鸡爪槭翅果表型性状变异
2017-01-11黄东静冯刚刘继虎万志兵
黄东静,冯刚,刘继虎,万志兵
(黄山学院生命与环境科学学院,黄山 245041)
鸡爪槭翅果表型性状变异
黄东静,冯刚,刘继虎,万志兵
(黄山学院生命与环境科学学院,黄山 245041)
为了阐明鸡爪槭(Acer palmatum)翅果表型性状间的关系及其变异规律。从黄山学院校园内随机选择10棵生长健康的鸡爪槭单株,对10棵鸡爪槭的9个翅果表型性状进行调查分析。10棵鸡爪槭的9个翅果表型性状的遗传变异系数范围为5.68%~19.71%;9个翅果表型性状间相关系数的范围是-0.451 4~0.729 4,这表明翅果表型性状间关系密切复杂;基于9个翅果表型性状划分的4个组别间表型性状差异明显,可为营建不同用途的鸡爪槭提供基本材料。研究表明鸡爪槭翅果表型性状变异丰富,多层次的变异为不同选育目标奠定了基础。
鸡爪槭;翅果;表型性状;变异规律
表型指基因型在个体水平上的表达,是基因型和环境因素互作的结果;而表型性状是检测遗传变异最直接和最简便的方法[1]。目前对于植物表型性状的研究主要是从遗传上进行研究,侧重于寻找具有优良性状的遗传材料,为遗传改良奠定基础[2]。植物的表型性状包括种实、花、叶及树体性状等,其中种实性状是植物有性生殖系统的重要组成部分,是生存和繁衍适应性的集中表现[3]。相比较而言,种实变异具有较高的稳定性,是植物遗传变异的表征,也是遗传多样性研究的先导和基础,其测定和评价的结果对于林木遗传育种具有指导意义[4]。黄勇等[5]以小果油茶种实为研究对象,通过分析10个种实性状得出小果油茶种实变异程度高,其数量性状变异程度高于质量性状,这为小果油茶的选择育种提供了理论依据。王丹等[6]研究不同海拔的茶条槭,结果表明茶条槭翅果表型性状变异丰富,对其良种选育具有一定的指导意义。
鸡爪槭(Acer palmatum),又称鸡爪枫,槭树科槭树属落叶小乔木,原产于中国,朝鲜半岛和日本也有分布[7]。鸡爪槭树姿优美,叶形奇特,为世界著名观赏树种,也是槭树属植物中培育园艺品种最多、园林绿化应用最广泛的树种之一[8]。近年来,鸡爪槭在园林应用中越来越广泛,对其形态变异研究主要集中在叶色变化[9]方面,而对种实形态变异研究较为少见[10]。为此,试验以鸡爪槭翅果表型性状为研究对象,探讨鸡爪槭翅果表型性状的变异规律,以期为后续鸡爪槭优良单株、良种的选育和种质资源开发利用提供理论依据。
1 材料与方法
