胰腺癌细胞总RNA转染树突细胞与胰腺癌-树突融合细胞激发特异性细胞毒T淋巴细胞能力的比较研究
2017-01-17李宏宇许文达郭晓钟
陈 江 李宏宇 王 迪 许文达 郭晓钟
胰腺癌细胞总RNA转染树突细胞与胰腺癌-树突融合细胞激发特异性细胞毒T淋巴细胞能力的比较研究
陈 江 李宏宇 王 迪 许文达 郭晓钟
目的比较人胰腺癌MiaPaCa-2细胞总RNA电转染树突细胞(Dendritic Cell,DC)与DCM iaPaCa-2融合细胞体外激发抗原特异性细胞毒T淋巴细胞(Cytotoxic T Lymphocyte,CTL)能力的差异。方法自6例胰腺癌患者外周血单核细胞中分离、培养DC。使用电穿孔法将M iaPaCa-2细胞总RNA转染DC,使用细胞融合方法将胰腺癌M iaPaCa-2细胞抗原负载DC,以未负载抗原的DC为对照。使用流式细胞术(FCM)检测PE-MUC/FITC-CD86抗体双标细胞评估融合效率;四甲基偶氮唑盐(MTT)检测转染各组DC存活率;混合细胞培养法评价各组DC体外刺激自体T淋巴细胞增殖能力;ELISA法检测各组DC体外激发抗原特异性CTL因子释放量。结果采用PEG-DMSO诱导的DC与M iaPaCa-2的融合细胞同时表达DC表型和MUC1分子,CD86与MUC1双阳性表达率为(42.3±7.30)%;融合细胞组DC存活率呈时间依赖性下降,转染后96h的存活率降低至62.81%,而M iaPaCa-2总RNA转染组DC细胞存活率稳定在85%左右,两组间差异有统计学意义(P<0.05);转染M iaPaCa-2总RNA DC刺激自体T细胞增殖指数(DC∶T=1∶10)为8432±611.25,显著高于DC-MiaPaCa-2融合细胞(DC∶T=1∶10)5672±107.51(P<0.05);且MiaPaCa-2总RNA转染DC激发特异性CTL分泌IL-12p70、IL-10和IFN-γ细胞水平亦显著异于DC-M iaPaCa-2融合细胞(P<0.05)。结论胰腺癌细胞总RNA转染DC较胰腺癌-树突融合细胞有更强的体外抗原特异性CTL激发能力。
树突细胞;RNA转染;细胞融合;胰腺肿瘤;细胞毒性T淋巴细胞
胰腺癌是一种常见的消化道恶性肿瘤,多数病例在发现时已属晚期,手术治疗及放、化疗作用有限,患者预后极差,临床迫切需要探索新的有效治疗手段[1]。树突细胞(Dendritic cell,DC)是体内功能最为强大的抗原递呈细胞(antigen presenting cell, APS),能够刺激并致敏T淋巴细胞,产生细胞毒性T淋巴细胞(cytotoxic T lynphocytes,CTLs),是机体免疫应答的核心。近年来出现的以DC为基础构建的肿瘤疫苗技术对多种肿瘤均有显著的治疗作用,为胰腺癌的临床治疗开辟了新的思路[2-4]。肿瘤细胞总RNA转染DC疫苗和DC-肿瘤细胞融合疫苗是当前DC肿瘤疫苗构建免疫治疗的两个热点方向[5]。本研究通过比较人胰腺癌M iaPaCa-2细胞总RNA电转染DC和DC-M iaPaCa-2融合细胞体外激发抗原特异性CTL能力的差异,为临床选择更为高效的DC肿瘤疫苗构建策略提供前期实验依据。
材料与方
一、一般材料及试剂
筛选2014年6月至2016年1月期间沈阳军区总医院消化科收治的6例HLA-A2+晚期胰腺癌患者,男性4例,女性2例,年龄35~65岁,平均年龄50岁。全部患者均经组织病理学检查(剖腹探查活检4例,细针穿刺活检2例)证实为胰腺癌。入选前未经放、化疗及免疫治疗。所有患者均签署知情同意书,并经本院伦理委员会批准。
