四川牡丹生境群落的数量分类与排序1)
2017-01-17夏小梅王娟彭培好刘贤安
夏小梅 王娟 彭培好 刘贤安
(成都理工大学,成都,610059)
四川牡丹生境群落的数量分类与排序1)
夏小梅 王娟 彭培好 刘贤安
(成都理工大学,成都,610059)
根据四川牡丹野外样方调查数据,采用双向指示种分析(TWINSPAN)方法对四川牡丹生境群落进行数量分类,并采用典范对应分析(CCA)方法对四川牡丹分布区群落与环境因子的关系进行定量研究。结果表明:数量分类将四川牡丹生境群落24个样方划分为10个群丛;CCA样方和物种排序结果显示,四川牡丹生境群落分布格局是由坡度、海拔、土壤自然含水率等环境因子综合决定的。而四川牡丹生境群落类型在很大程度上由群落优势种的分布格局决定,揭示了四川牡丹群丛类型和物种分布格局随环境梯度变化的趋势。
四川牡丹;生境群落;数量分类;CCA
数量分类和排序是揭示植物与环境之间复杂生态关系的有效途径,已成为生态学研究的重要内容之一[1-4]。目前,TWINSPAN是植物群落分类中使用最为频繁的多元等级分类方法[5-9],CCA是植被梯度分析与环境解释中最为常用的方法之一[10-13]。
四川牡丹(Paeoniadecomposita)属国家Ⅱ级重点保护植物[14],主要分布于岷江上游、大渡河两岸的马尔康、金川、丹巴、康定等地[15]。目前,关于四川牡丹的研究主要集中在种子萌发[16-17]、种子生理生化特性[18]等方面,但对四川牡丹生长环境群落分类、排序的相关研究未见报道。文中采用TWINSPAN及CCA方法对四川牡丹生境群落进行了数量分类与排序,探讨了四川牡丹生长环境与群落类型的关系,以期为探索国家珍稀濒危植物四川牡丹的保护提供科学依据。
1 研究地概况
本研究区位于四川省阿坝藏族羌族自治州、甘孜藏族自治州,地理位置在30°20′~31°59′N,102°1′~103°16′E,海拔2 204~2 880 m。该地区年均降水量650 mm,年均蒸发量为2 000 mm,干湿季节分明,干旱时间长。区域土壤分布主要是山地燥红土、燥褐土,具有熟化度低,土层薄,石砾多,渗透性强,保水保肥性差,有机质含量低,贫氮、缺磷、富钾的特点[19-20]。
2 研究方法
取样方法及野外调查:四川牡丹分布较为集中,且生长环境复杂多样,文中根据分布情况,共做24个10 m×10 m的样方,测定并记录经纬度、海拔、坡度、坡位、土壤自然含水率、土壤酸碱度、植被盖度、人为干扰率等环境因子,同时记录样方内各物种种名、株数、高度和盖度。
数据处理:依据基本调查数据,计算各样方内植物种类的重要值(IV)[21]。为有效剔除偶见种,选定频度不小于10%的38个种,形成24×38的原始矩阵,以此作为数量分类的分析对象。
应用WinTWINS 2.3统计软件进行TWINSPAN群落分类[22]。采用CANOCO for Windows5软件进行CCA[23-25]分析,CCA分析的环境因子包括经、纬度,海拔,坡度,坡位,土壤自然含水率,土壤酸碱度,植被盖度,人为干扰率。
3 结果与分析
3.1 四川牡丹生境群落数量分类
以24个样方的物种重要值为基础,采用TWINSPAN方法,将四川牡丹生境群落划分为10个群丛(图1),分别论述如下。
小果小檗(Berberisamurensis)+四川牡丹+小叶蔷薇(Rosawillmottiae)-须芒草(Andropogonvirginicus)群丛(Ⅰ):包括样方18、19,位于小金县美沃乡妖精沟山脊的上坡位,海拔2850~2 880 m,坡度30°~37°,土壤为暗棕壤,群丛总盖度为65%~85%。灌木层以小果小檗为建群种,优势种为小叶蔷薇和四川牡丹,其盖度为35%左右;草本层主要有须芒草、钩柱唐松草(Thalictrumuncatum)、细裂叶莲蒿(Artemisiagmelinii)、淡黄香青(Anaphalisflavescens)、甘西鼠尾草(Salviaprzewalskii),其盖度为40%。
