乳酸菌在极端条件下抗逆性的研究进展
2017-01-14崔美岩张淼许宗为王佳蕊王栋张志霞陈骏谈重芳
崔美岩,张淼,许宗为,王佳蕊,王栋,张志霞,陈骏,谈重芳
(郑州大学河南省离子束生物工程重点实验室,郑州450000)
乳酸菌在极端条件下抗逆性的研究进展
崔美岩,张淼,许宗为,王佳蕊,王栋,张志霞,陈骏,谈重芳
(郑州大学河南省离子束生物工程重点实验室,郑州450000)
乳酸菌在自然界中的分布范围非常广泛,其中,有些特殊种类的乳酸菌在某些极端条件下仍然具有很好的生存能力。筛选具有较强耐盐性、pH值耐受性以及较宽温度生长范围的乳酸菌,能够大大拓宽其应用范围,其耐性机制也是微生物学家研究的热点。综述了乳酸菌在高质量分数盐、极低pH值、极高温、极低温等极端条件下,对乳酸菌的筛选与鉴定,耐性机理及应用。
耐盐性;pH值;温度;乳酸菌
0 引 言
极端环境指对生物生长产生限制因子的环境,如高温、低温、高酸、高碱、高盐等环境。乳酸菌能够发酵动植物产品形成特征性的发酵食品[1],耐盐性、耐酸性乳酸菌经常存在于酱类、泡菜等发酵食品中,对产品的风味、质地等特征具有非常重要的作用[2],耐盐性乳酸菌是腌渍过程的主要微生物类群[3]。检测益生菌的一个重要指标就是胃肠道环境的耐受性,胃液的pH值在3.0以下,对于微生物来说,这是一个极其严峻的环境,只有极少数耐酸的乳酸菌才能以活菌形式通过胃部。因此,开发出在强酸环境中也能较好存活的耐酸性乳酸菌和扩大乳酸菌的应用范围是息息相关的[4]。本文将从乳酸菌在极端温度、pH值、盐质量分数等恶劣生存条件下的筛选分离,所包含菌株种类以及耐受机理做一综述。
1 乳酸菌的耐盐性
乳酸菌能够影响食品的气味、色泽、质量甚至保藏时间等。由高盐浓度引起的高渗透压,触发细胞内水分外流,细胞胞质分离,从而引起细胞在结构和生理上的损伤,导致细胞停止生长,甚至死亡[5]。因此高浓度的食盐能够抑制致病性及一些腐败性微生物的生长,但是也会对乳酸菌的生长造成影响。筛选耐盐性比较强的乳酸菌并非易事,文献中所提到的乳酸菌对盐度的耐受性大多在20%以内。
耐盐性乳酸菌大多应用于食品中,比如酱油、农家大酱、腌制食品等。采用高盐稀醪发酵工艺生产酱油时需要在酱醅发酵过程中添加酵母菌和乳酸菌,但是酱油的NaC l质量分数高达18%,于是耐盐度达18%的乳酸菌就成为生产酱油的关键因素[6]。在大酱发酵过程中,高浓度的盐会抑制乳酸菌的生长,对最终大酱质量有很大影响,所以耐盐的乳酸菌对大酱品质的好坏起着决定性作用[7]。嗜盐乳酸菌是鱼酱发酵的最后阶段的一种优势菌株,这时鱼酱的颜色、风味、气味已经形成。据推测,这些嗜盐乳酸菌对鱼酱的风味及气味的形成起着重要作用。
1.1 常见耐盐性菌株