人胰腺癌细胞株M iaPaCa-2(沈阳军区总医院消化科实验室);RPM I 1640、小牛血清(Hyclone公司,美国),rhGM-CSF、rhIL4、TNFα(PeproTec公司,美国),鼠抗人CD80、HLA-DR、CD83和CD86单抗(Santa-Cruz公司,美国),兔抗人MUC1多克隆抗体(DPC-Biermann公司,德国),TRIzol、MTT、50%PEG-10%DMSO溶液、Ficoll淋巴细胞分离液(Sigma公司,美国),3H标记甲基胸腺嘧啶(中科院原子能所),IL-12p70、IL-10、TNF-α和IFN-γ细胞因子检测试剂盒(BD公司,美国)。
二、DC的分离、培养和鉴定
参照文献[6]方法,通过Ficoll密度梯度离心法,分离来自胰腺癌患者100m L外周血中单核细胞(PBMCs),贴壁1 h后,取出未贴壁的细胞另作培养备用。贴壁细胞继续培养20 h后,更换新鲜培养液,加入rhGM-CSF(800 IU/m L)和rhIL-4(500 IU/ m L)。37℃,5%CO2,培养5 d后加入TNF-α(10 ng/ m L),培养至7 d,收集成熟DCs。使用倒置显微镜和电镜连续观察体外培养5~10 d DCs形态学变化。
三、胰腺癌M iaPaCa-2细胞总RNA提取、鉴定和转染
收获M iaPaCa-2细胞,加入Trizol试剂1m L充分裂解,5m in后加入氯仿200μL,4℃12 000 r/ m in离心15m in,加入异丙醇500μL,室温放置20 m in,4℃12 000 r/m in离心15m in,弃上清,加入75%乙醇1m L漩涡器震荡混均,4℃12 000 r/m in离心15 m in。弃上清,超净台晾干后溶于25μL DEPC水。分光光度计测定A260/A280值,计算RNA纯度和含量,1%变性凝胶电泳测RNA完整性后-80℃保存。
[7]的方法,取200μL培养5 d的未成熟DCs细胞悬液加入2mm的电极杯,加入20μg M iaPaCa-2总RNA。调整电压300 V、间隔125ms、4个脉冲,持续250ms进行电转染。电穿孔后立刻置入4℃冰箱静置10min,移入加有完全培养基的12孔培养板中,37℃、5%CO2培养0~96 h。以未转染RNA的mock-DC作为对照组。
四、细胞融合
选取连续传代培养20代后仍能稳定表达MUC1蛋白[8]的M iaPaCa-2细胞,使用25μg/m L丝裂霉素处理30m in,去增殖后与培养5 d的DC以3∶1混合,离心尽量吸尽上清,置37℃水浴中,滴加预热的50%PEG-10%DMSO溶液0.8m L,作用2 m in后加入1m L血清终止反应,未排除细胞间粘附及DC吞噬M iaPaCa-2细胞对实验结果的影响,以不加融合剂的mock-DC为对照。收集培养细胞,采用FITC-CD86及PE-MUC1双标记法,经流式细胞仪检测细胞融合率,融合率(%)=(融合组双标率-共培养组双标率)×100%。同时,使用MTT法检测各组DC存活率变化。
五、DC标志物检测
使用流式细胞法,分别检测转染M iaPaCa-2细胞总RNA及与M iaPaCa-2细胞融合后48 h的DC特异性标志物CD80、DC特征性成熟标志CD83、共刺激分子CD86以及MHC类分子HLA-DR等的表达水平,比较两者差异。
六、不同RNA转染组DC体外刺激自体T淋巴细胞增殖实验