1.1 材料来源与采集地的概况
试验材料来自于安徽省黄山市黄山学院校园内,地理位置东经118°17′、北纬29°41′,海拔高度163 m,属于北亚热带湿润季风气候,年平均气温16.3℃,1和7月份平均温度分别为3.5和27.9℃,年降水量1 359~1 701 mm,全年无霜期229 d。土壤类型为黄红壤,由于南校区地势较为平坦,立地条件相差不大。
1.2 翅果性状及测定
2014年11月,随机选择10株生长健康、树龄相同、无性繁殖的鸡爪槭单株,将所选单株的翅果全部采集并将翅果保存在干燥阴凉的环境中,按照采集的顺序及其对应的单株将所采得的翅果进行编号。选择遗传相对稳定、易于获得和测定的表型性状,包括翅长(wing length,WL)、翅宽(wing width,WW)、翅形系数(WL/WW)、小坚果长(samara vertical diameter,SVD)、小坚果宽(samara horizontal diameter,SHD)、种形系数(SVD/SHD)、小坚果厚(samara side diameter,SSD)、着生痕长(a trace length,ATL)、小坚果重(samara weight,SW)共计9个表型性状。待翅果自然风干后,每个单株随机抽取50粒两翅完整的翅果。用电子游标卡尺对每个单株所抽取的翅果分5组测量,每组10粒共计50粒翅果,测量精度为0.01 mm。用1/1000电子天平称量每粒小坚果的质量,称量精度为0.001 g并将测量结果详细记录。
1.3 数据统计与分析
研究数据采用EXCEL2003软件进行整理和初步分析。采用SPSS19.0软件对10份材料的9个翅果表型性状进行系统分类、方差分析和性状相关性分析等,在进行聚类分析时需先对数据进行标准化处理。
2 结果与分析
2.1 鸡爪槭翅果表型性状的差异分析
10棵鸡爪槭的翅果表型性状的均值分析表明:翅宽、小坚果长、小坚果厚和小坚果重均值最大是1号单株,其中翅宽和小坚果厚均值最小是3号单株,小坚果宽和小坚果重均值最小分别是6号单株和9号单株;翅长、小坚果宽和着生痕长均值最大是2号单株,翅长和小坚果长均值最小是7号单株,着生痕长均值最小是6号单株;翅形系数均值最大是3号单株,最小为7号单株;种形系数均值最大是4号单株,最小是2号单株。其中1、2号单株相比较于其余单株翅果生长发育较好,而3、7号单株相比较而言较差。
