & Frahm, 2004), 这种重叠性涉及广泛的脑区, 主要包括与运动控制相关的脑区, 如初级运动皮层(Dechent et al., 2004)、辅助运动区(Kasess et al., 2008)、基底神经节和小脑(Decety et al., 1994)等, 以及镜像神经系统(Abreu et al., 2012; Wu et al., 2013)。由运动前区、顶上小叶和顶内沟(Buccino et al., 2001)、颞上沟(Grèzes,Frith, & Passingham, 2004)以及额下回(Rizzolatti &Craighero, 2004)等组成的镜像神经系统在动作执行、观察和表象(Fogassi et al., 2005), 以及理解动作意图时(Tkach, Reimer, & Hatsopoulos, 2007)活性提高, 被广泛认为是动作模仿的神经基础(Chaminade,Meltzoff, & Decety, 2005)。另一方面, 运动专家在完成专项动作有关的任务时具有与新手不同的行为学表现和神经生理学特征。如 Wu等人(2013)发现运动员具有较强的视觉感知能力, 表现为预判过程中更高的准确率和视觉搜索过程中更为稳定的注视点, 以及相关功能脑区更高的功能活性。Jin等人(2011)发现专业羽毛球运动员优异的预判能力与较大的P300和P2振幅有关。然而, 在运动表象过程中, 运动技能水平的差异是否会导致行为学表现和神经生理学特征的差异还不清楚。本研究的第一个假设是与新手相比, 运动专家具有执行效果更好的运动表象。我们将通过考察运动表象与实际执行的时间一致性(外在行为表现)和镜像神经系统的激活程度(内在神经机制)两个方面来验证此假设。
人类运动与器械使用息息相关, 器械使用给人脑带来可塑性改变(Draganski et al., 2004), 同时大脑将器械纳入使用者的身体图式(Sposito, Bolognini,Vallar, & Maravita, 2012)。持器械状态下, 躯体感觉输入会对运动表象时的大脑活动产生影响(Vargas et al., 2004)。经颅磁刺激(TMS)研究发现, 被动触摸橡皮球带来的躯体感觉输入可以提高表象捏球时的运动诱发电位(Motor evoked potential, MEP)水平(Mizuguchi et al., 2011)。另外, 羽毛球运动员持球拍表象发球比持塑料棒时表现出更大的 MEP,而新手未表现出差异(Wang et al., 2014), 这提示外部的躯体感觉输入对大脑皮层兴奋性的影响与运动技能掌握水平有关, 躯体感觉输入影响运动专家的运动表象。然而, 器械带来的躯体感觉输入影响运动专家运动表象的神经机制仍不清楚。有研究发现运动表象时的大脑活动与任务难度相关, 任务越容易, 镜像神经系统的兴奋性越低(Bakker et al.,2008)。与不持球相比, 篮球运动员在持篮球条件下进行运动表象更具生态效度。因此, 本研究的第二个假设是在器械依赖的运动项目中, 运动专家持器械进行运动表象较不持器械更容易, 表现为镜像神经系统激活程度的下降。我们将通过运动表象问卷的主观评分、镜像神经系统激活程度变化以及二者之间的相关关系来验证此假设。
2 方法
2.1 被试
从上海体育学院招募篮球运动员(平均年龄19.2 ± 1.4岁, 年龄范围18~21岁)和新手各12人(平均年龄18.8 ± 1.6岁, 年龄范围18~22岁)。所有被试均为右利手(Oldfield, 1971), 视力或矫正视力正常。篮球运动员为国家一级运动员, 平均训练时长每周 10.0 ± 1.8 小时, 平均训练年限 10.7 ± 1.4 (年限范围8~13年)。新手为未接受过任何运动项目专业训练的普通大学生。实验前所有被试均签订了知情同意书。
2.2 实验任务
实验任务是要求被试采用动觉表象先想象拍三下球, 然后从 5个不同的角度投篮。5个角度分别为: 标准的投篮点和以篮筐为中点、中线两侧对称45°和90°的4个点。 5个点与篮筐的距离相等(见图 1A)。表象角度的设置不作为实验条件, 主要用于避免单一角度表象带来的学习效应。为考察躯体感觉输入对运动表象的影响, 表象分为持球和不持球两个条件进行。持球条件的设置形式在满足实验室环境的同时兼顾运动项目的生物学特征, 包括:(1)由于被试投篮时主要是右手投篮, 左手仅起辅助作用, 因此持球条件均为右手持球, 左手按键;(2)球放置于被试腹部。被试右手持球的姿势为5个指腹按球, 手心中空。这与篮球运动员在球场控球时的姿势相同。不持球条件下, 左手按键、右手自然放于体侧。两种持球条件下被试均须自主控制表象的进行, 并通过按键标志表象的开始和结束。第一次按键后, 被试开始表象; 当表象进行到篮球出手时, 被试再次按键, 表示表象结束。本研究重点考察动觉表象的神经机制, 所以表象内容不包括篮球空中飞行入框部分。