分别收集不同转染组DCs,调节细胞浓度为1 ×105/m L作为刺激细胞,分选并调节患者自体PBMCs中的悬浮T淋巴细胞,浓度为1×106/m L作为反应细胞,按照刺激与效应细胞l∶10,l∶20,l∶40和l∶80比例加入到96孔板中,终体积200μL,在37℃,5%CO2培养5 d,收获细胞前18 h加入3HTdR,每孔1μCi。以RPM I-1640培养基替代效应细胞的4个孔作为对照组,使用液闪烁仪检测每分钟脉冲数(CPM)。
七、不同RNA转染组DC体外激发抗原特异性CTL释放细胞因子
以不同转染组DC为刺激细胞,以患者自体T淋巴细胞为效应细胞,刺激与效应细胞按l∶10比例混合反应14 d后,应用ELISA法,按试剂盒操作说明书检测细胞培养上清中IL-12p70、IL-10、TNF-α和IFN-γ水平。细胞培养上清用试剂盒中样本稀释液稀释50倍后检测。每个样本设3个复孔,取平均值。
八、统计学处理
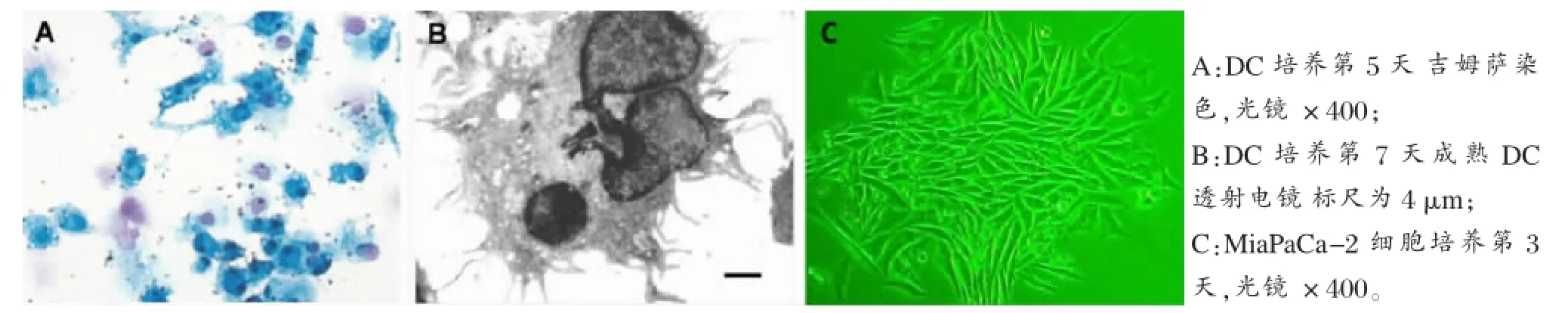
图1 体外培养DC及胰腺癌M iaPaCa-2细胞形态表现
结 果
一、DC与M iaPaCa-2细胞的体外培养和鉴定DC经体外分离、培养,数量可达初始数量的10~15倍。镜下可见胞体向四周伸出大量树枝状或裙褶状不规则突起,部分突起末端呈球状膨大(图1 A、B)M iaPaCa-2细胞呈集落样生长,条索状,异型性明显(图1 C)。
二、融合细胞的鉴定与融合率的检测
倒置显微镜下可见,经50%PEG-10%DMSO溶液诱导融合后,部分DCs与M iaPaCa-2细胞先发生细胞膜融合,形成双核的巨大细胞,而后细胞核逐渐发生融合,形成DC与M iaPaCa-2细胞的杂交细胞(DC-M iaPaCa-2)。
M iaPaCa-2为MUC1阳性细胞,不表达CD86抗原;而CD86为成熟DC特异性表面标志物之一,DC为CD86阳性细胞,不表达或低表达MUC1抗原。采用FITC-CD86及PE-MUC1双标记法检测融合细胞,结果示融合细胞组CD86及MUC1双阳性的DC比率为(42.3±7.30)%(图2),而共培养组CD86及MUC1双阳性DC比率为(7.21±1.06)%,除去细胞粘附等因素作用,DC-M iaPaCa-2融合细胞形成率为(35.09±6.24)%。
三、DC存活率变化情况
M iaPaCa-2细胞总RNA转染后0~96 h,DC存活率变化较小,稳定在85%左右,而M iaPaCa-2细胞与DC融合后,融合细胞存活率呈现时间依赖性降低,96 h时,DC-M iaPaCa-2融合细胞存活率低至61.23%(P<0.05),见图3。
四、不同分组DC体外刺激自体T淋巴细胞增殖实验
以M iaPaCa-2总RNA转染DC、DC-M ia-PaCa-2融合细胞及mock-DC作为刺激细胞,各组刺激细胞与自体T淋巴细胞混合培养,结果显示在DC∶T=1∶10时,DC-M iaPaCa-2 RNA细胞刺激自体T细胞增殖指数为8432±611.25,显著高于DCM iaPaCa-2融合细胞的5672±107.51(P<0.05)及Mock-DC的257±32.16(P<0.05);且在DC∶T=1∶20时,DC-M iaPaCa-2 RNA组细胞刺激自体T细胞增殖指数为5852±153.03,亦高于DC-M iaPaCa-2融合细胞组的4276±130.01(P<0.05)及Mock-DC组的108±17.18(P<0.05);而当DC∶T=1∶40及1∶80时,DC-M iaPaCa-2 RNA组细胞与DC-M ia-PaCa-2融合细胞刺激自体T细胞的增殖指数无显著差异(P>0.05)(图4)。
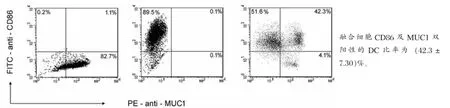