常用变异系数(CV)比较不同性状间的变异程度,变异系数越大则性状值离散程度越大[11]。鸡爪槭翅果表型性状的变异系数见表1。从表1可以看出,鸡爪槭翅果各表型性状在不同单株间的变异程度不同,变异系数由大到小依次为:小坚果重(19.71%)>种形系数(14.58%)>翅长(9.86%)>小坚果厚(9.35%)>翅宽(9.28%)>小坚果长(9.07%)>翅形系数(7.58%)>小坚果宽(6.78%)>着生痕长(5.68%)。其中小坚果重的变异系数最大,而着生痕长变异系数最小,这说明鸡爪槭翅果不同表型性状的离散程度不同,稳定性也不同。由种形系数(14.58%)大于翅形系数(7.58%)说明小坚果的稳定性小于翅的稳定性;翅长(9.86%)大于翅宽(9.28%),小坚果长(9.07%)大于小坚果宽(6.78%)这表明翅果横向的长短变异大于纵向的长短变异。
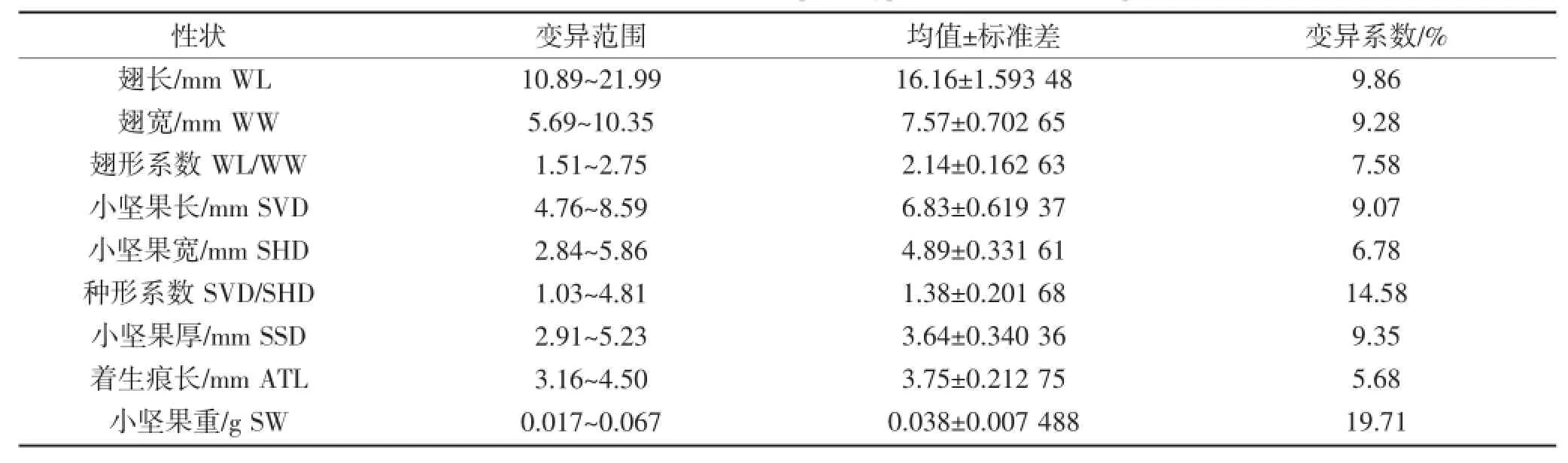
表1 鸡爪槭翅果表型性状变异系数Table 1 Variable coefficient in samara phenotypic variation of Acer palmatum
为了明确鸡爪槭翅果表型性状在单株间的差异,进行了方差分析,结果表明翅果的9个表型性状在不同单株间均达到了极显著的水平(P<0.01)。这表明翅果表型性状在不同单株间存在着极显著的差异,同时也说明在不同单株间存在着广泛的变异。
对10棵鸡爪槭翅果表型性状指标在α=0.05水平下进行多重比较,由分析结果可以看出:翅长、小坚果宽、着生痕长和小坚果重方面1、2号单株间没有显著性差异,而翅宽、翅形系数、小坚果长、种形系数、小坚果厚方面1、2号单株间有显著性差异;小坚果厚方面除1、2和3号单株间有显著性差异,其余单株间没有显著性差异;3、4号单株间在小坚果厚、着生痕长和小坚果重方面有显著性差异,在其余表型性状方面有显著性差异;5、6和7单株间在翅长、小坚果厚、着生痕长和小坚果重方面没有显著性差异;8、9、10号单株间在翅长、翅宽、翅形系数、小坚果宽、种形系数、小坚果厚、着生痕长和小坚果重方面没有显著性差异。
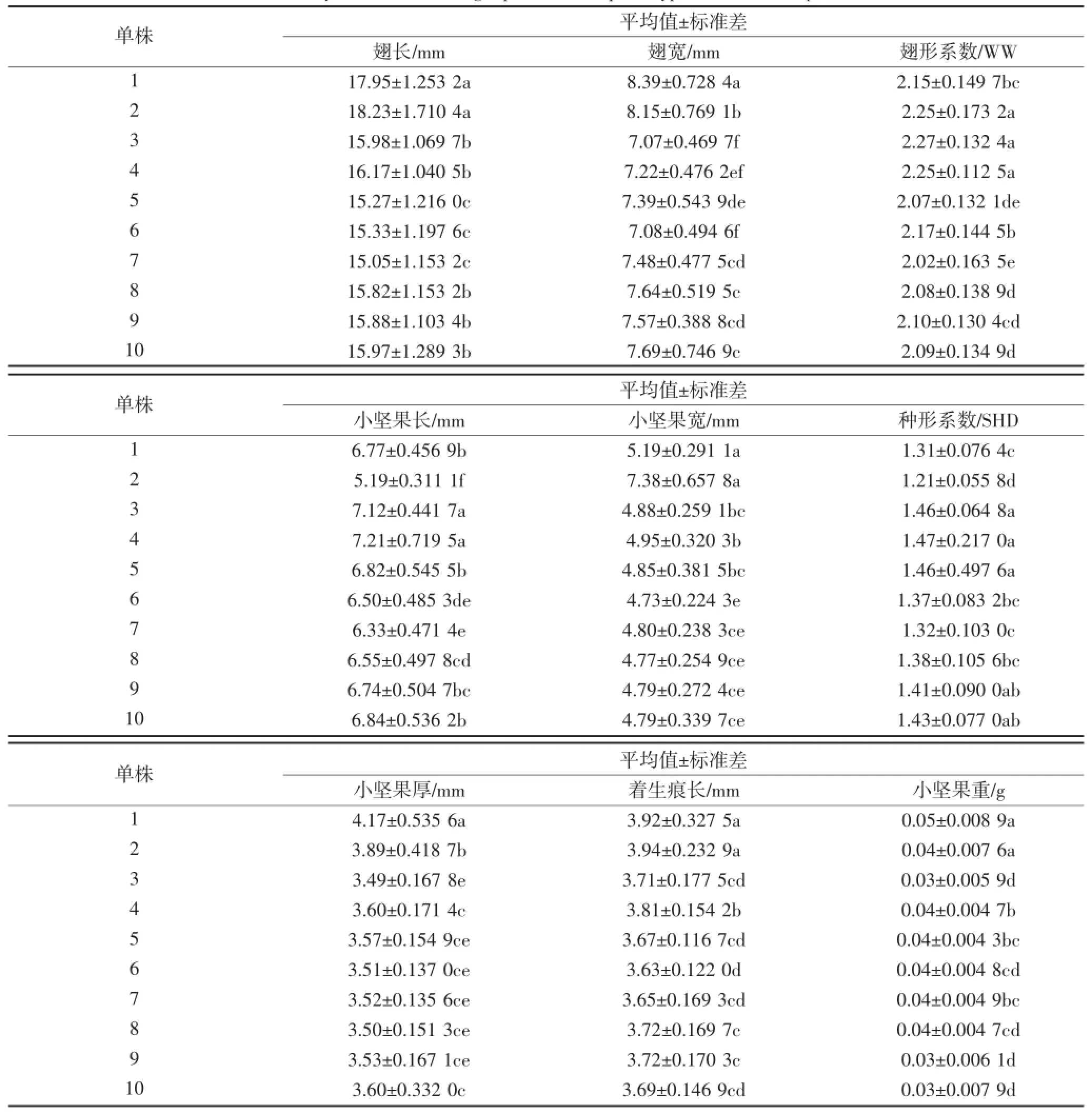
表2 不同单株鸡爪槭翅果表型性状差异分析Table 2 Analysis of different single plant samara phenotypic traits of Acer palmatum
2.2 鸡爪槭翅果表型性状间的相关性