正式实验前, 对新手进行为期 3周的投篮训练。投篮训练频率为每周3次, 每次训练时间半小时。之后, 使用专业数字摄像机(Sony PXW-F37, 50帧/秒)逐一拍摄所有被试的投篮情况, 并计算从拍球开始到篮球出手之间的时间间隔。将每位被试25次投篮(每个投篮角度各5次)的平均时间定义为实际执行时间。最后, 为使被试熟悉本研究中使用的运动表象程序, 我们对所有被试进行了为期一周的不同持球条件、不同投篮角度下运动表象训练。一共训练3次, 每次半小时。
2.3 实验设计
本研究采用2 × 2的混合设计, 组别(篮球运动员和新手)为被试间变量, 持球条件(持球和不持球)为被试内变量。
2.4 实验程序
实验程序采用 E-prime 2.0软件编制, 为组块设计。整个实验分为持球和不持球两个部分, 每个部分包含5个组块, 每个组块有5个试次。为抵消持球条件先后顺序可能带来的影响, 对两个部分的顺序进行了抵消平衡。每个试次的开始先呈现一个“+”, 呈现时间为1 s。然后呈现一张描述投篮角度的文字, 如“右侧 45°”, 被试通过头圈上的反射镜面观看屏幕显示的内容。被试清楚任务内容后按键开始表象, 表象结束时再按键。两次按键的时间间隔为表象时间, 期间屏幕显示黑屏, 且被试闭上眼睛。表象结束后被试睁开眼睛, 屏幕呈现“休息”,休息时间为4~8 s伪随机(见图1B)。休息结束后呈现下一个“+”, 预示着下一个试次的开始。休息阶段用作fMRI数据分析的基线条件。
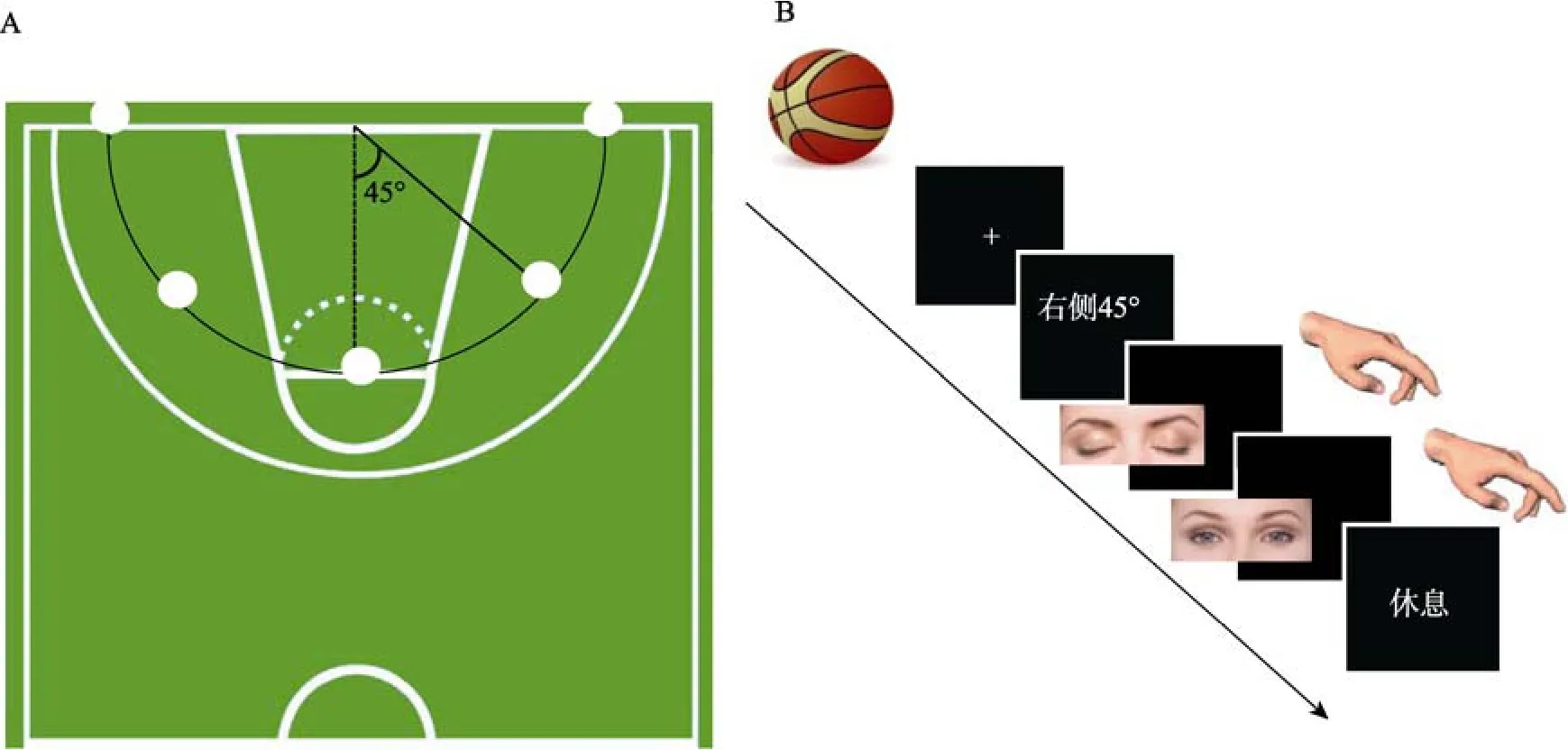
图1 实验设计示意图
2.5 数据采集与分析
2.5.1 行为数据采集与分析
采集fMRI数据时, 使用E-prime 2.0软件同步采集表象时间。将表象时间与实际执行时间的差值定义为运动表象与实际执行的时间一致性, 采用2(组别: 篮球运动员、新手) × 2(持球条件: 持球、不持球)的两因素重复测量方差分析考察不同持球条件下两组被试的时间一致性是否存在统计学差异。
正式实验前使用一般表象能力问卷考察两组被试的一般表象能力是否存在差异。该问卷使用7点评分(7代表最清晰), 包括8个项目, 分别评估一般动觉(1, 3, 5, 7)和视觉(2, 4, 6, 8)表象能力。正式实验后使用自编表象策略问卷和改编的运动表象问卷(Movement Imagery Questionnaire, MIQ-R;Hall & Martin, 1997), 对被试动觉表象策略和表象质量进行评估。表象策略问卷使用5点评分(5代表最积极), 包括4个项目, 分别询问被试是否使用了第一人称动觉表象, 使用该策略的容易程度, 是否能区分不同的持球条件, 以及区分的容易程度。运动表象问卷采用 7点评分(7代表最清晰), 包括 8个项目, 前4个评估表象的动觉特点, 后4个评估表象的视觉特点, 要求被试在持球和不持球条件下分别作答。为确保被试可以准确报告自己的主观感受, 被试回答问卷前先由主试讲解每个题目的意思直到被试充分理解。
对一般表象能力问卷和表象策略问卷得分采用非参数的Mann-Whitney U检验考察两组被试是否存在一般动觉表象能力、一般视觉表象能力及表象策略的差异。对运动表象问卷得分采用 Mann-Whitney U检验考察不同持球条件下的组间差异;采用 Wilcoxon符号秩检验考察不同持球条件间的表象质量评分是否存在组内差异。
2.5.2 fMRI数据采集、预处理与统计分析
采用西门子 3.0T全身磁共振成像仪(Siemens Magnetom Trio 3.0T)进行结构像与功能像数据的采集。采用 12导阵列线圈, 以平面回波成像(EPI)序列采集T2*功能像, 扫描参数:TR=2000 ms, TE=30 ms, 翻转角度为 90°, 层厚 4 mm, 视野 240 mm ×240 mm, 体素大小为 3.3 mm × 3.3 mm × 4 mm, 共33层, 逐层扫描。
