不同外源激素对香草兰花芽分化的影响
2017-01-21王辉顾文亮庄辉发赵青云朱自慧宋
王辉+顾文亮+庄辉发+赵青云+朱自慧+宋应辉
摘 要 为探究香草兰花芽分化调控措施及相关机理,在花芽分化前期,以分别喷施细胞分裂素(6-BA)、脱落酸(ABA)、生长素(IAA)和赤霉素(GA)为处理,以清水为对照,分析各处理条件下功能叶内源激素和营养物质含量变化,并统计各处理对花芽分化率的影响。结果表明:6-BA和ABA处理均能增加功能叶中玉米素核苷(ZR)和ABA含量,提高其在花芽分化过程中的蛋白质、可溶性糖和淀粉含量;IAA处理能提高花芽分化中后期功能叶IAA、ABA、ZR和 GA含量,却降低整个分化过程中蛋白质、可溶性糖和淀粉的含量;GA处理降低了ZR和ABA含量,对蛋白质和淀粉含量影响不大;除GA处理外,6-BA、ABA、IAA处理均在一定程度上提高了花芽分化率,以6-BA最为显著,可参考在生产上施用。
关键词 香草兰 ;花芽分化 ;外源激素 ;内源植物激素
中图分类号 Q946.885 文献标志码 A Doi:10.12008/j.issn.1009-2196.2016.11.015
Abstract To investigate the regulation measures on vanilla flower bud formation and related mechanism, the plants were sprayed with water(CK), 6-Benzylaminopurine(6-BA), abscisic aci(ABA), auxin(IAA) and gibberellin(GA)individually before the onset of floral induction. The results showed that the treatment of 6-BA increased the contents of zeatin riboside(ZR), ABA , protein, soluble sugar and starch of functional leaves , and so did the treatment of ABA. The treatment of IAA increased the hormone contents of IAA, ABA, ZR and GA in the late stages of floral differentiation, but it reduced the contents of protein, soluble sugar and starch at full flowering stage. Spraying GA reduced the contents of ZR and GA. The treatments of 6-BA, ABA, IAA could enhance the ratio of flower bud formation, 6-BA treatment got the most significant effect, which could be applied in production.
Keywords Vanilla planifolia Andrews ; Flower bud differentiation ; Plant external hormones ; Endogenous plant hormones