鸡爪槭翅果表型性状间存在复杂的关系,使测得的数据存在重叠现象,因此为了更进一步分析翅果表型性状间的关系,对鸡爪槭翅果表型性状进行相关性分析,由表3可知鸡爪槭的9个翅果表型性状间相关关系除翅形系数与种形系数、小坚果厚和小坚果重间没有显著性相关,其余表型性状间具有极显著性相关或显著性相关;翅长与种形系数相关性呈负显著性相关(-0.102 1*),与其余翅果表型性状间相关关系均达极显著性水平;翅宽与其他表型性状间相关关系均具有极显著性相关,其中与翅形系数(-0.334**)和种形系数(-0.168 1**)呈负极显著性相关;翅形系数与翅长(0.447 3**)呈正极显著性相关而与翅宽(-0.333 4**)呈负极显著性相关,与种形系数、小坚果厚和小坚果重没有显著性相关;小坚果重与小坚果宽和小坚果厚的相关系数达到0.700 0以上,可见小坚果饱满程度越好重量越大且相关性越紧密;小坚果长、小坚果宽、小坚果厚和小坚果重两两之间的相关关系均达到极显著性相关,说明小坚果长、小坚果宽、小坚果厚和小坚果重间相关性紧密;着生痕长仅与种形系数(-0.113 7**)呈负极显著性相关;翅性状(翅长、翅宽)与小坚果性状(小坚果长、小坚果宽、小坚果厚和小坚果重)间存在正向相关,且均达到极显著性相关,说明翅性状和小坚果性状的遗传途径可能是相互关联的,使翅性状和小坚果性状的内在关联度高。
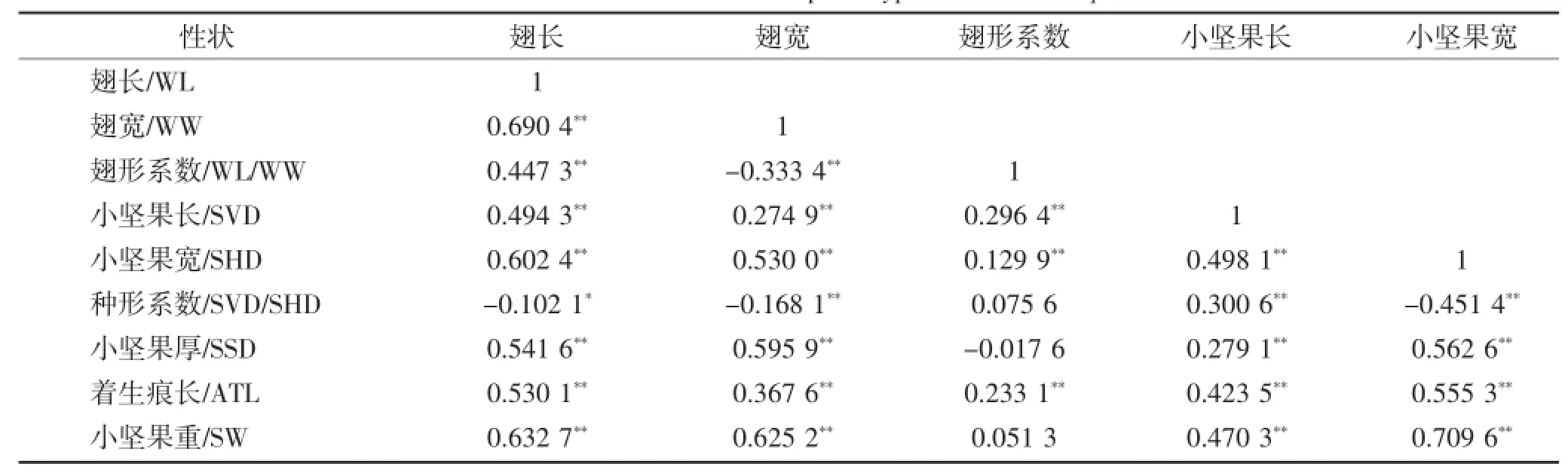
表3 鸡爪槭翅果表型性状间的相关性Table 3 Correlation between samara phenotypic traits of Acer palmatum

续表3 鸡爪槭翅果表型性状间的相关性Continued table 3 Correlation between samara phenotypic traits of Acer palmatum
2.3 鸡爪槭翅果表型性状聚类
由于鸡爪槭翅果9个表型性状指标存在不同的量纲和数量级单位,为了使不同量纲、不同数量级的数据能放在一起进行比较,先将10棵鸡爪槭的9个数量性状数据进行标准化处理,之后采用系统聚类法中的重心聚类法进行分析。图1反映了鸡爪槭个体间的亲疏关系、表型变异层次和各组单株的特征,以欧式平均距离5为阈值,将其分成4组,计算各组性状的平均值用于各组特征分析,结果见表4。

