江西金盆山自然保护区种子植物区系研究
2017-01-03李健星刘以珍李恩香
李健星,刘以珍,葛 刚,陈 江,李恩香
(南昌大学 生命科学学院,南昌330031)
江西金盆山自然保护区种子植物区系研究
李健星,刘以珍,葛 刚,陈 江,李恩香*
(南昌大学 生命科学学院,南昌330031)
江西金盆山自然保护区位于江西省龙南县、全南县与信丰县的交汇处,为武夷山脉与南岭山脉之间的过渡区域,保护区植物种类丰富、区系地理成分复杂。经野外实地调查、查阅相关文献资料,对保护区内种子植物的科、属、种分布区类型进行统计分析。结果表明:(1)金盆山保护区共有野生种子植物161科659属1 474种,其中裸子植物6科7属7种,占江西裸子植物总种数(83种)的8.43%,被子植物155科652属1 467种,占江西被子植物总种数(4 369种)的33.58%。(2)区系分析显示,金盆山保护区内科的分布区类型中热带分布和温带分布分别占总科数(除去世界分布)的64.22%和35.77%,热带分布占优势地位;属的分布区类型中热带分布和温带分布分别占总属数(除去世界分布)的50.95%和47.55%,二者接近平衡;种的分布区类型中热带分布种和温带分布分别占总种数(除去世界分布)的33.98%和37.15%,而且中国特有种419种,占总种比的28.88%。(3)金盆山保护区种子植物具有单种属、寡种属多,植物区系起源古老的特点,其中有寡种属212属,占总属比的32.17%;单种属374属,占总属比的56.75%;其中寡种属所含种数537种,占总种比的36.43%。(4)与邻近6个保护区的植物区系比较表明,金盆山保护区与武夷山脉的联系较南岭山脉更近,而且金盆山保护区表现出比邻近的阳际峰等武夷山脉地区热带性质更明显。
植物区系;种子植物;江西金盆山
植物区系(flora)是指某一地区,或者某一时期,某一分类群,某类植被等所有植物种类的总称,是一个地区的植物长期发展适应其所处的自然环境的结果[1]。江西植被类型复杂,同时组成植被的植物区系成分也特别丰富,这与江西自然条件与植物区系的历史起源均有密切的关系,无论在亚热带特有种属、孑遗植物、古热带植物区系成分上以及其与世界各植物区系的联系上,均有其独特性[2]。江西金盆山自然保护区位于江西省龙南县、全南县与信丰县的交汇处,为武夷山脉与南岭山脉之间的过渡区域,属于中亚热带常绿阔叶林区,保护区内气候湿润,地形复杂,使得保护区植物种类丰富、区系地理成分复杂,在中国植物区系分区系统中属于赣南-湘东丘陵亚地区[3]。近年来,关于江西植物区系的研究越来越多,主要有庐山、马头山、武夷山、井冈山、三清山、铜钹山等保护区以及江西野生种子植物的研究[4-11],对金盆山保护区也有一些研究[12-14],但未见有关金盆山植物区系的报道。本文在野外调查的基础上,查阅相关文献资料,初步分析了金盆山的种子植物区系特征,为保护区植物资源的保护和利用奠定基础。
1 自然概况
江西金盆山自然保护区地处N25°20′~25°25′,E114°19′~114°34′。属武夷山山脉,地势起伏不断,由南而北倾斜,但都多为海拔300~500 m的低山,主峰为陀秀岽海拔高953.5 m,规划总面积3 711.7 hm2,森林覆盖率99.98%。气候上属亚热带湿润季风区,降雨量充沛,拥有优越的水热条件,年平均气温19.5 ℃,年平均降雨量1 600 mm,极端最高气温39.4 ℃,最低气温-4 ℃,无霜期285 d。土壤为山地黄壤、紫色土和红壤,地带性土壤为红壤。岩石主要由花岗岩和砂岩构成。金盆山属中亚热带南部,水热条件优越,适于多种植物生长。植被类型为江西南部典型的亚热带常绿阔叶林区域,金盆山自然保护区是该区域中仅有的几个认为干扰较少、自然植被完整的区域之一。