统计分析前使用DPARSF V2.0 软件包(Yan &Zang, 2010)对功能数据进行预处理。首先进行时间校正去除同一时间点图像因采集时间差异引起的误差, 接着进行头动校正, 排除头部平动大于1 mm、转动大于 1.5°的被试对象; 然后将校正后的图像进行标准化, 采用SPM 自带EPI 模板, 归一化到MNI(Montreal Neurological Institute)坐标系统, 并将每个体素重采样至3 mm × 3 mm × 3 mm; 最后使用高斯核函数进行平滑, 降低空间噪声, 全宽半高值为6 mm。
为了避免按键动作对表象任务带来数据污染,每次表象的前和后各500 ms的扫描图像未纳入统计分析。在 MATLAB 7.1 (The Mathworks, Inc.,Natick, MA, USA)中加载SPM8 (http://www.fil.ion.ucl.ac.uk/spm)对功能数据进行了基于广义线性模型(general linear model, GLM) (Friston, Holmes,Price, Büchel, & Worsley, 1999)的分析。首先进行个体水平的一阶分析, 对每个被试运动表象时的大脑活动进行激活区检测, 并对图像进行二阶分析。
基于GLM构建基于双因素方差分析的2(组别:篮球运动员和新手组) × 2(持球条件: 持球和不持球)的F检验, 通过对比(contrast)的方式考察运动技能水平对运动表象的影响和躯体感觉输入对运动表象的影响以及运动技能水平与躯体感觉输入的交互效应。以交互作用脑区的极值点坐标为圆心,以6 mm为半径作球形ROI, 使用REST软件 (http://resting-fmri.sourceforge.net) 提取每个被试在各 ROI内的平均信号值进行参数比较。计算各ROI内持球和不持球条件平均信号值的差值以及持球和不持球条件下表象质量评分的差值。对信号值差值和表象质量评分差值进行相关分析, 考察持球对运动表象的影响。F检验结果未进行多重比较校正; 相关分析以p< 0.05为有显著统计学意义。
3 结果
3.1 行为结果
两组被试实际执行和运动表象的时间如表1所示。对运动表象与实际执行的时间一致性的分析发现组别主效应显著(F(1, 22)=5.42,p=0.029, η2=0.198), 两种持球条件下篮球运动员的时间一致性均显著优于新手(见图2)。

表1 实际执行和运动表象平均时间(秒)和标准差

图2 运动想象与实际执行的时间一致性(M ± SD)
两组被试(篮球运动员: 14.9±2.5, 新手: 14.3±2.5)在表象策略问卷上的得分无统计学差异(Z=0.65,p=0.52), 这说明所有被试都较好地遵从了实验要求, 即采用了动觉表象。
一般表象能力测试结果显示两组的一般动觉表象能力无统计学差异(Z=0.69,p=0.485, 篮球运动员:M ± SD=5.97 ± 0.55; 新手:M±SD=6.1±0.52); 两组的一般视觉表象能力无统计学差异(Z=1.13,p=0.26, 篮球运动员:M ± SD=5.86 ± 0.56;新手:M ± SD=6.04 ± 0.63)。
两种持球条件下篮球运动员的动觉表象得分均显著高于新手(持球条件下:Z=2.49,p=0.012,Φ=0.51; 不持球条件下:Z=2.64,p=0.008,Φ=0.54), 两组的视觉表象得分无统计学差异(持球条件下:Z=1.66,p=0.097; 不持球条件下:Z=1.57,p=0.117)。比较不同的持球条件发现, 除篮球运动员的视觉表象外, 两组被试在持球条件下的表象评分均显著高于不持球(动觉表象: 篮球运动员,Z=2.36,p=0.018,Φ=0.68; 新手,Z=2.02,p=0.044,Φ=0.58; 视觉表象: 篮球运动员,Z=1.75, p=0.081; 新手,Z=2.61,p=0.009,Φ=0.75) (见表 2)。

表2 不同持球条件下的运动表象平均得分和标准差
3.2 fMRI结果
3.2.1 全脑分析结果
全脑范围内两组在两种条件下运动表象引起的(运动表象vs.基线)激活增加的脑区如表3所示。篮球运动员在两种持球条件下都激活了额叶和顶叶皮层, 包括额中回/额下回、中央前回/辅助运动区和顶上小叶/顶下小叶。而新手只在持球条件下表现出相应激活; 在不持球条件下, 新手主要激活了左侧辅助运动区。