大多数乳酸菌的耐盐浓度都在10%以下,在0-6%的浓度范围内几乎所有乳酸菌都可以正常生长。耐盐度能达10%以上的乳酸菌基本都属于片球菌属和四联球菌属这二个属内。Natteewan Udomsil[8]等人从发酵鱼酱中分离出7株具有代表性的耐盐性的乳酸菌,这7株乳酸菌在25%浓度的培养基上经过七天的培养,菌落数达到10~100 mL-l,这些乳酸菌大都为四联或二联球菌,并且具有产挥发性化合物的能力。郭迪[6]从酱醅中分离筛选出一株耐盐性乳酸菌,在18%和20%的N aC l的质量分数下都能生长,并产生乳酸。对该菌进行分子生物学鉴定,证明筛选菌应属四联球菌属中嗜盐四联球菌中的一种。B.Kimura[9]等人从鱼酱中分离出能够产生组胺的Tetragenococcus muriaticus,一种四联球菌属嗜盐乳酸菌。Tetragenococcus muriaticus与T.halophilus有很近的亲缘关系[10],这种细菌能够在25%的盐质量分数下生长[11]。具有极强耐盐性的其他乳酸菌种属尚未见报道。
不同乳酸菌的耐盐能力存在差别(见表1),即使是同属不同种,甚至不同亚种的乳酸菌,其耐盐能力也有所不同[12]。
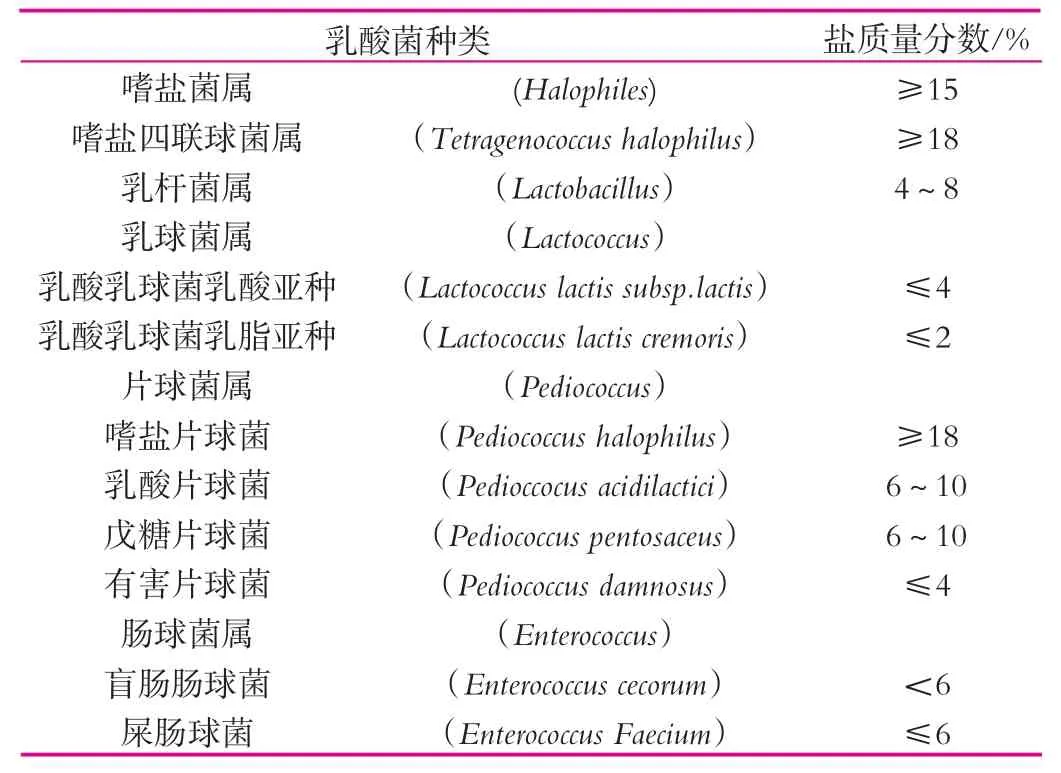
表1 部分乳酸菌的耐盐能力[13]
1.2 耐盐性乳酸菌的应用
耐盐性乳酸菌的应用是非常广泛的,(1)嗜盐微生物可以改善传统食品加工工艺[14];(2)高盐环境下嗜盐微生物及新型嗜盐酶发挥催化作用[8];(3)实现在高盐培养基中积累某些特定化合物,如四氢吡啶、甘油、组胺等[9];(4)嗜盐微生物应用于高盐环境中有机污染物降解或转化,生产可替代能源等方面[15-16]。(5)嗜盐微生物能产生独特化合物,而这些化合物是非高盐生长所必需的,如细菌视紫红质等,在制备细菌视紫红质的废料中可以提取不饱和脂肪酸,这扩大了嗜盐菌的应用范围[17]。
1.3 耐盐性机理
细胞能适应盐应激,是乳酸菌在自然环境及工业生产中存活、生长的重要因素。为了维持胞内的渗透压,乳酸菌利用许多策略来应对外界渗透压的变化。目前嗜盐微生物耐盐机制的研究主要集中在细胞吸K+排Na+作用、胞内积累小分子相容性溶质及嗜盐酶的氨基酸组成特性三个方面[14]。高效的N a+/H+逆向转运蛋白存在于一些嗜盐微生物中,在微生物耐受高盐环境胁迫的应答中起到了不可忽视的作用[18]。在嗜盐微生物中存在一种依赖累积胞内高浓度K+/C l-来维持渗透压平衡的机制[14],K+有利于恢复细胞的形态和渗透压,是酶活性和基因表达调节的细胞信使[19]。细菌中常见的K+吸收系统有:T rKA、K trA、Kup和Kdp[20-21]。当细胞受到盐胁迫时,通过外排N a+、吸收K+来恢复细胞内水和离子的平衡。积累相容性溶质来抵抗外界高盐环境是嗜盐菌主要的耐盐机制。在高渗条件下,由于分解代谢相容性溶质的酶被抑制,因此相容性溶质能在细胞内高浓度积累,执行渗透保护功能[12]。
2 耐酸碱性
2.1 耐酸碱性菌株的筛选
