施肥对降香黄檀幼苗生长和光合的影响1)
2017-01-17王楠王宏信李向林苑洋马炳琛陈博臧公胜
王楠 王宏信 李向林 苑洋 马炳琛 陈博 臧公胜
(三亚学院,三亚,572022)
施肥对降香黄檀幼苗生长和光合的影响1)
王楠 王宏信 李向林 苑洋 马炳琛 陈博 臧公胜
(三亚学院,三亚,572022)
以2年生降香黄檀(DalbergiaodoriferaT. Chen)幼苗为研究对象,通过盆栽试验研究不同氮(N)、磷(P)、钾(K)配比下的施肥处理对苗木生长和光合的影响。结果表明:(1)施肥显著促进了降香黄檀幼苗的生长,苗高、地径、生物量均存在显著的差异(P<0.01),随着施肥量的增加,地径、苗高、总生物量、茎生物量、叶生物量和根生物量的增大幅度均表现出增大的趋势,并在N3P3K1(氮、磷、钾质量分别为1.8、0.6、0.3g)配比下出现最大值(根和叶生物量除外),然后增幅均呈递减趋势;施肥后茎生物量在总生物量中所占比例显著增大,而叶生物量和根生物量在总生物量中所占比例均显著减小(P<0.01);(2)施肥显著提高了降香黄檀幼苗的光合能力,随着肥量的增加,最大净光合速率(Pmax)增幅逐步增大,并在N3P3K1配比下达到最大值172.38 nmol·g-1·s-1,随后增幅呈递减趋势;Pmax、叶生物量和总生物量间均为正相关关系,光合能力的提高和叶生物量的增大是总生物量增大的原因。
降香黄檀;施肥;生物量;光合能力
A potted experiment with 2-year-old seedlings was conducted to study the growth (height, ground diameter, and biomass), biomass allocation, and leaf photosynthesis ofDalbergiaodoriferaunder different N, P, K contents at four levels with orthogonal design. The results showed that: (1)Fertilization had obvious promoting effect on the growth ofD.odoriferaseedlings, and the height, ground diameter, total biomass, stem biomass, leaf biomass and root biomass had significant differences and were increased with the increase of fertilizer amount before N3P3K1combination (N3:1.8 g, P3:0.6 g, K3: 0.3 g) (leaf biomass and root biomass excepted); the proportion of stem biomass to total biomass was significantly and positively correlated with total biomass after fertilization, while the leaf biomass and root biomass showed opposite trends with total biomass (P<0.01); (2)The photosynthetic capacity ofD.odoriferawas significantly increased after fertilization and the light-saturated net photosynthetic rate (Pmax) increased with the increasing fertilizer amount and peaked at N3P3K1combination (172.38 nmol·g-1·s-1); total biomass was significantly and positively correlated withPmaxand leaf biomass. The enhanced photosynthetic capacity and increased leaf biomass contributed to the increase of the total biomass.