香草兰(Vanilla planifolia Andrews)是兰科(Orchidaceae)香草兰属(Vanilla)多年生热带藤本植物。其鲜豆荚加工后被广泛用于调制各种高级香烟、名酒、特级茶叶、高档香水和糕点,被誉为“天然食品香料之王”。全世界香草兰总面积约8万hm2,鲜豆荚年产量约9 000 t[1]。我国海南、云南和四川等地均有一定面积的种植。
作为热带地区附加值较高的作物之一,产量不稳定在一定程度上制约了香草兰产业的快速发展。有研究表明,相对于授粉成功率、保荚率和单荚重等产量构成因素,花芽分化对香草兰产量的影响是最大、最实际的[2]。因此,不少科研工作者开展了相关研究,对香草兰花芽分化时期、过程中物质累积和内源激素变化等生理特性做了探讨和分析[3-4],而关于其花芽分化调控的研究尚未见报道。本文拟通过研究不同外源激素对香草兰花芽分化及相关生理生化特性的影响,以期为探索促花栽培措施提供理论依据和技术方法。
1 材料与方法
1.1 材料
试验在中国热带农业科学院香料饮料研究所高龙试验基地进行,供试植株为2012年定植的‘热引3号香草兰,所有供试的香草兰植株均为荫蔽条件下露地栽培。
1.2 方法
1.2.1 处理与采样
设3个试验小区,各小区内随机设置处理,每个处理选30株(15条攀缘柱)长势良好、大小一致的香草兰为供试植株,每株标记8~10个节间为取样和观测茎段,于2015年12月4日(花芽分化前)进行不同外源激素喷施处理。处理1(T1)喷施100 mg/L的6-BA(T1),处理2(T2)喷施100 mg/L的ABA(T2),处理3(T3)喷施100 mg/L的IAA(T3),处理4(T4)喷施100 mg/L的GA(T4)[5],清水对照,喷施程度以全株叶片滴水为止。激素喷施处理后第20天开始,每隔13~15 d取样,分别于花芽分化前期(2015年12月25日及2016年1月11日)、花芽分化中期(2016年1月25日及2016年2月5日)、花芽分化后期(2016年2月19日及2016年3月5日)共6次从标记茎段采集功能叶,经液氮处理后,-80℃超低温冰箱中保存。于 2016 年4月6日分别统计各处理花芽数量并计算花芽分化率。
1.2.2 测定方法
样品中IAA、GA、ZR和ABA含量采用酶联免疫吸附测定法(ELISA)测定[6],植物激素含量测定试剂盒购自中国农业大学;蛋白质含量测定用考马斯亮蓝 G-250比色法;可溶性糖和淀粉含量测定采用硫酸-蒽酮比色法[7];开花中后期统计标记茎段的花芽数和总芽数,每个重复统计3株,计算各重复平均花芽分化率。
1.2.3 数据处理与统计方法
采用Excel 2010软件处理实验数据与图表,采用SPSS 20.0进行显著性分析。
2 结果与分析
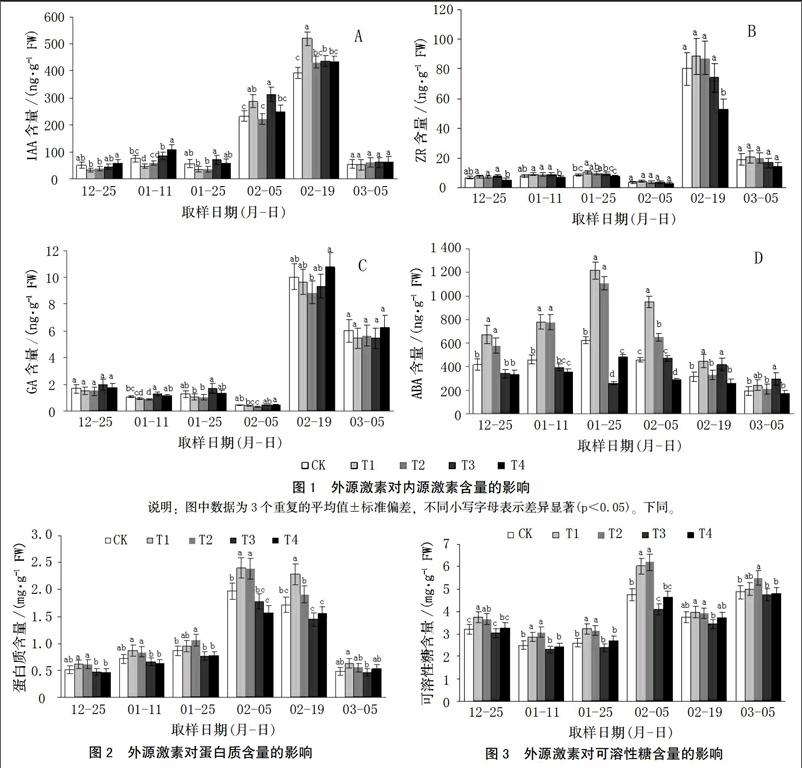
2.1 外源激素对香草兰内源激素含量的影响
由图1可知,与对照相比,T1和T2处理降低了香草兰花芽分化前期叶片内IAA含量,并降低了整个分化过程中叶片内GA含量,其中T2处理条件下GA含量在花芽分化中期显著低于对照;T1和T2处理提高了整个花芽分化过程中ZR和ABA含量,其中ABA含量提高显著,在高峰期比对照分别高94.12%、77.28%。