乳酸菌不但包括在食品发酵中使用的一般认为安全(GRAS)的微生物,而且还包括胃肠道中普遍存在的共生体和具有潜在益生作用的益生菌。对这些微生物来说,食品和胃肠道中的酸性环境对它们的生存是一个很大的挑战[22]。pH值的测定范围一般为2~10,多数研究表明大多数菌群的生长都会受到影响在pH值为2的情况下。针对乳酸菌耐酸碱性的实验有人工模拟胃液、耐胆盐、胰液实验等。J.Bautista-Gallego[23]等从食用橄榄中分离乳酸菌进行人工模拟胃液、胰液实验,发现乳酸菌在胰液中的生存率要比在胃液中的生存率高。这或许说明乳酸菌的耐酸性要比耐碱性普遍要弱。有关耐酸性乳酸菌的分离与筛选研究非常多,柏建玲[24]等对17株乳酸菌进行耐胃液实验,发现4株生长状况比较好的乳酸菌菌株,对其驯化,使菌株的胃液耐受性得到了不同程度的提高。
2.2 常见耐酸碱性菌株
嗜酸乳杆菌(Lactobacillus acidophilus)是人和动物肠道中的重要微生物,对人和动物具有益生作用,被广泛用于发酵食品与微生态制剂[25-27]。研究表明不是所有的嗜酸乳杆菌都能耐受低pH和高浓度胆盐环境,仅有少数嗜酸乳杆菌(L.aci-dophilus)具有这种能力[28]。耐酸性较好的菌株大多都为杆菌属,球菌数比较少见,但也有相关报道。如Lactobacillus coryniformis subsp.Torquens、Lb. bifermentans、Lb.amylophilus、Lb.helueticus和Lb.delbrueckii subsp.Bulgarcus[29],Lactobacillus plantarum、Lactococcus lactis[4]。云月英[30]等从酸菜汁中分离出6株耐酸性比较强的菌株,在pH值为3的情况下生长良好。采用传统的形态学、生理生化特性分析对这F株菌进行鉴定。鉴定结果:3株杆菌中植物乳杆菌、弯曲乳杆菌、干酪乳杆菌各1株;3株球菌中屎链球菌2株,鸟肠球菌1株。黄文强[31]等利用厌氧菌分离技术,从狼山鸡肠道中分离乳酸菌,通过形态、生理生化特性、16SrRNA基因序列测定以及同源性分析对其进行鉴定,并对菌株进行耐酸性、抗菌活性等筛选,耐酸性菌株为植物乳杆菌、卷曲乳杆菌、德氏乳杆菌以及嗜酸乳杆菌。段云浩[32]等从14株乳酸菌分离菌中筛选出2株具有低pH值耐受性的菌株,在pH 3.0的环境中能够正常生殖,经鉴定这两株菌为植物乳杆菌。研究表明,大多数杆菌属的耐酸性要比球菌属好。
2.3 耐酸性乳酸菌的应用
大部分乳酸菌在通过消化道到达肠道之前,因不能耐受胃液酸度而死亡,而少数菌能耐酸,此类耐酸性乳酸菌能在人体中更有效地发挥保健作用,因此具有很大的开发价值[4]。(1)降胆固醇、降血压和调节肠道微生态等多种保健作用;(2)运用于发酵食品业;(3)在畜牧业中的应用;(4)抑菌作用,可用作生物保护剂。
2.4 乳酸菌耐酸性机理
目前酸对微生物菌体的影响还未完全清楚,其耐酸性机制有很多种,大量的研究表明,H+-ATPase和谷氨酸脱羧酶(GAD)体系是乳酸菌维持胞内pH值稳定所使用的主要机制[22]。H+-ATPase在细菌中普遍存在,但还并不清楚它的分子结构和操纵子。该多聚体酶既能够利用质子合成ATP,又能利用ATP水解提供的能量将质子运送出细胞。在LAB中,后者活性在低pH值环境中会升高,这对于维持△pH是至关重要的[33]。