降香黄檀(DalbergiaodoriferaT. Chen)是蝶形花科黄檀属植物,为常绿半落叶乔木树种,是国家二级保护野生植物。降香黄檀的心材质地坚硬,花纹美观,抗压耐腐,是制造高档家具、乐器和工艺品的上等材料。此外,从其心材中提取的降香油气味清香,是优质香料的定香剂,同时具理气止痛等医疗功效。由于降香黄檀具有较高的经济价值,导致其被过度砍伐,天然资源日益枯竭。因此,发展人工培育种植降香黄檀是保护该树种和增加市场供给的必然趋势。由于天然降香黄檀生长的石灰岩质山区土壤瘠薄,导致该树种生长缓慢,心材形成时间长[1]。有研究表明,降香黄檀心材的形成起始胸径为8~10 cm,树木需要生长6年以上才能达到该值,完全成才需要30~40年[2]。因此,如何加快树木生长速率和缩短心材形成时间是目前人工栽植降香黄檀所面临的主要问题之一[1,3-4]。
施肥是人工促进树木生长的主要方法。大量研究表明,在一定范围内,氮、磷、钾肥料的增加对树木生长具有促进作用,而超出该范围的施肥会对树木的生长造成伤害[5-7]。在降香黄檀的育苗工作中,由于没有一个合适的施肥方法而经常出现苗木生长缓慢和肥料浪费等现象[4,8-9]。因此,研究氮、磷、钾元素的施加量及其配比对苗木生长的影响,对今后的生产实践工作具有重要的指导意义。此外,随着施肥量的增加,树木的总生物量呈增大趋势,然而生物量在各器官中的分配格局及其机制还不是很清楚[7]。有研究认为,随着土壤养分含量的变化,叶量和光合速率均会发生改变[5,10-11],进而影响生物量的累计和分配,然而,两者间的关系及对生物量影响程度还不清楚。本研究以降香黄檀幼苗为对象,研究不同氮、磷、钾添加对幼苗生长和光合的影响,主要有以下两个目的:(1)确定最佳元素组合,为合理人工培育降香黄檀提供科学依据;(2)探讨施肥对降香黄檀生物量分配格局的影响及其机制。
1 材料与方法
1.1 试验材料
试验于2015年4—11月进行,样地位于海南省三亚学院校内实验基地。供试降香黄檀幼苗为2年生实生苗,2015年4月选取健壮且生长一致的幼苗(平均苗高50.6 cm)移植到塑料盆(规格为上口径21.0 cm,底16.0 cm,高19.5 cm)中培养。每盆装土4.0 kg。试验所用土壤取自试验林地,消毒后风干、碾碎装盆。测得土壤pH值为4.85,有效氮、有效磷和有效钾质量分数分别为112、37、76 mg·kg-1。幼苗定植后缓苗2周后进行施肥处理。氮、磷、钾肥分别为尿素(含氮46%)、钙镁磷肥(含P2O514%)、硫酸钾肥(含K2O 50%)。
1.2 试验设计