表4 鸡爪槭聚类分组中4组翅果表型性状均值的比较Table 4 Mean values of samara phenotypic traits of each group after clustering analysis of 10 Acer palmatum
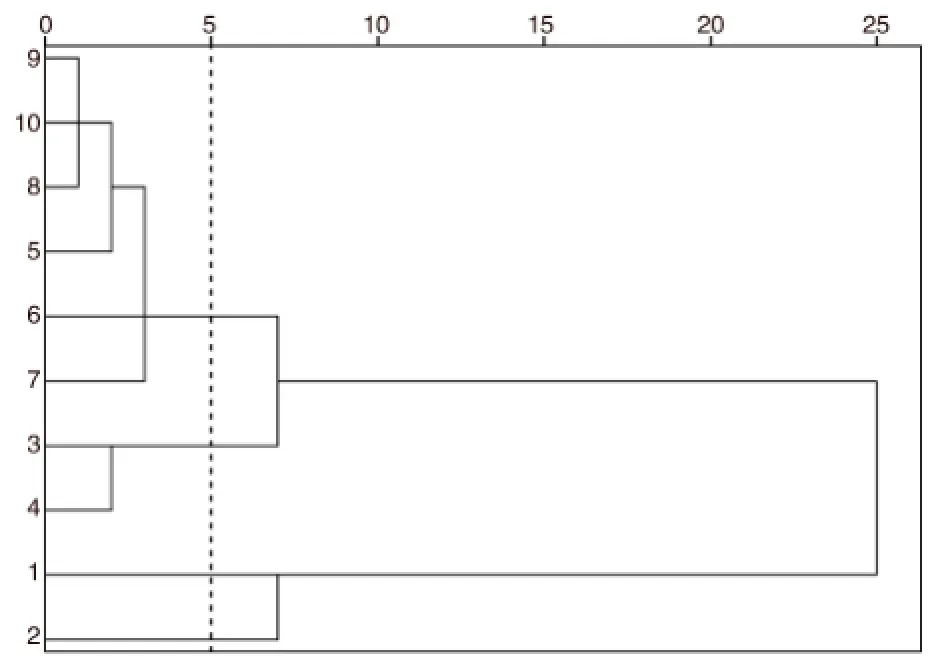
图1 10棵鸡爪槭表型性状的聚类分析Fig.1 Clustering analysis for phenotypic markers of 10 Acer palmatum
图1表明,第1组共有6个单株。本组的表型性状(除翅宽和种形系数)平均值在所有4组中均最小,且相对于其余3组比较均匀,特征不太明显,这对不同利用目标的优良单株选育来说价值不大。第2组共有2个单株,分别是3号单株和4号单株。本组的表型性状值在所有4组中大多数较大,其中翅形系数(2.26)和种形系数(1.47)的值最大;翅宽(7.14 mm)是所有组中最小的,本组的基本特征是翅宽小,翅和小坚果的系数大。第3组和第4组分别仅有1个单株,分别是1号单株和2号单株。第3组翅的表面积较大,翅长仅次于第4组而居第2,翅宽在各组中最大,为8.39 mm,由于翅形系数(2.15)较小说明翅形态趋向于圆形;小坚果体形较大,小坚果宽、小坚果厚和小坚果重在各组中最大,分别达到5.19 mm、4.17 mm和0.047 g,但本组种形系数在4组中较小,为1.31,说明小坚果形态更趋向于圆形;翅长和着生痕长仅次于第4组而居第2位;本组表型性状的基本特征是翅和小坚果的形态均更趋向于圆形,着生痕长较长4组中位居第2。第4组翅的表面积大小与第3组相似,但翅形系数(2.25)较大在4组中位居第2,这说明翅的形态更趋向于椭圆形;小坚果形态方面与第3组相似,也更趋向于圆形;着生痕长在各组中最大,达到了3.94 mm;本组的表型性状基本特征是翅的形态更趋向于椭圆形,小坚果的形态更趋向于圆形,着生痕长在各组中居第1位。
3 结论与讨论
表型性状受环境及栽培因素影响较大,在相同的生态环境下,植物不同性状的多样性能够较好的揭示其遗传本质;种实各性状的遗传稳定性不一致或许是不同自然条件长期作用所造成的[12]。试验材料全部来自同一地点,可以消除生态环境差异引起的翅果表型性状变异。已有的报道表明,不同地区鸡爪槭小坚果长、小坚果宽、小坚果厚和百粒重差异极显著,变异系数分别为10.007%、19.688%、10.089%和23.205%[10],翅果表型性状在不同地理分布区差异很大。