谷氨酸脱羧酶体系(GAD)是由Small和Waterman描述的一个简单的调控机制模型。它由一个谷氨酸脱羧酶、一个氨基酸通透酶和一个谷氨酸γ-氨基丁酸反向转运体组成。乳酸菌细胞通过特异的转运体摄入谷氨酸之后,在细胞内经过谷氨酸脱羧酶脱羧消耗一分子的胞内质子,反应的产物γ-氨基丁酸(GA-BA),通过反向转运体被排到胞外。最终结果是胞内氢离子的消耗导致细胞内的pH值升高,胞外的谷氨酸被转变成碱性略强的GABA而导致胞外pH值的略微升高[22]。
还有研究表明,耐酸性乳酸菌可以在强酸性条件下利用碳源(如葡萄糖)进行代谢的耐酸机理,而ATP酶活性不是影响耐酸性的主要因素[34]。此外,乳酸菌耐酸特性是多种机制共同作用的结果,如双组分信号转导系统,维持胞内外pH平衡,细胞膜组成变化,蛋白质、损伤修复等[35]。
3 极限温度耐受性
3.1 极高温耐受性乳酸菌的筛选
对于乳酸菌温度的耐受性的研究相对来说比较少,乳酸菌的最适温度一般在30~40℃,10℃及以下乳酸菌生长微弱甚至不生长[36]。乳酸菌温度测试范围大都在4~70℃之间。崔国艳等[37]从不同地方采集青贮玉米饲料、果渣、牛奶、新鲜的泡菜汁等样品中筛选耐高温乳酸菌,筛选出的六株乳酸菌在70℃条件下仍能生长。李检秀[38]等通过鼠李糖乳杆菌Lactobacillus rhamnosus JCM 1553进行紫外诱变,选育得到一株遗传性能稳定、耐47℃高温的L乳酸高产突变菌株,为工业化发酵生产L乳酸提供了新的备选菌种。
3.1.1 常见耐高温乳酸菌菌株
楚德强[39]从高温纤维乙醇发酵介质中分离到一株高耐受性乳酸菌,经16S rDNA分子生物学鉴定为乳酸片球菌(Pediococcus acidilactici)。另外,凝结芽孢杆菌属[40]、德氏乳杆菌[37]等都有耐高温特性相关报道。
3.2 极低温耐受性乳酸菌的筛选
来源不同、种类不同的乳酸菌温度生长范围及最适生长温度是不同的。来源于低温环境中的乳酸菌由于相对生长温度范围较低,被称之为低温乳酸菌[41]。其在生物保鲜方面的应用价值具有很大的潜力,也是现代研究乳酸菌的一个热点问题。邓梅[42]等从新疆冷水鱼肠道中分理出一株耐低温乳酸菌,其生长温度范围在4~37℃。
3.2.1 常见耐低温乳酸菌菌株
耐低温乳酸菌所属种类比较广泛,高阳[43]等从新疆阿勒泰地区额尔齐斯河流域冷水鱼肠道中分离耐低温的乳酸菌,24株菌其中有8株菌为嗜冷菌,最适生长温度在20℃以下,16为耐冷菌,通过生理生化和16SrDNA序列同源性分析显示24株耐低温乳酸菌隶属于6个属,其中肉杆菌属(Carnobacterium)3株,乳球菌属(Lactococcus)9株,肠球菌属(Enterococcus)7株,环丝菌属(Brochothrix)1株,魏斯氏菌属(W eissella)2株,链球菌属(Streptococcus)2株。李小艳[44]等于10℃条件从四川泡菜中分离获得乳酸菌36株,经过产酸能力初筛、低温测试复筛以及低温乳酸菌界定后,得到2株耐冷性强、温度生长范围为2~40℃的菌株,经鉴定菌株3m-1为植物乳杆菌(Lactobacillus plantarum),菌株8m-9为肠膜明串珠菌(Leuconostocmesenteroides)。