每种元素施肥量按照4.0 kg土壤中该元素总含量的100%(N1、P1、K1分别为0.45、0.15、0.30 g)、200%(N2、P2、K2分别为0.90、0.30、0.60 g)、400%(N3、P3、K3分别为1.8、0.6、1.2 g)和800%(N4、P4、K4分别为3.6、1.2、2.4 g)进行添加,分别在5、7、9月的月初进行施肥。试验采用3因素4水平正交设计(表1),共16个处理,每个处理6个重复,对照记为N0P0K0,设20个重复。施肥处理6个月后进行生长测定。

表1 降香黄檀苗木施肥试验设计
1.3 生长测定
施肥处理前后,均用直尺和游标卡尺测定所有苗木的树高和地径。施肥后,每种处理选取3株苗木进行生物量测定,将幼苗用去离子水洗净,分根、茎、叶分别称其鲜质量,置于烘箱中65 ℃下烘干至恒质量(0.000 1 g),测定生物量。
1.4 光合测定
在2015年的5—10月的每月中旬,选择天气晴朗的上午,采用LI-6400便携式CO2/H2O红外分析仪和6400-02B红/蓝光源叶室(LI-COR,Lincoln,USA)测定气体交换过程以及相应的环境因子。每株苗木选取冠层上部向阳方3~5片叶片进行活体测定。测定时设定叶室的温度25 ℃,相对湿度控制在(60±5)%,外界空气进入叶室的流速500 μmol·s-1,CO2摩尔分数设定为400 μmol·mol-1。测定时先采用饱和光强对叶进行充分光诱导,当光合速率稳定后开始测定,每片叶子测定5~6个重复。测定输出的参数包括单位面积最大净光合速率(Pn)、气孔导度(gs)、蒸腾速率(T)等。用于光合测定的叶片面积采用扫描图像处理获得,随后将叶片在65 ℃下烘干至恒质量(精度0.000 1 g),获取其比叶面积(SLA,cm2·g-1):
SLA=叶面积/叶干质量。
(1)
单位质量最大净光合速率Pmax(nmol·g-1·s-1)由下式获得:
Pmax=(Pn×SLA)/10。
(2)
1.5 元素测定
测定光合的同时摘取与光合测定叶片相同位置的鲜叶100 g,放于冷藏箱4 ℃保存后带回实验室分析。在实验室内,将样品置于烘箱中65 ℃烘至恒质量(精度0.000 1 g),然用将样品粉碎、过筛、装袋,用于元素含量测定。叶氮含量采用全自动凯氏定氮仪(Kjeltec 8400,Foss,Hillerød,Danmark)测定。
1.6 数据处理
应用SPSS 13.0统计软件对数据进行相关分析和方差分析,并用Ducan法进行多重比较,方差分析前对所有数据进行方差齐性检验和正态分布检验。采用线性回归中的全部回归法分析参数间的关系。所有作图均由SigmaPlot 10.0完成。
2 结果与分析
2.1 不同施肥处理降香黄檀幼苗的生长