比较两组差异发现, 篮球运动员在额上回、额中回、额下回、额内侧回以及楔前叶表现出显著较大激活, 新手在辅助运动区和额上回表现出显著较大激活(见图3、表4)。
比较不同持球条件发现持球条件下表现出显著较大的基底神经节激活, 包括左侧苍白球和右侧壳核(见图4、表5)。
3.2.2 ROI分析结果
F检验的结果显示, 两侧中央后回, 右侧额下回以及左侧扣带回表现出组别与持球条件的交互作用(见表6)。参数估计揭示两组被试ROI内的平均信号值表现出相反的趋势: 篮球运动员持球条件下的平均信号值低于不持球, 而新手持球条件下的平均信号值高于不持球(见图5)。
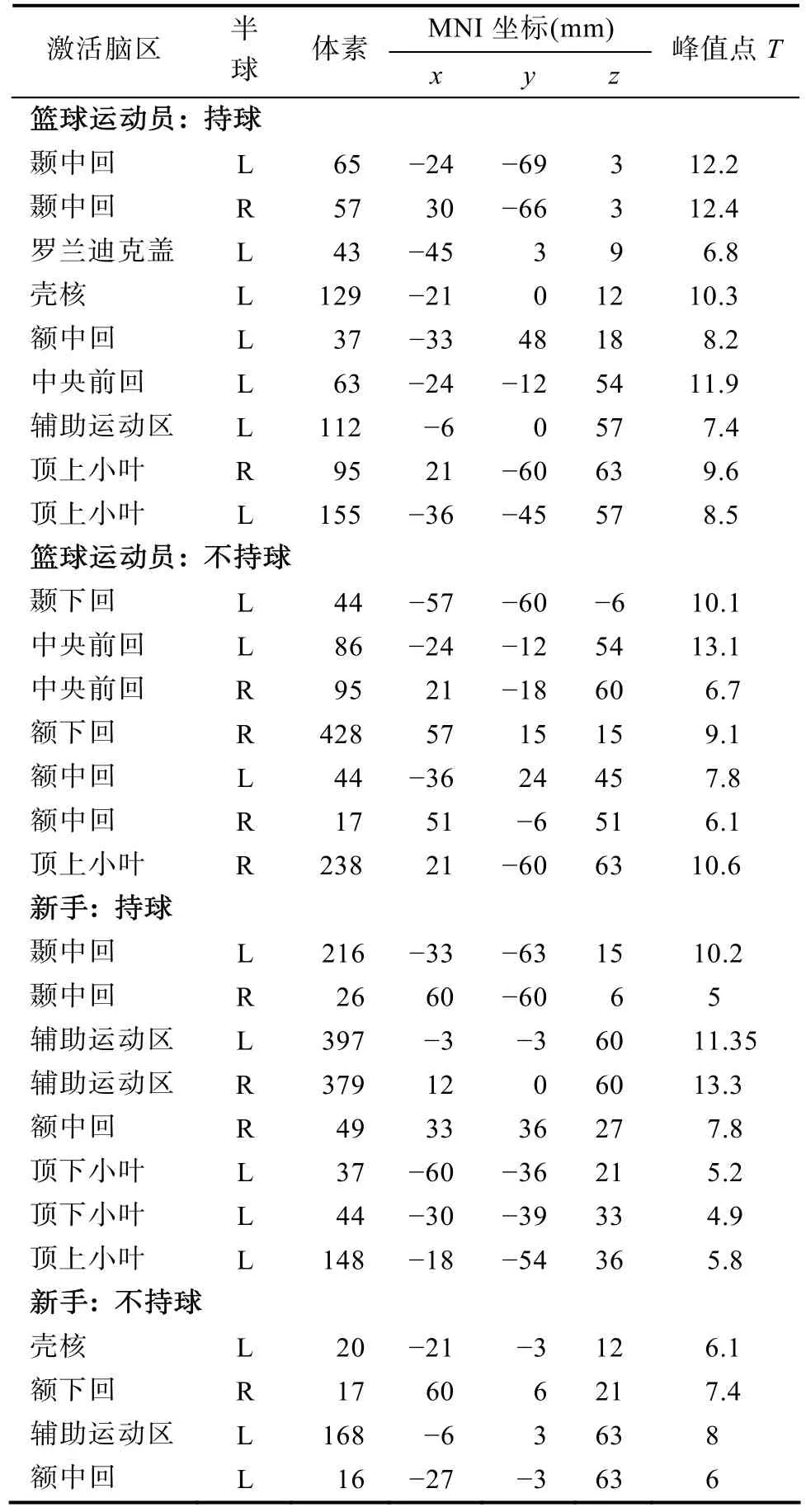
表3 不同持球条件下运动表象激活的脑区

表4 运动表象的组间差异

图3 运动表象的组间差异

图4 持球条件下激活较大脑区

表5 持球条件间差异的脑区

表6 组别与持球条件的交互脑区
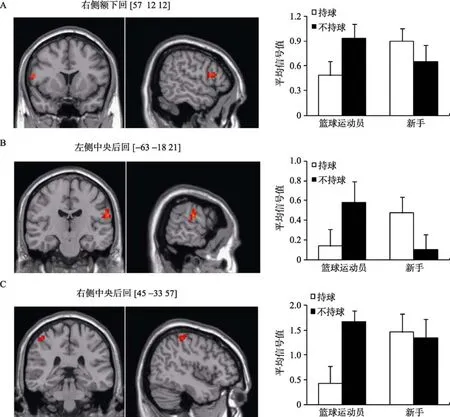
图5 组别与持球条件的交互作用脑区及其参数估计
3.2.3 ROI与表象质量评分的相关
对于篮球运动员, 持球与不持球条件下右侧额下回的平均信号值的差值与动觉表象质量评分的差值显著相关(r=−0.62,p=0.032) (见图6)。对于新手, 持球条件与不持球条件下交互脑区的平均信号值的差值与表象质量评分的差值未发现显著相关。两组的交互脑区平均信号值与视觉表象质量评分均未发现显著相关。
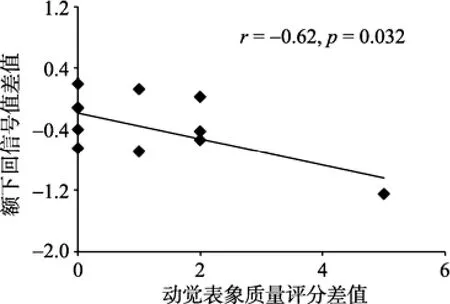
图6 持球对运动表象的影响
4 讨论
研究结果包括两个方面: 运动表象过程中运动技能水平越高, 时间一致性越好, 镜像神经系统激活越大; 躯体感觉输入调节不同运动技能水平被试运动表象过程中镜像神经系统的激活程度, 表现为有躯体感觉输入的情况下, 运动员镜像神经系统激活程度较低。
4.1 运动技能水平对运动表象的影响
器械依赖的技能学习往往伴随着大脑结构与功能的可塑性改变(Chang, 2014)。Draganski等人(2004)对普通人进行为期 3个月的杂技训练, 发现训练使负责加工和存储视觉空间运动的相关脑区灰质体积发生改变。本研究发现运动员较新手表现出显著较大的额下回激活(图3、表4), 额下回是镜像神经系统的重要组成部分(Abreu et al., 2012)。来自神经损伤案例的证据表明, 镜像神经系统受损使病人很难进行运动表象(Eisen, Lemon, Kiernan,Hornberger, & Turner, 2015), 这提示镜像神经系统参与运动表象过程。然而, 镜像神经系统的功能不是天生的, 而是经过大量的后天学习获得的(Catmur, Walsh, & Heyes, 2007)。已有研究发现键盘乐器演奏家和新手想象演奏乐器时都激活了镜像神经系统, 然而演奏家表现出显著较大的激活(Harris & de Jong, 2014)。以运动员为被试的研究也发现了类似的结果。优秀高尔夫运动员表象瞄准−挥杆动作系列时具有不同于新手的脑激活模式, 表现为高尔夫运动员在顶上小叶和运动前区表现出显著较大激活, 而新手激活了较为广泛的皮层下脑区(Milton, Solodkin, Hluštík, & Small, 2007)。镜像神经元的激活和完成动作的熟练程度相关(Buccino,Binkofski, & Riggio, 2004), 而运动表象与运动执行的神经基础具有重叠性(Dechent et al., 2004), 因此在运动想象时专家较新手表现出显著较大的镜像神经元的激活。在本研究中, 篮球运动员报告了较高的动觉表象质量, 同时表现出显著较大的镜像脑区激活, 这与已有研究发现一致。
运动员还激活了包括额上回和额中回在内的前额叶皮层。前额叶皮层按照动作预期结果进行快速动作序列的编排(Tucker, Luu, & Pribram, 1995;Shima, Isoda, Mushiaka, & Tanji, 2007), 无前额叶皮层参与的动作表现将产生组织紊乱的现象(Braun et al., 1998)。此外, 运动员还表现出较大的楔前叶激活。楔前叶与视觉空间功能有关, 在动作执行或想象中负责将注意指向特定空间(Cavanna &Trimble, 2006), 并通过协调注意在空间目标之间的转换, 使动作顺利执行(Oshio et al., 2010)。这提示运动员可能按照任务要求编排整套动作序列, 在拍球阶段将注意集中于球, 而在投篮阶段则转移到瞄准篮筐, 从而保证投篮任务的流畅执行。同时,行为结果显示两种持球条件下篮球运动员的运动表象与实际执行的时间一致性均显著优于新手。值得注意的是, 两组的一般运动表象能力并无显著性差异, 然而篮球运动员的动觉表象质量评分显著高于新手。这些发现共同说明篮球运动员具有较强的专项动作动觉表象能力, 这种能力与长期的专项训练有关。