3.3 极限温度条件下乳酸菌的应用
工业中,利用高温乳酸菌可以缩短生产周期,节约冷却用水,减少污染的可能性。在众多固态发酵领域中一个难以解决的问题是固态发酵中迅速产生的高温对乳酸菌有杀灭作用,因而选育耐高温菌株具有极为重要的理论意义和广阔的应用前景[45]。耐低温乳酸菌常用于腌渍发酵青菜、低温青贮饲料、低温冷藏肉、泡菜中等,在青藏高原地区多在低温条件下进行青贮饲料,但是市售乳酸菌添加剂达不到低温储藏的目的,因此筛选低温条件下能够生存的乳酸菌对于青藏高原地区青贮饲料行业来说非常重要。
4 展 望
有关乳酸菌温度耐受性机理的研究还未见报道。乳酸菌在农业、医药、养殖业、食品业等方面都具有重要的作用,随着技术的发展,它的诸多用途将会被人们发现。其实普通的乳酸菌,活力极弱,它们只能在相对受限制的环境中存活,一旦脱离开这些环境,其自身也会遭到灭亡。筛选出一些强有力的、能够抵御各种不良环境的优良种质资源,才能发挥乳酸菌的生物学功效。在各种极端条件下,对乳酸菌生长状况进行研究是未来乳酸菌研究的一个热点。优良乳酸菌菌种资源的筛选将会极大地拓展乳酸菌的应用范围,并且提升其应用价值。
[1]施安辉,周波.乳酸菌分类、生理特征及在食品和酿造工业上的应用[J].中国调味品,2001(11):26-29.
[2]商君,钟方旭,王亚林,等.几种发酵蔬菜中乳酸菌的分离与筛选[J].食品科学,2007,28(4):195-198.
[3]曲玲童,牛文静,孙征,等.锦州腌渍小菜中耐盐乳酸菌的筛选与产酸性能研究[J].中国酿造,2012,31(6):83-87.
[4]杨郁,张丽靖,天知吾诚.耐酸性乳酸菌的筛选及应用的初步研究[J].浙江农业科学,2006(4):463-465.
[5]PRASAD J,MCJARROW P,GOPAL P.Heat and osmotic stree responses of prabiotic lactobacillus Mamnosus Hn001(DR 20)in ralaotion to viability after drying[J].Applied and Environmental Microbiology,2003,69(2):917-925.
[6]郭迪,杨汝德,潘力.酱油酿造中耐盐性乳酸菌的筛选及分子生物学鉴定[J].遵义师范学院学报,2008,10(1):52-54.
[7]AMARELA TERZIC-VIDOJEVIC,SANJA MIHAJLOVIC,GORDANA UZELAC,et al.Characterization of lactic acid bacteria isolated from artisanal Travnik young cheeses,sweet creams and sweet kajmaks over four seasons[J].Food Microbiology,2014,39:27-38.