施肥显著促进了降香黄檀幼苗的生长,苗高、地径、总生物量及各器官生物量均存在显著的差异(表2,P<0.01)。和对照相比,施肥显著促进了降香黄檀幼苗地径和苗高的生长(P<0.01),但地径和苗高生长增大的幅度却因施肥量的不同而表现出不同的变化趋势(表2)。N3P3K1组合下的苗木具有最大的苗高和地径,分别为119.87、12.10 mm,而N1P1K1则为最小,苗高和地径分别为99.61、10.93 mm(表2)。
随着施肥量的增加,苗木的总生物量、茎生物量、叶生物量和根生物量均明显增大,并在施肥组合N2P3K4至N3P4K2间达到最大值,随后增大的幅度略有下降(表2)。茎生物量、叶生物量、根生物量和总生物量最大值分别为66.54 g(N3P3K1)、15.55 g(N2P3K4)、18.98 g(N2P3K4)和100.47 g(N3P3K1)。

表2 施肥对降香黄檀苗木生长和光合的影响
注:表中数值为平均值±标准差;同列数据后不同小写字母表示差异显著(P<0.01)。
随着总生物量的增大,各器官生物量均表现出增大的趋势,但各器官生物量在总生物量中的比例却有所不同(图1)。茎生物量在总生物量中所占比例和总生物量呈显著正相关关系,相关系数为0.60,而叶生物量和根生物量在总生物量中所占比例以及根冠比均和总生物量呈显著负相关关系,相关系数分别为0.59、0.60、0.61。
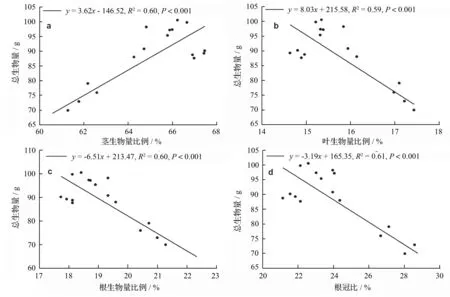
图1 施肥处理后降香黄檀幼苗生物量分配
2.2 不同施肥处理降香黄檀幼苗光合作用
施肥显著提高了降香黄檀幼苗的光合能力,并因施肥组合的不同而有所差异。随着肥量的增加,Pmax逐步增大,并在N3P3K1组合下达到最大值172.38 nmol·g-1·s-1,而后Pmax明显下降,并在N4P3K2组合时出现施肥光合最低值106.74 nmol·g-1·s-1(表2)。Pmax和叶片氮质量分数呈显著正相关关系(P<0.01,图2),相关系数为0.78。
光合能力和降香黄檀幼苗生长具有相关性,Pmax和叶生物量、总生物量间均为显著正相关关系(P<0.01,图3),相关系数为0.74和0.65。随着叶生物量的增加,总生物量也呈增大趋势,两者间相关系数为0.87。
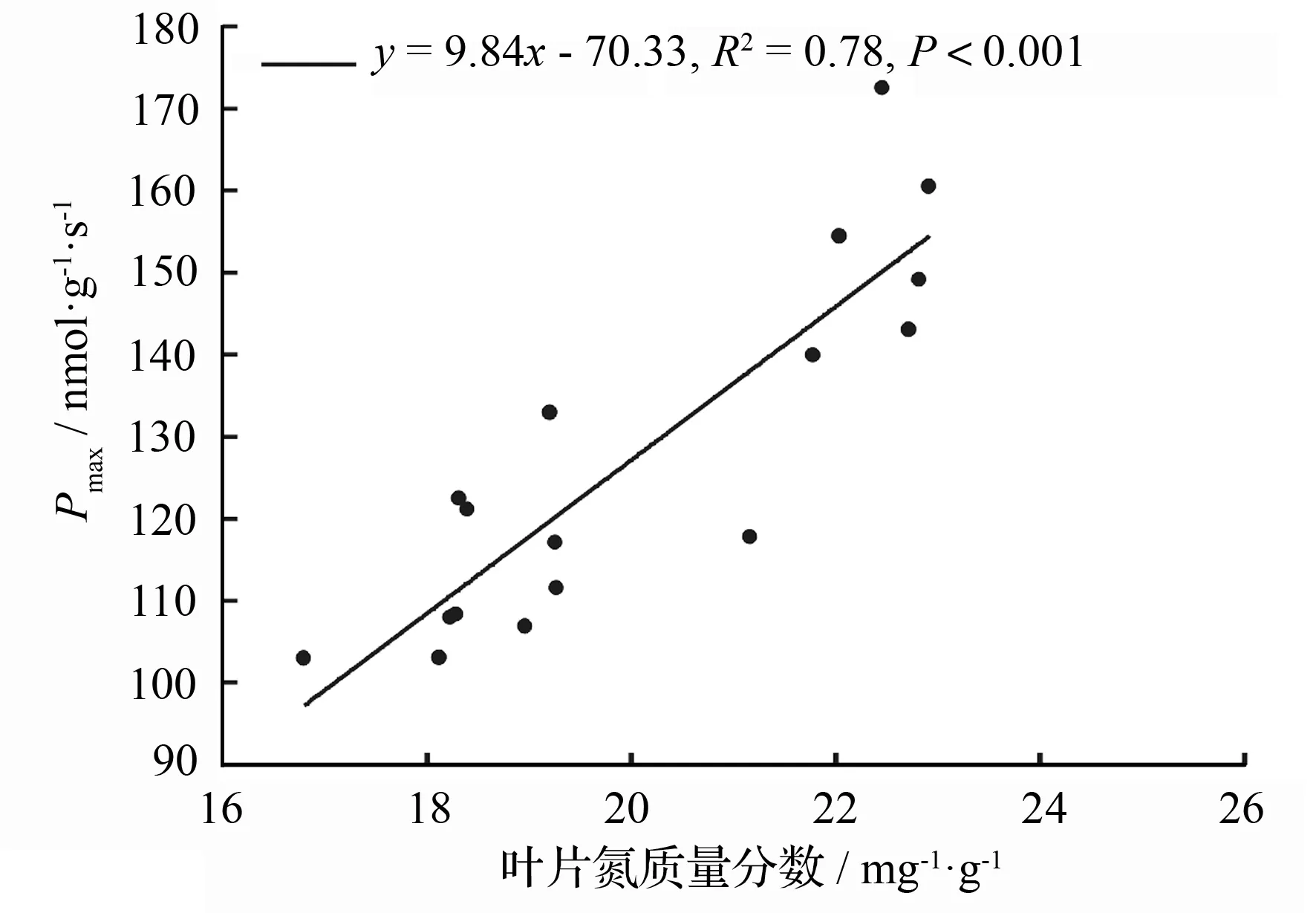
图2 降香黄檀幼苗最大净光合速率(Pmax)和叶片氮质量分数关系
3 结论与讨论
3.1 施肥对降香黄檀幼苗生长的影响
人工施肥可以改善苗木营养状况而促进其生长,氮、磷、钾元素作为苗木生长所需的大量元素对促进其生长尤为明显[12]。本研究表明,通过施氮、磷、钾肥有效地促进了降香黄檀的生长,其苗高、地径和总生物量均明显大于对照(表2),其增幅范围分别为7.59%~29.48%、2.25%~13.19%和18.18%~70.00%,相似的研究结果在对降香黄檀幼苗[13],以及其他针叶树苗[14]、阔叶树苗[15-17]的研究中得以体现。