本研究发现新手表现出显著较大的辅助运动区激活。辅助运动区参与运动计划, 主要与自我启动而非外部诱发的动作序列有关(Cunnington,Windischberger, Deecke, & Moser, 2003)。一种可能的解释是新手尝试通过调用动作序列相关的记忆来完成动作准备, 由其行为表现可知这种尝试效果不佳, 这可能因为短期的练习不足以使新手形成稳固的动作技能和运动图式。
4.2 躯体感觉输入对运动表象的影响
不同于无器械运动项目, 器械依赖的运动项目中效应器与运动器械(如手与篮球)之间的相互作用对运动技能的掌握至关重要。本研究发现, 与不持球条件相比, 持球条件主要激活与运动控制相关的基底神经节区域(图4、表5); 而篮球运动员和新手在不同持球条件下额下回和中央后回激活程度存在交互效应(表6)。ROI分析发现, 篮球运动员在持球条件下的平均信号值低于不持球(图 5), 并且持球与不持球条件下右侧额下回平均信号值的差值与动觉表象质量评分的差值显著相关(图 6); 而新手则表现出相反趋势, 即在持球条件下的平均信号值高于不持球。
感官反馈的研究发现, 如果在技能学习初期借助了感官反馈, 则在熟练阶段仍然需要相应感官反馈的辅助才能完成对技能经验的有效调用(Proteau,Tremblay & DeJaeger, 1998; Tremblay & Proteau,1998)。对于篮球运动员, 篮球的躯体感觉输入伴随着运动训练的始终, 长期专项训练使篮球运动员将球有机地纳入运动模式和身体表征中。中央后回是初级躯体感觉中枢, 主要表征触觉。结果显示持球条件下中央后回的激活低于不持球, 与中央后回的激活模式相似, 持球条件下额下回的激活低于不持球。这可能是由于持球条件下的表象任务与实际训练中的运动状态匹配, 篮球运动员需要较少心理资源即可激活已有运动模式, 任务难度相对较低, 从而相对自动化地完成投篮的动作序列。也就是说,篮球运动员动用较少的神经表征就能达到较好的表象效果, 这与相关分析得到的持球条件下表象质量评分与额下回激活呈显著负相关的结果一致。在不持球条件下, 篮球运动员需要动用较多的资源弥补躯体感觉信息缺失带来的运动模式失匹配, 任务难度增加, 表现为镜像神经系统和运动控制相关脑区的较大激活。这与已有研究发现一致, 即对于熟练掌握的动作, 动作难度小, 心理资源耗费少, 同时还能保持较优的动作表现(Bakker et al., 2008)。对新手来说, 投篮动作的掌握水平较低, 实验前为期3周的实地训练尚不能使新手将球纳入运动模式和身体表征, 表现为ROI脑区的激活模式与运动员不同。
需要指出的是, 持球带来的躯体感觉输入影响中央后回和额下回的活动。然而本研究的结果还不足以揭示这两个脑区之间的相互关系, 探讨躯体感觉输入情况下中央后回与额下回的关系需要新的研究数据支持。
4.3 躯体感觉输入对篮球运动员运动表象影响的神经生理机制
运动表象的神经生理研究中, 多采用fMRI或者 TMS对大脑皮层活动进行分析以推测运动表象导致的中枢神经系统变化。fMRI通过大脑血氧依赖水平BOLD (blood oxygen level dependent)信号来分析大脑的活动, TMS通过给予运动皮层一个瞬间的磁场刺激导致的 MEP振幅变化来反映运动皮层的兴奋性。
在TMS研究中, 已有研究(Wang et al., 2014)发现在表象羽毛球发球动作时, 运动员较新手运动皮层兴奋性增高, 并且持器械条件较不持器械条件下增高得更大。运动皮层兴奋性的这种改变可能与它的功能有关。运动皮层主要负责迅速、准确地输出运动程序, 而运动皮层兴奋性越高, 越能满足运动的要求。运动员在持器械下表象发球动作, 较不持器械条件更具生态效度, 因此表现出更高的皮层兴奋性。