金花小檗(Berberiswilsonae)+四川牡丹-钩柱唐松草群丛(Ⅱ):包括样方2、3、4、6,位于马尔康县脚木足乡石口咀山脊的上坡位,海拔2 557~2 702 m,坡度20°~40°,土壤为黄棕壤,群丛总盖度在50%~70%。乔木层盖度在20%以下,主要有白桦(Betulaplatyphylla);灌木层以金花小檗为建群种,常见有四川牡丹、宽卵叶山蚂蟥(Desmodiumfallax)、对刺雀梅藤(Sageretiapycnophylla)、金露梅(Potentillafruticosa)、尖瓣瑞香(Daphneacutiloba);草本层常见有大火草(Anemonetomentosa)、西南唐松草(Thalictrumfargesii)、淡黄香青、杯柄铁线莲(Clematisconnata)等。
四川牡丹+散生栒子(Cotoneasterdivaricatus)+金露梅-大火草群丛(Ⅲ):仅有样方7,分布在马尔康县脚木足乡山脊的中坡位,立地条件同群丛Ⅱ,海拔2 552 m,坡度24°,群丛总盖度为65%。以四川牡丹为建群种,小叶蔷薇和小果小檗为优势种,常见种有蜡莲绣球(Hydrangeastrigosa)、须芒草、钩柱唐松草、细裂叶莲蒿、淡黄香青、甘西鼠尾草等。

D.分级水平;N.样方数量;1~24.样方号。
四川牡丹+匍匐栒子(Cotoneasteradpressus)+野花椒(Zanthoxylumbungeanum)-须芒草群丛(Ⅳ):包括样方8、9、17,分布在马尔康县脚木足乡和小金美沃乡,海拔2 434~2 751 m,坡度22°~42°,土壤为黄棕壤和暗棕壤,群丛总盖度在60%~70%。灌木层盖度为20%,主要有匍匐栒子、宽卵叶山蚂蝗、金花小檗、小叶蔷薇、金露梅。草本常见种有细裂叶莲蒿、千里光(Senecioscandens)、大火草、西南唐松草等。
四川牡丹+高山栎(Quercussemecarpifolia)+宽卵叶山蚂蝗-希陶蹄盖蕨(Athyriumdentigerum)群丛(Ⅴ):包括样方15、16,分布在金川县卡撒乡,海拔2 725~2 775 m,坡度20°~25°,土壤为棕壤,群丛总盖度在60%~65%。以四川牡丹为建群种,宽卵叶山蚂蝗、高山栎为优势种,常见乔木有川楝(Meliatoosendan),灌木有黄杨(Buxussinica),草本有匍匐风轮菜(Clinopodiumrepens)、狗脊(Woodwardiajaponica)、希陶蹄盖蕨等。
四川牡丹+川西锦鸡儿(Caraganaerinacea)+散生栒子-大火草群丛(Ⅵ):仅含样方1,位于马尔康县卓格基乡居民楼后山坡上,人为采集较为严重,海拔2 686 m,坡度较缓,为10°,土壤为黄棕壤,群丛总盖度90%。其中乔木层盖度为20%,主要有铁杉(Tsugachinensis)、黄果冷杉(Abiesernestii);灌木层盖度为35%,常见有川西锦鸡儿、散生栒子;草本层盖度50%,主要有杯柄铁线莲、扁竹兰(Irisconfusa)、大火草、草木樨(Melilotusofficinalis)、桔梗(Platycodongrandiflorus)、荩草(Arthraxonhispidus)等。
四川牡丹+宽卵叶山蚂蝗-大火草群丛(Ⅶ),包括样方5、10、11、12、13,分布在马尔康县脚木足乡和马尔康镇俄尔雅村,海拔2 557~2 662 m,坡度20°~34°,土壤为黄棕壤和棕壤,群丛总盖度在60%~70%。灌木层主要有宽卵叶山蚂蝗、野山楂(Crataeguscuneata)、匍匐栒子、南方六道木(Zabeliadielsii)、金花小檗;草本层主要有淡黄香青、荩草、狗脊、西南唐松草、杯柄铁线莲等。
小马鞍羊蹄甲(Bauhiniafaberi)+四川牡丹-求米草(Oplismenusundulatifolius)群丛(Ⅷ):仅有样方14,分布在马尔康县脚木足乡石口咀山脊的下坡位,海拔2 434 m,坡度42°,土壤为黄棕壤,群丛总盖度为70%。乔木层盖度为25%,主要有岷江柏木(Cupressuschengiana);灌木层盖度为30%,以小马鞍羊蹄甲为建群种,四川牡丹、匍匐栒子、宽卵叶山蚂蝗为优势种,草本层常见种有截叶铁扫帚(Lespedezacuneata)、大火草、西南唐松草、千里光、求米草等。