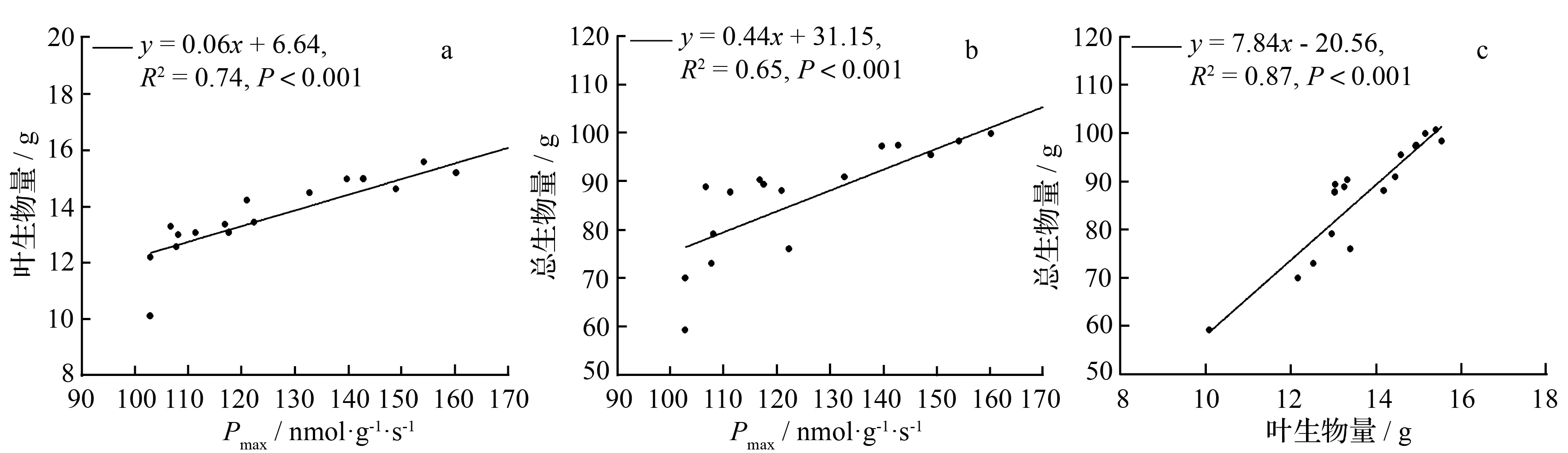
图3 降香黄檀幼苗最大净光合速率(Pmax)、总生物量和叶生物量间关系
养分元素对苗木生长的影响随着施加量和元素比例的不同,会表现出不同的反应,当养分元素超出一定量时,植物的生长会受到抑制。在本研究中,随着氮、磷、钾施加量的增加,降香黄檀的生长和生物量均表现出增大的趋势,但在配比N2P3K4至N3P4K2间苗木的生长较为稳定。这表明该区间的肥量已经满足苗木生长的需要,此时氮磷钾元素已不是限制植株生长的原因,而其他因素,诸如水分、温度、其他元素含量等环境因子成为限制其生长的主要因素[18]。在本研究中,N4P1K4配比后苗木生长表现出下降的趋势,这表明过多的元素含量(主要为氮元素)已对苗木的生长产生胁迫作用,导致其生长缓慢。有研究表明,当土壤中氮浓度过高时,苗木体内活性氧代谢失调,膜结构被破坏,渗透压发生改变,根系吸收功能受到影响,进而影响了植株的生长[18]。
施肥也会影响苗木生物量的分配格局。在本研究中,随着苗木总生物量的增大,茎生物量所占比例逐渐增大,由对照的60.58%增大到67.46%,增幅为1.17%~11.36%。叶生物量和根生物量所占比例以及根冠比随着苗木总生物量的增大均表现出减小的趋势,分别由对照的17.44%、22.34%和28.76%减小到14.64%、17.73%和21.56%。这是因为当土壤中养分元素缺乏时,苗木会加大根系生物量的分配比例以提高根系的养分吸收能力,近而茎叶所占的比例减小,而当土壤中养分元素供应充足时,苗木会将更多的生物量分配到茎的生长,因而叶和根的生物量比例减小[19-20]。根冠比能够较好地反映土壤养分供应水平对植物地上部和地下部生物量间关系的影响。本研究中,随着总生物量的增加,根冠比呈减小趋势,这也说明了施肥促进光合产物更多地向地上部分分配,降香黄檀幼苗在地上与地下部分的资源分配方面采取了一定的权衡策略。相似的结果在对其他树种的研究中也有所体现[21-24]。
3.2 施肥对降香黄檀幼苗光合作用的影响
在本研究中,施肥明显提高了降香黄檀的最大净光合速率,总体均值为129.15 nmol·g-1·s-1,比对照高出28.10%。这是因为施肥改善了土壤的养分利用状况,使叶片内的元素含量增加,进而光合作用增强。氮是氨基酸和核酸的必要组成成分之一,是光合物质代谢和植物生长的关键性因子,氮缺乏或过量都会导致叶绿素含量、酶含量和酶活性的下降,并进一步导致光合同化物的减少。在一定的范围内氮在叶中的含量和利用效率与叶的光合能力呈正相关[25-26]。本研究中,随着叶氮质量分数的增大Pmax呈明显增大趋势,这说明施肥后叶片氮质量分数的增加,是导致光合能力提高的原因。然而,当施肥量超出苗木的承受能力时,过多的养分含量会抑制根系的吸收和运输功能,影响光合酶的活性,进而导致光合能力的下降。