研究中,鸡爪槭9个翅果表型性状的变异系数为5.67%~19.71%,该结果表明,鸡爪槭的翅果表型性状在单株间也表现出丰富的变异,这为后期的新品种和优良品种的选育提供了良好的基础。鸡爪槭变异系数最大和最小的表型性状分别是小坚果重(19.71%)和着生痕长(5.68%);形态方面:翅长和翅宽均大于小坚果长和小坚果宽,说明翅形态变异大于小坚果形态变异;小坚果宽(6.78%)和着生痕长(5.68%)的变异系数最小,而小坚果厚(9.35%)的变异系数相对较大,这说明小坚果的纵向粗扁变异大于横向的长短变异。鸡爪槭翅果的质量表型性状是小坚果重,姜景民等[13]认为种子的大小和重量影响种子的发芽速度和发芽率,进而影响到苗木的早期生长,鸡爪槭翅果表型性状的小坚果重(19.71%)和百粒重(23.205%)[10]变异系数较高,这为后期的良种选育提供了一定的理论依据。
表型性状相关性是植物资源研究和利用的重要内容。通过对性状间的相关性研究,确定不同性状间的关系,可以在选择某一个性状的同时预测其对其他性状可能产生的影响或选择压[1]。研究通过对鸡爪槭翅果表型性状相关性的分析发现,鸡爪槭翅果性状间存在比较密切复杂的关系,翅果表型性状间相关关系除翅形系数与种形系数、小坚果厚和小坚果重间没有显著性相关,其余翅果表型性状间具有极显著性相关或显著性相关。因此,在相关性极强的性状间进行选择时,对其中一个性状进行改良,可以同时影响其他性状[14-16]。根据表4,改良小坚果宽和小坚果厚可以对小坚果重产生较大影响。而对相关性不显著的性状进行选择时,一个性状的改良对其他性状影响不大,所以在性状选择上应当有不同的改良方向和选择标准[17-18]。因此在改良性状时需要从遗传改良和提高栽培管理措施等多方面入手。
研究采用质心法将10棵鸡爪槭单株聚类为4组,每个组群形态都有自己的特征,这就从形态特征角度初步明确了个体间的亲缘关系,为不同培育目标优良单株的选择奠定了基础[11]。依据表型性状分类形成的群组(除第1组)有明显的形态特征,加之数量性状一般呈连续性分布,这在育种过程中具有较大的实际应用价值[19]。根据不同的育种目标,利用不同类型的优良基因资源进行基因重组和轮回选择,对于选育优良单株、种质改良创新以及提高种质资源开发利用均具有重要的指导意义。
[1]刁松锋,邵文豪,姜景民,等.无患子实生群体种实表型性状变异研究[J].西北农林科技大学学报,2014,42 (5):75-83.