使用fMRI, 本研究发现在表象篮球投篮时,运动员较新手镜像神经系统兴奋性增高, 但是外部感觉输入对不同技能水平被试产生了不同的影响。运动员在持器械条件下表象投篮任务时, 较不持球更符合实际运动训练情景, 表象难度下降, 表现在镜像神经系统激活程度下降; 而新手相反。Bakker等(2008)发现运动表象任务难度与镜像神经系统激活成正比, 本研究结果与该研究一致。另外, 运动表象过程中镜像神经系统激活程度的变化也可以反映长期运动训练导致的大脑可塑性改变。镜像神经系统的任务是根据感觉信息进行决策、判断, 并可以调节运动控制相关脑区的兴奋性。在持器械条件下, 运动员运动皮层兴奋性升高幅度更大, 而镜像神经系统可能不需要更大的兴奋性改变, 就可以精确地触发运动程序。
5 结论
综上所述, 本研究结果表明: 与新手相比, 运动专家进行动觉表象时具有专家优势, 表现为动觉表象质量较好, 镜像神经系统激活较高; 对运动专家来说, 躯体感觉输入对运动表象有调节作用, 表现为持球条件下运动表象质量较好, 镜像神经系统激活较低, 不持球条件反之。同时, 与躯体感觉输入相关的中央后回活动发现类似趋势。这提示持球条件下表象任务与实际训练中的运动状态匹配, 运动专家需要较少心理资源即可激活已有运动模式。
研究发现器械带来的躯体感觉输入可以显著提高运动员的表象质量, 且长期的器械使用可以使镜像神经系统产生可塑性改变。研究启示提高运动技能和促进运动康复时要结合相应器械的使用。
Abreu, A.M., Macaluso, E., Azevedo, R.T., Cesari, P., Urgesi,C., & Aglioti, S.M.(2012).Action anticipation beyond the action observation network: A functional magnetic resonance imaging study in expert basketball players.European Journal of Neuroscience, 35, 1646–1654.
Bakker, M., De Lange, F.P., Helmich, R.C., Scheeringa, R.,Bloem, B.R., & Toni, I.(2008).Cerebral correlates of motor imagery of normal and precision gait.NeuroImage,41, 998–1010.
Braun, A.R., Balkin, T.J., Wesensten, N.J., Gwadry, F.,Carson, R.E., Varga, M.,...Herscovitch, P.(1998).Dissociated pattern of activity in visual cortices and their projections during human rapid eye movement sleep.Science, 279, 91–95.
Buccino, G., Binkofski, F., Fink, G.R., Fadiga, L., Fogassi, L.,Gallese, V.,...Freund, H.J.(2001).Action observation activates premotor and parietal areas in a somatotopic manner: An fMRI study.European Journal of Neuroscience,13, 400–404.
Buccino, G., Binkofski, F., & Riggio, L.(2004).The mirror neuron system and action recognition.Brain and Language,89, 370–376.
Catmur, C., Walsh, V., & Heyes, C.(2007).Sensorimotor learning configures the human mirror system.Current Biology, 17, 1527–1531.
Cavanna, A.E., & Trimble, M.R.(2006).The precuneus: A review of its functional anatomy and behavioural correlates.Brain, 129, 564–583.