2 研究方法
2015年7月,根据金盆山的地势、植被类型以及人类活动等因素,设置了5条样线进行植物标本采集和区系调查。参考样线进行野外实地调查,记录样线周围的物种,采集植物标本并拍照记录。
标本存放于南昌大学标本馆(JXU)。结合相关的文献资料以及《中国植物志》[15]、《Flora of China》[16]、《江西植物志》[17-18]、《江西种子植物名录》[19]进行植物标本鉴定。金盆山种子植物名录参照《Flora of China》的恩格勒系统编制。
参考吴征镒[1,3,20-21]对中国种子植物区系地理划分的原则,对金盆山野生种子植物进行区系分析统计。
3 结果与分析
3.1 金盆山保护区种子植物的物种多样性
初步统计显示,金盆山有野生种子植物共161科659属1 474种,其中裸子植物6科7属7种,被子植物155科652属1 467种(按Hutchinson系统)。其中裸子植物种数占保护区种子植物总数的0.47%,被子植物种数占99.53%(表1),可见在整个保护区的被子植物占有绝对优势地位,也是保护区内植被的主要构建者。
根据各科包含种数,将金盆山的种子植物科分为5个等级,分别是单种科(1种)、寡种科(2~4种)、小科(5~9种)、中科(10~19种)和大科(≥20种)。
从表2可以得出,保护区内寡种科所占比例最大,随后是小科、单种科、中科和大科。但大科有18

表1 金盆山保护区种子植物科、属、种数量分析

表2 金盆山保护区种子植物科内种的数量分析
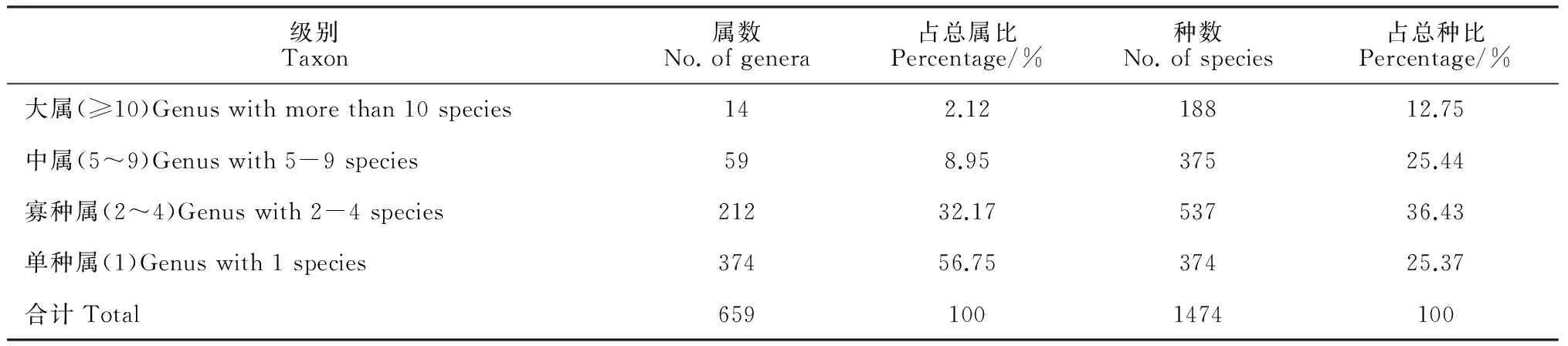
表3 金盆山保护区种子植物属内种的数量分析
科756种,占保护区种子植物所含种的51.29%,可见大科对保护区的区系建成有着明显的支撑作用,其中有禾本科(Gramineae)(55/93)(属数/种数,后同)、菊科(Compositae)(48/82)、豆科(Fabaceae)(33/73)、蔷薇科(Rosaceae)(24/66)、樟科(Lauraceae)(8/56)、壳斗科(6/46种)等,这些大科同时也是中国植物区系中的大科,可见该区系与中国植物区系源远流长,属于白垩纪-老第三纪传承至今的壳斗科是保护区内阔叶林的主要成分。另外本区系中寡种科和小科在科一级中占优势地位,体现了该区系的古老性,其中近于孑遗的蓝果树科(Nyssaceae)在山谷地带呈现群落分布,在乔木层中为优势种,呈孑遗性质的杉科(Taxodiaceae)、三尖杉科(Cephalotaxaceae)、三白草科(Saururaceae)在保护区内大量分布。
在植物区系研究中,植物属的分布区类型比科的分布区类型更有意义,属的分布区类型能够比较具体地反映植物界在进化过程中的分异情况,是进一步研究植物区系的起源、演化和分布区类型形成的起点或楔子[3]。在植物系统分类学上,属内的种具有平行的进化趋势,因此研究植物属的区系成分能更有效地揭示出植物区系的特征。根据各属包含种数的数量,再结合实际情况,将金盆山种子植物划分为4个等级,包括单种属(1种)、寡种属(2~4种)、中属(5~9种)与大属(≥10种)。
由表3可知,该区系植物中单种属最多,占总属的56.75%;其次是寡种属,占总属数的32.17%;中属有59属,占总属的8.95%;大属有14属,仅占总属数的2.12%。从各类型属所含种数来看,寡种属所含种数最多,其次是中属、单种属,大属所含种数最少,占总种数的12.75%。大属包括有堇菜属(Viola)、蓼属(Polygonum)、悬钩子属(Rubus)、锥栗属(Castanopsis)、青冈属(Cyclobalanopsis)、榕属(Ficus)、冬青属(Ilex)、蛇葡萄属(Ampelopsis)和苔草属(Carex)等。