在花芽分化前期,T3处理叶片内IAA和ABA含量低于对照,而在花芽分化中后期,该处理条件下两种内源激素的含量又高于对照,且后期(2016年2月19日)达显著水平,IAA和ABA含量分别比对照高11.26%和32.22%。图1-B、图1-C表明,相对于对照,T3处理条件下ZR和GA含量在花芽分化前期有所提高而后期有所降低。
纵观整个花芽分化过程,T4处理提高了各个时期功能叶IAA和GA含量,但总体上差异不显著;与对照相比,T4处理降低了ZR和ABA含量,在花芽分化中后期高峰时表现显著,分别降低34.32%和37.63%。
2.2 外源激素对香草兰营养物质含量的影响
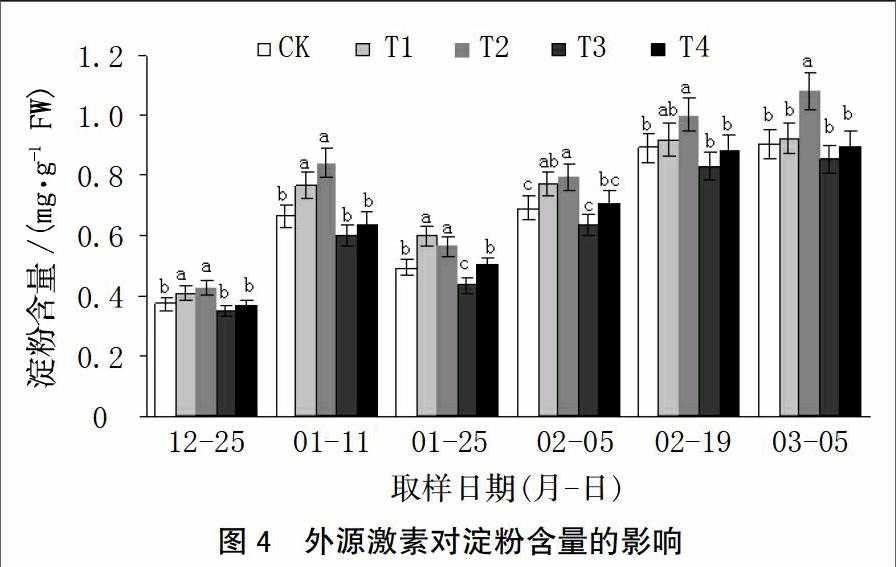
2.2.1 对蛋白质含量的影响
由图2可知,T1处理能提高各时期叶片蛋白质含量,6次取样分别比对照高19.18%、21.20%、9.22%、22.09%、33.12%和29.17%,中后期(2月5日和2月19日)提高显著;T2处理也能提高各时期叶片蛋白质含量,中期(1月25日和2月5日)比对照分别显著提高22.21%和21.12%;T3处理降低了各时期蛋白质含量;T4处理降低了前5次取样叶片中蛋白质含量,显著时(2月5日)比对照降低20.30%。
2.2.2 对可溶性糖含量的影响
由图3可知,T1和T2处理能提高叶片可溶性糖含量,且前中期提高显著,高峰期分别比对照提高19.76%和30.57%;T3处理各时期叶片可溶性糖含量均低于对照,高峰期比对照降低13.95%;T4处理可溶性糖含量也略高于对照。
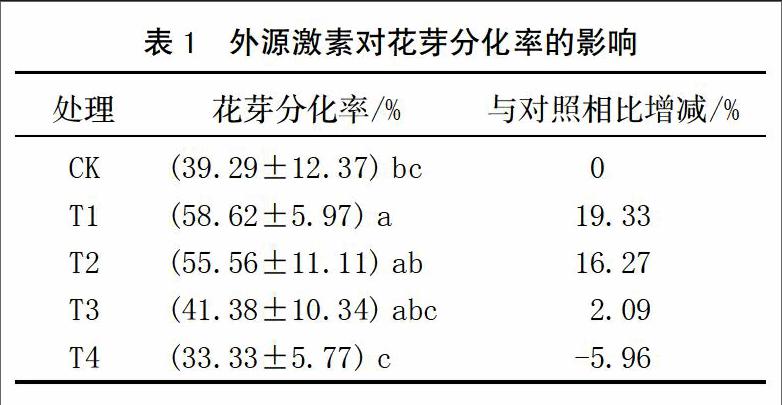
2.2.3 对淀粉含量的影响
与对照相比,T1、T2处理能提高各时期叶片淀粉含量,T3处理则相反,而T4处理对淀粉含量影响不大(图4)。其中,T1处理在前中期增加显著;T2处理在整个花芽分化过程中淀粉含量均显著高于对照、T3和T4处理;在花芽分化后期(3月5日)分别比对照、T3、和T4处理高19.49%、26.41%和20.32%。
2.3 外源激素对香草兰花芽分化率的影响
由表1可知,T1、T2和T3处理对香草兰花芽分化有促进作用。其中T1、T2处理花芽分化率达到58.62%和55.56%,比对照分别提高19.33%和16.27%;T3处理花芽分化率仅比对照提高2.09%;而T4处理花芽分化率降低了5.96%。方差分析结果表明:T1处理显著高于对照和T4处理,T2处理显著高于T4处理,其他处理之间差异不显著。
3 讨论与结论
本研究探讨了外源激素调控香草兰花芽分化的措施及相关机理。初步研究结果表明:在生产上施用一定浓度的6-BA可调控香草兰花芽分化。
花芽分化是作物在一定遗传背景控制下,从营养生长进入生殖生长的一个高度复杂的生理生化和形态建成过程。在这个过程中,作物内源激素含量变化和营养物质的积累运转均起着重要的调控作用[8]。利用外源激素是目前生产者为调节花期、保证花的质量而常采用的手段,但外源激素对作物生长发育的调控作用十分复杂。本研究通过施用几种外源激素,分析各处理对香草兰花芽分化影响及相关生理机制。结果表明:有利于花芽分化的6-BA和ABA处理都提高了香草兰花芽分化期功能叶ZR和ABA含量,并降低了其GA含量。说明在花芽生理分化期,ZR和ABA含量的提高有利香草兰成花,而GA对香草兰又是一种抑花激素。上述结果与前人研究一致[9-11]。