Chaminade, T., Meltzoff, A.N., & Decety, J.(2005).An fMRI study of imitation: Action representation and body schema.Neuropsychologia, 43, 115–127.
Chang, Y.(2014).Reorganization and plastic changes of the human brain associated with skill learning and expertise.Frontiers in Human Neuroscience, 8, 35.
Cocks, M., Moulton, C.A., Luu, S., & Cil, T.(2014).What surgeons can learn from athletes: Mental practice in sports and surgery.Journal of Surgical Education, 71, 262–269.
Cunnington, R., Windischberger, C., Deecke, L., & Moser, E.(2003).The preparation and readiness for voluntary movement: A high-field event-related fMRI study of the Bereitschafts-BOLD response.NeuroImage, 20, 404–412.
de Vries, S., Tepper, M., Otten, B., & Mulder, T.(2011).Recovery of motor imagery ability in stroke patients.Rehabilitation Research and Practice, 2011, 283840.
Decety, J., Perani, D., Jeannerod, M., Bettinardi, V., Tadary,B., Woods, R.,...Fazio, F.(1994).Mapping motor representations with positron emission tomography.Nature,371, 600–602.
Dechent, P., Merboldt, K.D., & Frahm, J.(2004).Is the human primary motor cortex involved in motor imagery?Cognitive Brain Research, 19, 138–144.
Draganski, B., Gaser, C., Busch, V., Schuierer, G., Bogdahn,U., & May, A.(2004).Neuroplasticity: Changes in grey matter induced by training.Nature, 427, 311–312.
Eisen, A., Lemon, R., Kiernan, M.C., Hornberger, M., &Turner, M.R.(2015).Does dysfunction of the mirror neuron system contribute to symptoms in amyotrophic lateral sclerosis?Clinical Neurophysiology, 126, 1288–1294.
Fogassi, L., Ferrari, P.F., Gesierich, B., Rozzi, S., Chersi, F.,& Rizzolatti, G.(2005).Parietal lobe: From action organization to intention understanding.Science, 308,662–667.
Friston, K.J., Holmes, A.P., Price, C.J., Büchel, C., &Worsley, K.J.(1999).Multisubject fMRI studies and conjunction analyses.NeuroImage, 10, 385–396.