马桑(Coriarianepalensis)+小雀花(Campylotropispolyantha)+四川牡丹-求米草群丛(Ⅸ):仅有样方24,分布在康定县金汤乡,海拔2 414 m,坡度58°,土壤为山地褐土,立地干燥,环境条件恶劣,群丛总盖度在75%。以马桑和小雀花为建群种,四川牡丹为优势种,常见种小叶蔷薇、求米草、两头毛(Incarvilleaargute)、大火草、川续断(Dipsacusasperoides)等。
四川牡丹+橿子栎(Quercusbaronii)+豪猪刺(Berberisjulianae)-求米草+大火草群丛(Ⅹ):包括样方20、21、22、23,分布在理县薛城乡和康定县孔玉乡,海拔2 068~2 218 m,坡度36°~63°,土壤为黄棕壤,群丛总盖度在40%~80%。没有乔木,灌木层盖度为35%,主要有橿子栎、豪猪刺、胡颓子(Elaeagnuspungens);草本层盖度为30%,主要物种有大火草、求米草、琉璃草(Cynoglossumfurcatum)等。
3.2 CCA排序
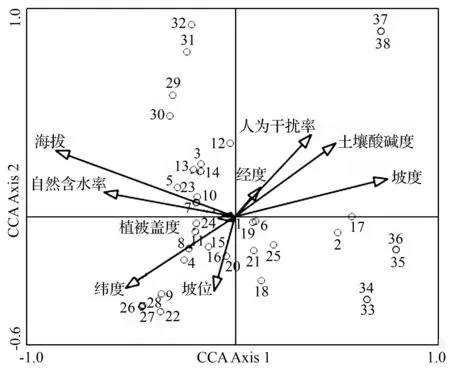
以24个样方、38个物种重要值形成的物种矩阵,以及经度、纬度、海拔、坡度、坡位、土壤自然含水率、土壤酸碱度、植被盖度、人为干扰率等9个环境因子为变量,进行CCA二维排序。CCA结果显示,第一轴和第二轴的特征值较高,能较好地反映四川牡丹生境群落与环境因子的关系,因此,选取前2个排序轴数据作出CCA二维排序图(图2)。CCA排序图中,带有箭头的线段表示不同的环境因子,线段的长短以及和排序轴的夹角表示该环境因子与排序轴的相关程度。
3.2.1 样方CCA排序
图2为24个样方的CCA排序图,结果显示,影响四川牡丹分布的主要环境因子是坡度、海拔、土壤自然含水率、人为干扰率、纬度、土壤酸碱度等。第一轴与坡度、土壤自然含水率、海拔相关,坡度在该轴内的作用更大,揭示了由坡度变化引起的土壤层厚度对四川牡丹分布格局的影响。沿着CCA排序第一轴从左至右,坡度逐渐升高,土壤自然含水率和海拔逐渐降低,群丛类型由四川牡丹+高山栎+宽卵叶山蚂蝗-希陶蹄盖蕨群丛(Ⅴ)过渡到四川牡丹+宽卵叶山蚂蝗-大火草群丛(Ⅶ),再到小马鞍羊蹄甲+四川牡丹-求米草群丛(Ⅷ),最后到四川牡丹+橿子栎+豪猪刺-大火草+求米草群丛(Ⅹ)。第二轴反映的群丛与环境的关系不明显,但对角线较为清晰地反映了四川牡丹生境群落与人为干扰率、土壤酸碱度、纬度等环境因子的变化情况,排序图从左下方到右上方,人为干扰逐渐增强,土壤pH值逐渐升高,纬度逐渐降低,群丛类型由四川牡丹+宽卵叶山蚂蝗-大火草群丛(VII)过渡到小马鞍羊蹄甲+四川牡丹-求米草群丛(VIII),最后到马桑+小雀花+四川牡丹-求米草群丛(IX)。

1~24.样方编号。
3.2.2 物种CCA排序
图3为四川牡丹生境群落38个物种的CCA排序图,由图3可知,四川牡丹生境群落各物种的分布格局与四川牡丹群丛类型的分布有较好的相似性,揭示了四川牡丹生境群落中各物种的分布在很大程度上决定了四川牡丹群丛类型的分布。沿着CCA第一轴从左至右,由分布在坡度相对较低、生长在海拔较高环境中的高山栎(26)、希陶蹄盖蕨(22)、宽卵叶山蚂蝗(8)、川楝(27)、匍匐风轮菜(28)等,逐渐过渡到分布在坡度较高、生长在海拔较低环境中的橿子栎(35)、豪猪刺(36)、马桑(37)、小雀花(38)、胡颓子(33)等。以四川牡丹、高山栎、宽卵叶山蚂蝗为优势种的群丛Ⅴ分布在CCA排序图最左端,以四川牡丹、橿子栎、豪猪刺为优势种的群丛Ⅹ分布在排序图的最右端,与样方分布图基本一致。沿着CCA排序图对角线从左下到右上角,由分布在纬度较高,生长在人为干扰率及土壤pH值较低环境中的高山栎(26)、宽卵叶山蚂蝗(8)、川楝(27)、云南勾儿茶(Berchemiayunnanensis)(9)、胡颓子(33)、鼠李(34),逐渐过渡到分布在纬度较低,生长在人为干扰率及土壤pH值较高环境中的马桑(37)、小雀花(38)、天门冬(32)、小果小檗(31)、蜡莲绣球(29)等,揭示了人为干扰、土壤酸碱度、纬度等环境因子共同影响了四川牡丹生境群落的组成与分布格局。同时以四川牡丹、宽卵叶山蚂蝗为优势种的群丛VII分布在排序图的左下角,以马桑、小雀花为优势种的群丛IX分布在排序图的右上角,与样方分布图也较为吻合。由上述可知,四川牡丹生境群落中各物种的分布格局在很大程度上决定着四川牡丹群丛类型的分布格局。