苗木生物量是光合产物累积的结果,光合能力的提高和叶量的增大均会导致光合产物的增加,因而影响生物量的累积和分配。本研究表明,Pmax、叶生物量和总生物量三者间均为正相关关系。这说明施肥后总生物量的增大是光合能力提高和叶生物量增大共同作用的结果。本研究也表明,施肥有助于光合能力和生物量的增大,但应注意施加的量和比例,对于降香黄檀幼苗而言,在N2P3K4至N3P4K2间的肥量最有利于苗木的光合和生长,且不会造成肥料的浪费。这对今后降香黄檀的实际育苗工作具有一定的指导意义。此外,本研究仅对光合速率进行了测定,而光合产物的累加是光合和呼吸共同作用的结果,因此今后研究还需加大对各器官呼吸的研究,这有助于深入探讨光合和生物量的关系。
[1] 邱治军,周光益,陈升华.海南特有珍贵红木树种:降香黄檀[J].林业实用技术,2004(6):41-42.
[2] 贾瑞丰.降香黄檀人工促进心材形成的研究[D].北京:中国林业科学研究院,2014.
[3] 吴国欣,王凌晖,俞建妹,等.降香黄檀幼苗年生长节律研究[J].浙江林业科技,2010,30(3):56-60.
[4] 倪臻,王凌晖,吴国欣,等.降香黄檀引种栽培技术研究概述[J].福建林业科技,2008,35(2):265-268.
[5] WRIGHT S J, YAVITT J B, WURZBURGER N, et al. Potassium, phosphorus or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest[J]. Ecology,2011,92(8):1616-1625.
[6] HERMANS C, HAMMOND J P, WHITE P J, et al. How do plants respond to nutrient shortage by biomass allocation?[J]. Trends in Plant Science,2006,11(12):610-617.
[7] SANTIAGO L S, WRIGHT S J, HARMS K E, et al. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition[J]. Journal of Ecology,2012,100(2):309-316.
[8] 朱声管,江海涛,蒋凡.1~3年生降香黄檀生长量研究[J].中国农业信息,2013(11):176-178.
[9] 吴国欣,王凌晖,梁惠萍,等.氮磷钾配比施肥对降香黄檀苗木生长及生理的影响[J].浙江农林大学学报,2012,29(2):296-300.