[8]NATTEEWAN UDOMSIL,SUREELAK RODTONG,SOMBOON TANASUPAWAT,et al.Proteinase-producing halophilic lactic acid bacteria isolated from fish sauce fermentation and their ability to produce volatile compounds[J].International Journal of Food Microbiology, 2010,141(3):186-194.
[9]B KIMURA,Y KONAGAYA,T.FUJII.Histamine formation by Tetragenococcus muriaticus,a halophilic lactic acid bacterium isolated from fish sauce[J].International Journal of Food Microbiology,2008,10(1): 71-77.
[10]ROLING,W VAN VERSEVELD,H W.Characterization of Tetragenococcus halophila Populations in Indonesian Soy Mash(Kecap) Fermentation[J].applied and environmental microbiology,Apr.1996,62 (4):1203-1207.
[11]SATOMI M,KIMURA B,MIZOI,et al.Tetragenococcus muriaticus sp.nov,a new moderately halophilic lactic acid bacterium isolated from fermented squid liver sauce[J].Int.J.Syst.Bacteriol.1997,(47): 832–836.
[12]陈卫,赵山山,张秋香.乳酸菌的耐盐机制[J].中国食品科学,2013,13 (10):1-5.
[13]张刚.乳酸细菌基础-技术和应用[M].北京:化学工业出版社,2007.
[14]王伟伟,唐鸿志,许平.嗜盐菌耐盐机制相关基因的研究进展[J].微生物学通报,2015,42(3):550~558.
[15]OREN A.Industrial and environmental applications of halophilic Microorganisms[J].Environmental Technology,2010,31(8/9):825-834. [16]ZHUANG XL,HAN Z,BAI ZH,et al.Progress in decontamination by halophilic microorganisms in saline wastewater and soil[J].Environmental Pollution,2010,158(5):1119-1126.
[17]李冬冬.用嗜盐菌制备视紫红质和分离检测不饱和脂肪酸[D].北京化工大学,2007.
[18]VENTOSA A,N IETO JJ,O ren A.Biology of moderately halophilic aerobic bacteria[J].Microbiology and Molecular Biology Reviews,1998,62(2):504-544.
[19]M I CALDERN,C VARGAS,FROJO,et al.Complex regulation of the synthesis of the compatible solute ectoine in the halophilic bacterium Chromohalobacter salexigens DSM 3043T[J].Microbiology,2004, 150(9):30-51.
[20]M P ARENA,P RUSSO,V CAPOZZI,et al.Effect of abiotic stress conditions on expression of the Lactobacillus brevis IOEB 9809 tyrosine decarboxylase and agmatine deiminase genes[J].Annals of Microbiology,2011,61(1):179-183.
[21]L LEONTIEV,A.SELIVERSTOV,V.LYUBETSKY,et al.Algorithm for a large-scale search for t-box transcription regulation in bacteria[J].Molecular Biology,2005,39(6):946-948.
[22]张艳国,田雷.乳酸菌的耐酸性机制[J].饲料工业,2007,28(4):62-64.
[23]J BAUTISTA-GALLEGO,F N ARROYO-LÓPEZ,K RANTSIOU,et al.Screening of lactic acid bacteria isolated from fermented table olives with probiotic potential[J].Food Research International, 2013,50:135-142.
[24]柏建玲,吴清平,张菊梅,等.耐胃液乳酸菌的筛选、鉴定与驯化[J].中国食品安全高峰论坛论文集.
[25]FULLER R.Probiotics in human medicine[J].Gut,1991,32:439-442.
[26]COLLINS J K,Thornton G,Sullivan G O.Selection of probiotic strains for Human Applications[J].Int Dairy,1998(8):487-490.
[27]SANDERS M E,Klaenhammer T R.The scientific basis of Lactobacillus acidophilus NCFM functionality as a probiotic[J].J Dairy Sci,2001, 84:319-331.
[28]HOVEL H,NÙRGAARD H,BRÙBECH P.Lactic acid bacteria and the human gastrointestinal tract[J].European Journal of Clinical Nutrition,1999,53:339-350.
[29]白雅莉,金山,步仁巴雅尔,等.酸马奶中分离到的乳酸菌耐酸性及抗霉菌活性的研究[J].乳业科学与技术,2010,(2):51-55.