蛋白质是植物生长发育中重要的结构物质,其含量的高低是植物体总代谢水平的重要指标。许多研究表明,蛋白质的大量积累是成花的重要物质基础。碳水化合物的供应是否充足,对花芽分化质量优劣也起决定性作用[12]。本研究表明:6-BA和ABA 能提高功能叶蛋白质、可溶性糖和淀粉含量,说明上述处理加强了碳水化合物的调运和分配能力,有利于花芽分化;GA处理降低了功能叶蛋白质含量,从而在一定程度上降低了花芽分化率,亦符合前期研究结果[3]。
多数研究认为,IAA处理抑制花芽分化[13-15]。从本研究结果来看,IAA处理虽然也提高了花芽分化中后期内源IAA的含量,但对香草兰花芽分化率影响很小,这可能与生产管理上养分供应充足有关。
综上,外源激素处理影响了香草兰功能叶的内源激素和营养物质,它们均与花芽分化表现一定的相关性,然而同一生理信号在不同植物或同一植物不同时期的作用可能不同,例如有些生理信号在香草兰的成花转变中起促进作用,但可能抑制花的发育,因此,要了解不同激素对香草兰花芽分化的作用和机理,除了需进一步开展不同激素、浓度和配比处理实验外,还应深入开展分子水平和信号转导方面的研究,以便从不同角度、不同层次来揭示其机理。
参考文献
[1] 顾文亮,陈娅萍,王辉,等. 不同蔬果处理下香草兰果荚脱落及其内源激素含量变化研究[J].热带作物学报,2015,36(3):551-556.
[2] 王 辉,庄辉发,王 华,等. 香草兰单株产量构成性状的通径分析[J]. 安徽农业科学,2010,38(29):16 198-16 199.
[3] 赵秋芳,陈娅萍,顾文亮,等. 香草兰花芽分化期蛋白质及碳水化合物变化研究[J]. 热带作物学报,2015,36(6):1 053-1 058.
[4] 赵秋芳,陈娅萍,顾文亮,等. 香草兰花芽分化期叶片矿质元素变化研究[J]. 热带农业科学,2015,35(2):8-12.
[5] 孟庆杰,王光全. 植物激素及其在农业生产中的应用[J].河南农业科学,2006,4(2):9-11.
[6] 孙宽莹,陈 彦. 植物内源激素的分析方法[J]. 湖北农业科学,2011,50(18):3 681-3 683,3 690.
[7] 李合生.植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2006:298.
[8] 何静茹,李振坚. 外源激素对美花石斛试管内花芽分化的影响[J]. 江苏农业科学,2015,43(11):71-73.
[9] 刘伦,王 超,姚改芳. 外源生长调节剂处理对‘满天红ב砀山酥梨后代成花的影响[J]. 南京农业大学学报,2015,38(3):381-388.
[10] 钱 桦,刘 燕,郑勇平,等. 施用6--BA对春石斛花芽分化及内源激素的影响[J]. 北京林业大学学报,2009,31(6):27-31.
[11] 李有梅,邢利博,张 东,等. 喷施IAA抑制富士幼树花芽孕育的机制[J]. 西北农业学报,2015,24(4):84 -89.
[12] 金亚征,姚太梅,丁丽梅,等. 果树花芽分化机理研究进展[J]. 北方园艺,2013(7):193-196.
[13] 秦建彬,魏翠华,余祖云,等. 大花蕙兰花芽分化与激素关系的研究[J]. 中国农学通报,2011,27(31):109-112.
[14] 王海波,赵君全,王孝娣,等. 新梢内源激素变化对设施葡萄花芽孕育的影响[J]. 中国农业科学,2014,47(23):4 695-4 705.
[15] 范茜茜,汪志辉,熊博,等. 几种调控措施对龙眼顶芽激素及成花坐果的影响[J]. 广西植物,2015,35(5):685-691.