[10] WEGENER F, BEYSCHLAG W, WERNER C. High intraspecific ability to adjust both carbon uptake and allocation under light and nutrient reduction inHalimiumhalimifoliumL.[J]. Frontiers in Plant Science,2015,6:1-10.
[11] ZHAO D L, REDDY K R, KAKANI V G, et al. Nitrogen deficiency effects on plant growth, leaf photosynthesis, and hyperspectral reflectance properties of sorghum[J]. European Journal of Agronomy, 2005,22(4):391-403.
[12] 武维华.植物生理学[M].北京:科学出版社,2008.
[13] 吴朝辉.不同光照强度和施肥水平对降香黄檀容器苗质量的影响[D].南京:南京农业大学,2010.
[14] 易咏梅,谢忠华,章定清,等.日本落叶松矿柱材幼林施肥长期效应[J].华中农业大学学报,2002,21(3):231-234.
[15] 丁彦芬,高俊飞,张利.配方施肥对榉树幼苗生长的影响[J].南京林业大学学报(自然科学版),2014,38(S1):35-38.
[16] 郑辉,刘彦慈,李帅英,等.氮磷对刺槐生长和蛋白质组分的影响[J].河北农业大学学报,2006,29(5):44-46.
[17] CLOSE D C, BAIL I, HUNTER S, et al. Effects of exponential nutrient-loading on morphological and nitrogen characteristics and on after-planting performance ofEucalyptusglobulusseedlings[J]. Forest Ecology and Management,2005,205(1/3):397-403.
[18] KHAN F, KHAN S, FAHAD S, et al. Effect of different levels of nitrogen and phosphorus on the phenology and yield of maize varieties[J]. American Journal of Plant Sciences,2014,5(17):2582-2590.
[19] MÜLLER I, SCHMID B, WEINER J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants[J]. Perspectives in Plant Ecology,Evolution and Systematics,2000,3(2):115-127.
[20] HENDRICKS J J, NADELHOFFER K J, ABER J D. Assessing the role of fine roots in carbon and nutrient cycling[J]. Trends in Ecology & Evolution,1993,8(5):174-178.
[21] 陈琳,曾杰,徐大平,等.氮素营养对西南桦幼苗生长及叶片养分状况的影响[J].林业科学,2010,46(5):35-40.
[22] 贾瑞丰,尹光天,杨锦昌,等.不同氮素水平对红厚壳幼苗生长及光合特性的影响[J].林业科学研究,2012,25(1):23-29.
[23] 王力朋,晏紫伊,李吉跃,等.指数施肥对楸树无性系生物量分配和根系形态的影响[J].生态学报,2012,32(23):7452-7462.
[24] 郝龙飞,王庆成,刘婷岩,等.指数施肥对斑叶稠李苗木生物量分配、光合作用及根系形态的影响[J].林业科学,2014,50(11):175-181.
[25] MEZIANE D, SHIPLEY B. Interacting determinants of specific leaf area in 22 herbaceous species: effects of irradiance and nutrient availability[J]. Plant, Cell and Environment,1999,22(5):447-459.
[26] GRASSI G, MAGNANI F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees[J]. Plant, Cell and Environment,2005,28(7):834-849.
Effect of Fertilization on Growth and Photosynthesis ofDalbergiaodoriferaSeedlings//
Wang Nan, Wang Hongxin, Li Xianglin, Yuan Yang, Ma Bingchen, Chen Bo, Zang Gongsheng
(Sanya University, Sanya 572000, P. R. China)//Journal of Northeast Forestry University,2017,45(1):25-29.
Dalbergiaodorifera; Fertilization; Biomass; Photosynthetic capacity
王楠,女,1982年1月生,三亚学院艺术学院园林系,讲师。E-mail:mysky20040216@126.com。
陈博,三亚学院海南黄花梨产业开发研究所,副教授。E-mail:mysky20040216@126.com。
2016年7月16日。
S792.28; Q945.11
1)海南省高等学校科学研究重点项目(Hnky2016ZD-13)。
责任编辑:程 红。