1~38.物种编号。
4 结论与讨论
应用TWINSPAN分类法将四川牡丹生境群落划分为10个群丛,这些群丛类型基本上代表了川西地区四川牡丹生境群落的类型。目前,TWINSPAN分类法虽已被广泛运用,但在具体分类过程中,还应该充分考虑群落的生境特征和植物物种在群落中的分布,避免由于软件的机械性而导致的不恰当分类结果,这样最后的群落分类结果才能更加符合植被分类原则。
由CCA二维排序图可知,四川牡丹生境群落物种的分布在很大程度上决定了四川牡丹群丛类型的分布。影响四川牡丹生境群落及物种分布格局的主要环境因子是坡度、海拔、土壤自然含水率等,其中地形因素坡度作用最大。在坡度较缓地带,土壤水分保持性较好,利于喜湿的植物生长,如甘西鼠尾草、蜡莲绣球、高山栎、川楝、匍匐风轮菜等,同时物种组成相对较为丰富。而坡度较大的区域,由于重力作用及地表径流等因素,水分不断下渗,从而导致其表层土壤的水分条件比坡度小的生境土壤含水量少,因此,抗旱植物生长较好,同时物种组成也相对简单,如橿子栎、豪猪刺、马桑等。
CCA结果显示,人为干扰也是影响四川牡丹生境群落分布的主要因素。近年来,阿坝藏族羌族自治州及甘孜藏族自治州旅游业发展较快,相关旅游活动对四川牡丹生境群落的结构、物种组成及其动态等产生了影响。旅游、放牧、祭祀、采摘等人为活动已经严重破坏了四川牡丹生长环境,应以就地保护为主,禁止人为干扰和破坏,为四川牡丹的良好生长提供有利条件。
[1] 江洪.川西北甘南云冷杉林的DCA排序、环境解释和地理分布模型的研究[J].植物生态学报,1994,18(3):209-218.
[2] 江洪,黄建辉,陈灵芝,等.东灵山植物群落的排序,数量分类与环境解释[J].植物学报,1994,36(7):539-551.
[3] 张新时.西藏阿里植物群落的间接梯度分析、数量分类与环境解释[J].植物生态学与地植物学学报,1991,15(2):101-113.
[4] 张峰,张金屯.历山自然保护区猪尾沟森林群落植被格局及环境解释[J].生态学报,2003,23(3):421-427.
[5] GAUCH H G, WHITTAKER R H. Hierarchical classification of community data[J]. Journal of Ecology,1981,69(20):537-537.
[6] ZHANG J T. Succession analysis of plat communities in abandoned croplands in the eastern Loess Plateau of China[J]. Journal of Arid Environments,2005,63(2):458-474.
[7] De CACERRESM, WISER S K. Towards consistency in vegetation classification[J]. Journal of Vegetation Science,2012,23(20):387-393.
[8] ZHANG J T, XIANG C L, LI M. Effects of tourism and topography onvegetation diversity in the subalpine meadows of the Dongling mountains of Beijing, China[J]. Environmental Management,2012,49:403-411.