图2 与M iaPaCa-2细胞融合前后DC内CD86、MUC1双抗原相对表达情况
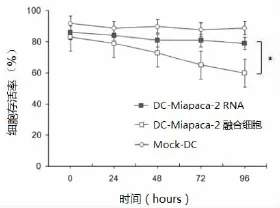
图3 不同分组DC存活率变化情况[转染后96 h,DCM iaPaCa-2细胞融合组细胞存活率显著低于M iaPaCa-2总RNA转染组DC(*P<0.05)]

图4 DC-MiaPaCa-2融合组,DC-MiaPaCa-2RNA转染组及M ock-DC组细胞刺激自体T细胞增殖实验
五、不同分组DC体外激发抗原特异性CTL释放细胞因子
M iaPaCa-2总RNA转染组DC、DC-M iaPaCa-2融合细胞及Mock-DC均具有刺激抗原特异性CTL分泌Th1型细胞因子的能力,但Mock-DC对CTLs的刺激作用较弱,几可忽略不计。ELISA法检测示,在DC∶T=1∶10时,M iaPaCa-2总RNA转染组DCs能激活自体特异性T细胞,其诱导特异性CTLs分泌IL-12p70和IFN-γ水平,显著高于DCM iaPaCa-2融合细胞(P<0.05);而M iaPaCa-2总RNA转染组DCs诱导特异性CTL分泌的IL-10水平,显著低于DC-M iaPaCa-2融合细胞;TNF-α水平两组间无显著变化(图5)。
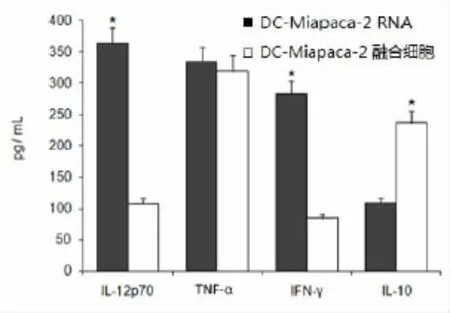
图5 DC∶T=1∶10时,DC-M iaPaCa-2RNA转染细胞诱导特异性CTL分泌IL-12p70和IFN-γ水平显著高于DCMiaPaCa-2融合细胞,IL-10水平则明显低于DC-M iaPaCa-2融合细胞(*P<0.05)
讨 论
DC肿瘤疫苗的实质是以特异性T细胞为基础的细胞免疫,而胰腺癌DC疫苗构建的关键在于选择合适的肿瘤抗原。胰腺癌缺乏特异性肿瘤抗原、异质性大、免疫原性差,使用单肿瘤抗原负载DC构建胰腺癌肿瘤疫苗时,其诱导的免疫反应通常较弱;而使用肿瘤全抗原负载DC构建胰腺癌疫苗则可激活针对不同肿瘤特异性抗原的多克隆细胞,减少肿瘤细胞克隆变异引起的抗原缺失,从而可以减少免疫逃逸的发生率,理论上是较具优势的DC肿瘤疫苗的构建策略[9]。
肿瘤细胞总RNA电转染DC疫苗与肿瘤-DC融合细胞疫苗,是当前临床研究中受关注较多的两种癌细胞全抗原负载DC疫苗制备方案。肿瘤-DC融合细胞是通过细胞融合技术将肿瘤细胞抗原导入DC,获得的杂交细胞既有DC的表面特征和功能,又能内源性表达肿瘤相关蛋白。负载有肿瘤细胞全抗原的DC能够体外激活T淋巴细胞多克隆性增殖,并促进相关CTLs分泌大量功能性细胞因子。而肿瘤细胞总RNA电转染DC,是通过RNA电转染技术,将含有肿瘤细胞全部抗原信息的总RNA导入DC胞内,在胞浆内对相关抗原基因进行转录,翻译,最终将肿瘤细胞抗原信息表达于DC表面,呈递并激活相关CTLs,实现特异性抗肿瘤免疫效应。
肿瘤-DC融合细胞疫苗的优势包括:①DC-肿瘤融合细胞能表达整个肿瘤细胞抗原决定簇,因而能诱导产生多克隆的CTL反应;②DC-肿瘤融合细胞既表达肿瘤抗原,又表达包括MHC、共刺激信号分子在内的DC表面抗原,使细胞介导的抗肿瘤免疫应答大大增强[10];③DC-肿瘤融合疫苗能够将已知和未知的肿瘤相关抗原加工后呈递给T淋巴细胞,发挥最佳的抗肿瘤免疫作用;④DC-肿瘤细胞融合后其抗原呈递作用持久[11],可诱导出更强的免疫应答反应。而肿瘤细胞总RNA转染DC疫苗的优势体现在:①转染的RNA只进入DC胞浆而非胞核,对细胞的损伤小;②RNA半衰期短,RNA转染DC疫苗更为安全;③负载的肿瘤抗原可从很少的癌细胞中经PCR扩增得来、RNA模板本身可经进一步的序列修饰,稳定性较强等[12]。RNA电转染法则更具有操作简单、安全、不受MHC遗传背景影响等优点[13]。