[2]曾杰,白嘉雨.植物天然居群表型变异研究的几个热点问题[J].广西林业科学,2007,36(2):65-70.
[3]Wheeler N C,Guries R P.Population structure,genetic diversity,and morphologicalvariation in Pinuscontorta Dougl[J].Canadian Journal of Forest Research,1982,12:595-606.
[4]孙荣喜,郑勇奇,张川红,等.不同群体国槐种子表型变异研究[J].河北农业大学学报,2011,34(3):65-70.
[5]黄勇,姚小华,王开良,等.小果油茶种实变异频率及其多样性指数分析[J].江西农业大学学报,2011,33(2):292-299.
[6]王丹,庞春华,高亚卉,等.茶条槭不同海拔种群的表型多样性[J].云南植物研究,2010,32(2):117-125.
[7]中国植物志编辑委员会.中国植物志[M].北京:科学出版社,2008.
[8]许延志.槭树科的地理分布[J].云南植物研究,1996,18 (1):43-50.
[9]陈继卫,沈朝栋,贾玉芳,等.鸡爪槭转色期叶色变化生理研究[J].核农学报,2010,24(1):171-175.
[10]夏涛,张得芳,杨昕,等.江浙地区3个鸡爪槭品种翅果表型变异研究[J].种子,2014(10):37-41.
[11]苗锦山,刘彩霞,戴振建,等.葱种质资源数量性状的聚类分析、相关性和主成分分析[J].中国农业大学学报,2010,15(3):41-49.
[12]Greipsson S,Davy A J.Seed mass and germination behaviour in populations of the dune-building grass Leymus arenarius[J].Annals of Botany,1995,76(5):493-501.
[13]姜景民,虞沫奎,胡世才,等.湿地松种子分级育苗的研究[J].林业科学研究,1996,9(3):290-295.
[14]郭宝林,杨俊霞,李永慈,等.主成分分析法在仁用杏品种主要经济性状选种上的应用[J].林业科学,2000,36 (6):53-56.
[15]田治国,王飞,王朴,等.新疆树上干杏形态学形状的多样性研究[J].西北农林科技大学学报:自然科学版,2012,40(3):147-153.
[16]康永祥,赵宝鑫,贠玉洁,等.毛梾天然群体种实表型多样性研究[J].西北农林科技大学学报:自然科学版,2011,39(9):107-117.
[17]刘芳芳,鲁亚星,于帅,等.黑果腺肋花楸花青素的提取工艺及其稳定性[J].延边大学农学学报,2015(3):196-201.
[18]周旋,何正飚,康宏樟,等.温带-亚热带栓皮栎种子形态的变异及其与环境因子的关系[J].植物生态学报,2013,37(6):481-491.
[19]Baeilieri R,Ducousso A,Kremer A.Genetic morphological ecological and phonological differentiation between Quercu spetraea(Matt.)Label.and Quercus robur L.in a mixed stand of Northwest of France[J].Silvae Genetic,1995,44(1):1-10.
Study on Samara Phenotypic Variation of Acer palmatum
Huang Dongjing,Feng Gang,Liu Jihu,Wan Zhibing
(College of Life and Environment Sciences,Huangshan University,Huangshan 245041)
10 Acer palmatum plants were selected randomly in Huangshan University campus to analyse 9 samara phenotypic traits to study the phenotypic variation within and among single plant.The results showed that the genetic variation coefficients of 9 characters were ranged from 5.68%-19.71%,the correlation coefficients from-0.451 4 to 0.729 4,which indicated that they had complicated relationships with each other.Significant differences in phenotypic traits of four groups divided based on the traits were identified,which provided basic information for accessions with different purposes.Phenotypic variation of Acer palmatum was rich and the variations provide information for the preliminary selection and breeding of seeds for different purposes.
Acer palmatum;samara;phenotypic traits;variation regularity
S792.35
A
1002-2090(2016)05-0036-06
10.3969/j.issn.1002-2090.2016.05.007
2015-12-01
国家自然基金(31400563);安徽省大学生创新训练项目;安徽省优秀青年人才基金项目(2012SQRL185)资助。
黄东静(1993-),女,黄山学院生命与环境科学学院林学专业2012级本科生。
万志兵,男,副教授,E-mail:wanzb626@hsu.edu.cn。