[9] 吴东丽,上官铁梁,张金屯,等.滹沱河流域湿地植被的数量分类和排序[J].西北植物学报,2005,25(4):648-654.
[10] ZHANG J T, RU W M, LI B. Relationships between vegetation and climate on the Loess Plateau in China[J]. Folia Geobotanica,2006,41(2):151-163.
[11] WANG Z R, YANG G J, YI S H, et al. Effects of environmental factors on the distribution of plant communities in a semiarid region of the Qinghai Tibet plateau[J]. Ecological Research,2012,27(4):667-675.
[12] ZUO X A, WANG S K, ZHAO X Y, et al. Scale dependence of plant species richness and vegetation environment relationship along agradient of dune stabilization in Horqin sandy land, Northern China[J]. Journal of Arid Land,2014,6(3):334-342.
[13] 吴昊,张明霞,王得祥.秦岭南坡油松-锐齿槲栎混交林群落不同层次多样性特征及环境解释[J].西北植物学报,2013,33(10):2086-2094.
[14] 傅立国,金鉴明.中国植物红皮书:珍稀濒危植物(第1册)[M].北京:科学出版社,1991:530-537.
[15] 洪德元,潘开玉.芍药属牡丹组的分类历史和分类处理[J].植物分类学报,1999,37(4):351-368.
[16] 景新明,郑光华,裴颜龙,等.野生紫斑牡丹和四川牡丹种子萌发特性及与其致濒的关系[J].生物多样性,1995,3(2):84-87.
[17] SONG H X, LIU G L, CHEN Q B. Effects of aqueous extracts ofPaeoniadecompositaseeds on germination and some metabolic activities associated with growth of wheat seedlings[J]. Acta Biologica Hungarica,2012,63(3):362-71.
[18] 汪源,刘光立,张倩,等.层积处理对四川牡丹种子生理生化特性的影响[J].北方园艺,2013(24):59-62.
[19] 谢学强.干旱河谷不同立地条件对引种湿热型植物的影响[J].贵州农业科学,2011,39(12):202-204.
[20] 谢学强,肖晓凤,秦丹,等.大渡河干旱河谷野生半夏及其驯化栽培技术[J].现代园艺,2014(19):25-26.
[21] 方精云,王襄平,沈泽昊,等.植物群落清查的主要内容、方法和技术规范[J].生物多样性,2009,17(6):533-548.
[22] 王合玲,张辉国,吕光辉.艾比湖湿地植物群落的数量分类和排序[J].干旱区资源与环境,2013,27(3):177-181.
[23] 艾尼瓦尔·吐米尔,阿不都拉·阿巴斯.乌鲁木齐南部山区岩面生地衣群落特征的研究[J].菌物学报,2009,28(2):178-188.
[24] 艾尼瓦尔·吐米尔,张婷,马衣拉·莫合买提,等.新疆南山小渠子林场岩面生叶状地衣分布格局的研究[J].干旱区资源与环境,2013,27(7):142-147.
[25] JOHN E, DALE M R T. Environmental correlates of species distributions in a saxicolous lichen community[J]. Journal of Vegetation Science,1990,1(3):385-392.
Quantitative Classification and Ordination ofPaeoniadecompositaHabitat Communities//
Xia Xiaomei, Wang Juan, Peng Peihao, Liu Xian’an
(Chengdu University of Technology, Chengdu 610059, P. R. China)//Journal of Northeast Forestry University,2017,45(1):37-40.
With the data fromPaeoniadecompositafiled survey, we studied quantitative classification ofP.decompositahabitat communities by TWINSPAN, and analyzed the relationship betweenP.decompositahabitat communities and environmental factors by CCA. The 24 plots ofP.decompositahabitat communities were classified into 10 types of plant association by quantitative classification. The result of CCA plots and species ordination showed the relationship between the distribution pattern ofP.decompositahabitat communities and slope, altitude, and soil moisture. The types ofP.decompositahabitat communities were determined by the distribution pattern of dominant species, showing the variation trend ofP.decompositahabitat community types and species distribution with environmental gradient.
Paeoniadecomposita; Habitat community; TWINSPAN; CCA
夏小梅,女,1991年7月生,成都理工大学旅游与城乡规划学院,硕士研究生,E-mail:905878484@qq.com。
彭培好,成都理工大学旅游与城乡规划学院,教授。E-mail:peihaop@163.com。
2016年3月15日。
S685.11;Q948.15
1)四川省第二次全国重点保护野生植物资源调查项目(80303-KZZ031)。
责任编辑:任 俐。