我们的研究发现,利用肿瘤细胞总RNA电转染技术,胰腺癌M iaPaCa-2细胞总RNA能够成功负载DCs,且转染过程中未出现抗原不良交互反应。而利用50%PEG-10%DMSO诱导DC的细胞融合技术,可使DC-M iaPaCa-2融合细胞的融合效率大于40%,凸显了该方法的有效性。胰腺癌M ia-PaCa-2细胞总RNA转染后0~96 h,DCs存活率稳定在85%左右,DC与胰腺癌M iaPaCa-2细胞融合后,细胞存活率在不同的时间点明显降低,提示此现象可能与50%PEG-10%DMSO融合剂对DCs的毒性蓄积和细胞膜的破坏作用有关。使用此两种方法负载胰腺癌M iaPaCa-2细胞总抗原后,DCs表面均可见细胞特异性CD80、HLA-DR、CD83和CD86分子的阳性表达,且表达水平无明显差异,说明两种方法对DC特异性共刺激分子及MHC表型分子的表达无显著影响。
本实验研究中,我们将DC-M iaPaCa-2融合细胞、DC-M iaPaCa-2 RNA转染细胞及Mock-DC与自体T细胞混合培养,结果显示在不同效靶比情况下,不同实验组DCs均能刺激自体T细胞增殖。但DC-M iaPaCa-2 RNA转染细胞对自体T细胞的增殖刺激能力显著高于DC-M iaPaCa-2融合细胞和Mock-DC,提示随着负载抗原效率的增加,DC刺激自体T细胞增殖的能力可显著增强。DC肿瘤疫苗的最终目的在于其诱导体内CTL反应,进而对肿瘤细胞产生特异性杀伤效应。随着T细胞的增殖和特异性CTLs的扩增,活化的CTLs可分泌大量Th1型细胞因子,如IL-12、TNF-α和IFN-γ等,同时减少Th2型细胞因子,如IL10等的释放,使得免疫反应本身向着有利于肿瘤细胞杀伤的Th1反应为主的方向发展。因此,DCs对体外CTL的刺激活化能力可间接使用IFN-γ等细胞因子的释放量表示[11]。本研究发现,DC-M iaPaCa-2融合细胞与M iaPaCa-2总RNA转染DCs均具有体外激发抗原特异性CTLs的能力,但DC-M iaPaCa-2 RNA可表现出更强的CTL激发效应。此发现与Van Tendeloo等[14]的研究结果类似。提示胰腺癌细胞总RNA转染DC法的抗原负载效率更强,其体外活化、激发抗原特异性CTLs的能力亦更强。
综上所述,本研究结果表明人胰腺癌细胞总RNA转染DC可表现出较胰腺癌-DC融合细胞更强的促进抗原特异性CTLs增殖和活化能力,此结果为临床选择更加有效的胰腺癌DC肿瘤疫苗构建方式奠定了前期实验基础。
参考文献
[1]Siegel R,Ma J,Zou Z,etal.Cancer statistics.CA Cancer JClin, 2014,64(1)∶9-29.
[2]Shore ND,Mantz CA,Dosoretz DE,etal.Building on sipuleucel-T for immunologic treatment of castration-resistant gastralological cancer.CancerControl,2013,20(1)∶7-16.
[3]Yokoyama S,Kitamoto S,HigashiM,etal.Diagnosisof pancreatic neoplasms using a novelmethod of DNA methylation analysis of mucin expression in pancreatic juice.PLoSone,2014,9(4)∶e93760.