[30]云月英,王文龙,王雅娟.耐酸乳酸菌的筛选及初步鉴定[J].江苏农业科学,2013,41(1):256-259.
[31]黄文强,王永娟,张龙,等.狼山鸡消化道乳酸菌的分离、鉴定以及生物学特性研究[J].安徽农业科学,2012,40(25):12541-12543.
[32]段浩云,田洪涛,檀建新,等.葡萄酒mLF高耐受性优良乳酸菌的筛选与鉴定[J].中国食品学报,2015,15(5):200-205.
[33]VAN DE GUCHTE M,SERROR P,CHERVAUX C,et al.Stress responses in lactic acid bacteria,Antonie van Leeuwenhoek.International Journal of General and Molecular Microbiology,2002,14(82): 187-216.
[34]杨郁,张丽靖,天知吾诚.乳酸菌耐酸机理的研究[J].食品工程,2007(4): 42-45.
[35]胡爱华,熬晓琳,陈岑,等.乳酸菌耐酸耐胆盐机制的研究进展[J].食品工业科技,2015(8):380-389.
[36]熊素玉,姚新奎,谭小海,等.不同温度及pH条件对乳酸菌生长影响的研究[J].新疆农业科学,2006,43(6):533-538.
[37]崔国艳,陈五岭,张雄鹰,等.耐高温乳酸菌的分离与鉴定[J].长治医学院学报,2009,23(2):102-104.
[38]李检秀,孙靓,吴军华,等.一株耐高温L乳酸高产菌的选育及其发酵特征[J].广西科学,2011,18(3):269-272.
[39]楚德强,赵凯,吴倩,等.一株高耐受性乳酸菌的分离及其在木质纤维素发酵生产高浓度L-乳酸中的应用[J].华东理工大学学报,2011,37 (5):521-523.
[40]蒲春,李鸿梅,王治同,等.耐高温乳酸菌的筛选与鉴定[J].粮油加工, 2010,(11):167-171.
[41]HUGASM,Monfort JM.Bacterial starter cultures for meat fermentation,Food chemistry,1997,59(4):547-554.
[42]邓梅.新疆冷水鱼肠道耐低温乳酸菌多样性及产细菌素菌株的筛选[D].石河子大学,2011.
[43]高阳.新疆冷水鱼肠道耐低温乳酸菌分离鉴定及系统发育研究[J].微生物学报,2013,53(1):82-91.
[44]李小艳,刘书亮,蒲彪,等.泡菜中低温乳酸菌的筛选及其特性研究[J].中国食品学报,2014,14(10):08-116.
[45]赵玉红,张立钢,庞伟娜.乳酸菌和酵母菌共生发酵生产面包的研究[J].食品工业科技,2003,24(3):61-62.
Research progress o f Lacticacid bacteria resistance under extrem e conditions
CUIMei-yan,ZHANGM iao,XU Zong-wei,WANG Jia-rui,WANG Dong,ZHANG Zhi-xia, CHEN Jun,TAN Zhong-fang
(Zhengzhou U niversity Henan Key Laboratory o f Ion Beam Bioengineering,Zhengzhou 450000,China)
Distribution of Lacticacid bacteria isvery broad in nature,among them,some specialkindsof lactic acid bacteria stillhasgood viability in extreme conditions.Screening lactic acid bacteriaw ith a strong salt tolerance,pH tolerance and aw idely growth temperature range can greatly broaden its range of applications,its resistancemechanism isahot research form icrobiologist.Wemake a review about the screening and identification of lactic acid bacteria at high concentrationsof salt,low pH,very high temperature,very low temperature and other extreme conditions,Containing its tolerancemechanism and application.
salt tolerance;pH;temperature;Lacticacid bacteria
Q935
B
1001-2230(2016)05-0028-04
2015-10-20
国家自然科学基金(41101244);河南省基础与前沿研究(1523 00410037,132300413205);郑州大学大学生创新训练计划。
崔美岩(1990-),女,硕士,从事乳酸菌的分离鉴定研究。
谈重芳