Greenberg, D.L., & Knowlton, B.J.(2014).The role of visual imagery in autobiographical memory.Memory & Cognition,42, 922–934.
Grèzes, J., Frith, C., & Passingham, R.E.(2004).Brain mechanisms for inferring deceit in the actions of others.Journal of Neuroscience, 24, 5500–5505.
Hall, C.R., & Martin, K.A.(1997).Measuring movement imagery abilities: A revision of the movement imagery questionnaire.Journal of Mental Imagery, 21, 143–154.
Harris, R., & de Jong, B.M.(2014).Cerebral activations related to audition-driven performance imagery in professional musicians.PLoS One, 9, e93681.
Jin, H., Xu, G.P., Zhang, J.X., Gao, H.W., Ye, Z.E., Wang,P.,...Lin, C.D.(2011).Event-related potential effects of superior action anticipation in professional badminton players.Neuroscience Letter, 492, 139–144.
Kasess, C.H., Windischberger, C., Cunnington, R., Lanzenberger,R., Pezawas, L., & Moser, E.(2008).The suppressive influence of SMA on M1 in motor imagery revealed by fMRI and dynamic causal modeling.NeuroImage, 40, 828–837.
Keller, P.E.(2012).Mental imagery in music performance:Underlying mechanisms and potential benefits.Annals of the New York Academy of Sciences, 1252, 206–213.
Lotze, M., & Cohen, L.G.(2006).Volition and imagery in neurorehabilitation.Cognitive and Behavioral Neurology,19, 135–140.
Milton, J., Small, S.L., & Solodkin, A.(2008).Imaging motor imagery: Methodological issues related to expertise.Methods, 45, 336–341.
Milton, J., Solodkin, A., Hluštík, P., & Small, S.L.(2007).The mind of expert motor performance is cool and focused.NeuroImage, 35, 804–813.
Mizuguchi, N., Sakamoto, M., Muraoka, T., Nakagawa, K.,Kanazawa, S., Nakata, H.,...Kanosue, K.(2011).The modulation of corticospinal excitability during motor imagery of actions with objects.PLoS One, 6, e26006.
Oldfield, R.C.(1971).The assessment and analysis of handedness: The Edinburgh inventory.Neuropsychologia,9, 97–113.
Oshio, R., Tanaka, S., Sadato, N., Sokabe, M., Hanakawa, T.,& Honda, M.(2010).Differential effect of double-pulse TMS applied to dorsal premotor cortex and precuneus during internal operation of visuospatial information.NeuroImage, 49, 1108–1115.
Papaxanthis, C., Pozzo, T., Skoura, X., & Schieppati, M.(2002).Does order and timing in performance of imagined and actual movements affect the motor imagery process?The duration of walking and writing task.Behavioural Brain Research, 134, 209–215.
Proteau, L., Tremblay, L., & DeJaeger, D.(1998).Practice does not diminish the role of visual information in on-line control of a precision walking task: Support for the specificity of practice hypothesis.Journal of Motor Behavior,30, 143–150.
Rizzolatti, G., & Craighero, L.(2004).The mirror-neuron system.Annual Review of Neuroscience, 27, 169–192.
Shima, K., Isoda, M., Mushiake, H., & Tanji, J.(2007).Categorization of behavioural sequences in the prefrontal cortex.Nature, 445, 315–318.
Solodkin, A., Hlustik, P., Chen, E.E., & Small, S.L.(2004).Fine modulation in network activation during motor execution and motor imagery.Cerebral Cortex, 14, 1246–1255.
Sposito, A., Bolognini, N., Vallar, G., & Maravita, A.(2012).Extension of perceived arm length following tool-use:Clues to plasticity of body metrics.Neuropsychologia, 50,2187–2194.
Tkach, D., Reimer, J., & Hatsopoulos, N.G.(2007).Congruent activity during action and action observation in motor cortex.Journal of Neuroscience, 27, 13241–13250.
Tremblay, L., & Proteau, L.(1998).Specificity of practice:The case of powerlifting.Research Quarterly for Exercise and Sport, 69, 284–289.
Tucker, D.M., Luu, P., & Pribram, K.H.(1995).Social and emotional self-regulation.Annals of the New York Academy of Sciences, 769, 213–240.
Vargas, C.D., Olivier, E., Craighero, L., Fadiga, L., Duhamel,J.R., & Sirigu, A.(2004).The influence of hand posture on corticospinal excitability during motor imagery: A transcranial magnetic stimulation study.Cerebral Cortex, 14, 1200–1206.
Wang, Z., Wang, S., Shi, F.Y., Guan, Y., Wu, Y., Zhang, L.L.,...Zhang, J.(2014).The effect of motor imagery with specific implement in expert badminton player.Neuroscience,275, 102–112.
Wu, Y., Zeng, Y., Zhang, L., Wang, S., Wang, D., Tan, X.,...Zhang, J.(2013).The role of visual perception in action anticipation in basketball athletes.Neuroscience, 237, 29–41.
Yan, C.G., & Zang, Y.F.(2010).DPARSF: A MATLAB toolbox for "pipeline" data analysis of resting-state fMRI.Frontiers in Systems Neuroscience, 4, 13.