[4]Tanaka T,Kitamura H,Inoue R,etal.Potential survival benefit of anti-apoptosis protein∶survivin-derived peptide vaccinewith and w ithout interferon alpha therapy for patients with advanced or recurrenturothelial cancer-results from phase Iclinical trials.Clin Dev Immunol,2013,2013∶262967.
[5]Chen J,Guo XZ,Li HY,et al.Comparison of cytotoxic T lymphocyte responses against pancreatic cancer induced by dendritic cells transfected w ith total tumor RNA and fusion hybrided with tumor cell.Exp BiolMed(Maywood),2015,240(10)∶1310-1318.
[6]Zeis M,Siegel S,Wagner A,et al.Generation of cytotoxic responses in mice and human individuals against hematological malignancies using survivin-RNA-transfected dendritic cells.J Immunol,2003,170(11)∶5391-5397.
[7]M ilazzo C,Reichardt VL,M uller MR.Induction ofmyelomaspecific cytotoxic T cells using dendritic cells transfected w ith tumor-derived RNA.Blood,2003,101(3)∶977-982.
[8]Chen J,LiHY,Wang D,et al.Human dendritic cells transfected w ith amplified MUC1 mRNA stimulate cytotoxic T lymphocyte responses against pancreatic cancer in vitro.J Gastroenterol Hepatol,2011,26(10)∶1509-1518.
[9]Zitvogel L,RegmanltA,Lcizer A,etal.Eradication ofwatablished murine tumorsusing a novel cell free vaccine∶dendritic cellderived exosomes.NatMed,2005,4(5)∶594.
[10]Trefzer U,Weingart G,Chen Y,et al.Hybrid cell vaccination for cancer immune therapy∶first clinical trial with metastatic melanoma.Int JCancer,2000,85(9)∶618-626.
[11]Koido S,Hara E,ToriiA,etal.Induction of antigen-specific CD4-and CD8-mediated T-cell responses by fusions of autologous dendritic cellsandmetastatic colorectal cancer cells.Int JCancer, 2005,117(4)∶587-595.
[12]Okano K,FukuiM,Suehiro Y,etal.Evaluation of anmRNA lipofection procedure for human dendritic cells and induction of cytotoxic T lymphocytes against enhanced green fluorescence protein.Tumour Biol,2003,24(6)∶317-324.
[13]Heiser A,Coleman D,Dannull J,etal.Autologous dendritic cells transfected with prostat specific antigen RNA stimulate CTL responsesagainstmetastatic prostatetumors.JClin Invest,2002,109 (3)∶409-417.
[14]Van Tendeloo VF,Ponsaerts P,Lardon F,et al.Highly efficient gene delivery by mRNA electroporation in human hematopoietic cells∶superiority to lipofection and passivepulsing ofmRNA and to electroporation of plasmid cDNA for tumor antigen loading of dendritic cells.Blood,2001,98(1)∶49-56.
Com parison of cytotoxic T lymphocyte induction by dendritic cells transfected with totalpancreatic can-cer RNA and fusion hybrided w ith tumor cell in vitro
CHEN Jiang,LIHong-Yu,WANG Di,XUWen-Da, GUO Xiao-Zhong.Departmentof Gastroenterology,Shenyang GeneralHospital of People Liberation Army, Shenyang 110016,China.Corresponding author:GUO Xiao-Zhong,Email:Guoxiaozhong1962@aliyun.com
ObjectiveTo compare theability of specific cytotoxic T lymphocytes(CTL)induction stimulated by dendritic cells(DC)transfectedw ith total RNA of human pancreatic cancer MiaPaCa-2 cell lines with DC fused with MiaPaCa-2 cell lines.MethodDCswere isolated and cultured from peripheral blood mononuclearcells(PBMCs).TotalRNA derived from MiaPaCa-2 cell lineswas transfected into DCsby electroporation.DCswere fused w ith M iaPaCa-2 cellsand the fusion efficiency wasassessed by flow cytometry (FCM).DC-MiaPaCa-2 hybridswere identified asPE-MUC1/FITC-CD86 double positive cells.The survival rate of DC wasdeterm ined by MTTmethod.The lymphocyte proliferation ability was evaluated bymixed cell culturemethod.The cytokines releasing of antigen-specific CTLsweremeasured by ELISA assay.ResultsDC-MiaPaCa-2 hybridswhich possessed both the phenotype of DC and MUC1 protein.The co-expression rate of both CD 86 and MUC1 positivity was(42.3±7.30)%.The survival rate of DCs transected w ith M ia-PaCa-2 total RNA were stabilized around 85%and the survival rate of DCs fused with MiaPaCa-2 cellswas 62.81%at96 hours(P<0.05).The autologous T cell proliferation index of the total RNA of M iaPaCa-2 celllines transfection DC group was8432±611.25,significantly higher than the DC-M iaPaCa-2 hybrids5672± 107.51(P<0.05);The secretion of IL-12p70,IL-10 and IFN-γlevels of the total RNA of MiaPaCa-2 cell lines transfection DC-specific CTL w ere significantly differentw ith DC-M iaPaCa-2 hybrids(P<0.05).ConclusionDCs transfected with the total RNA of pancreatic cancer cells had a stronger ability to stimulate specific CTL in vitro than the DCs fusedwith pancreatic cancer cells.
Dendritic cells;RNA transfection;Cell fusion;Pancreatic cancer;Cytotoxic T lym phocytes
2016-09-19)
(本文编辑:乔伟光)
10.3969/j.issn.1672-2159.2016.05.006
110016沈阳军区总医院消化科
郭晓钟,E-mail:Guoxiaozhong1962@aliyun.com
国家自然科学基金(81071982)