3.2 植物区系的地理成分分析
3.2.1 科的地理成分分析 按照吴征镒对种子植物地理成分的划分标准,对金盆山种子植物区系科的地理成分进行了统计分析。江西金盆山种子植物区系中的161科共有9个分布区类型和10个变型(表4),其中世界广布科52科;热带科70科(2-7型),占总科数的64.22%;温带科(8-14型)39科,占总科数的35.77%。R/T(热带性质科/温带性质科)约为1.80,得出该区系在科的分布呈现明显的热带性质,与保护区在中国植物区系分区系统中属于赣南-湘东丘陵亚地区所体现出来的典型中亚热带气候相符合。
世界广布科有52种,包含有毛茛科(Ranunculaceae)、菊科、蔷薇科等这些大科等,这些科在金盆山植物区系中有着不可替代的作用。
热带性质的科有70科,其中泛热带分布的有41科,占种子植物总科数的37.61%。主要有樟科、山茶科(Theaceae)等。虽从所包含种的数量看该分布类型小于世界分布科,但因含有该地区的优势科如樟科、山茶科以及山矾科(Symplocaceae)等,在保护区的森林生态系统中仍具有较高的重要值。东亚(热带、亚热带)及热带南美间断有9科,包含有木通科(Lardizabalaceae)、杜英科(Elaeocarpaceae)、冬青科(Aquifoliaceae)等。旧世界热带分布的有海桐花科(Pittosporaceae)、八角枫科(Alangiaceae)和芭蕉科(Musaceae)3科,其变型——热带亚洲、非洲(或东非,马达加斯加)和大洋洲间断分布的水蕹科(Aponogetonaceae)。热带亚洲至热带大洋洲分布有2科,分别是交让木科(Daphniphyllaceae)、马前科(Loganiaceae)和姜科(Zingiberaceae)。热带亚洲至热带非洲分布的只有其变型南非(主要是好望角)分布的杜鹃花科(Ericaceae)。热带亚洲分布有3个变型,其中缅甸、泰国至华西南分布只含有伯乐树科(Bretschneideraceae),越南(或中南半岛)至华南(或西南)分布只有大血藤科(Sargentodoxaceae),全分布区东达新几内亚分布的只有清风藤科(Sabiaceae)。
温带性质的有39科,其中北温带分布的有29科,主要有松科(Pinaceae)、壳斗科、藤黄科(Hypericaceae)等。壳斗科植物如华南栲(Castanopsisconcinna)、米槠(C.carlesii)、甜槠(C.eyrei)、青冈(Cyclobalanopsisglauca)、苦槠(C.sclerophylla)等在本区中常为群落的建群种占有重要地位。东亚和北美洲间断分布,包含有木兰科(Magnoliaceae)、五味子科(Schisandraceae)、蓝果树科(Nyssaceae)等。旧世界温带分布只有菱科(Hydrocaryaceae)。东亚分布有3科,包含有三尖杉科(Cephalotaxaceae)、猕猴桃科(Actinidiaceae)和旌节花科(Stachyuraceae)。
3.2.2 属的地理成分分析 植物属的区系成分与科比较它能更彻底地反映出一个地区植物区系的特点[22]。按照吴征镒等对中国种子植物属的分布区类型划分方法,将金盆山野生种子植物的659属划分为14个分布型,16个亚型(表4),其中热带分布属(2-7型)占金盆山总属比的50.95%;温带分布属(8-14型)占总属比的47.55%;R/T值为1.07,而中国特有属9种占总属比的1.52%。可见,该区系中种子植物地理成分比较复杂;热带分布比温带分布稍高,金盆山属的区系表现出一定的热带分布,这一点与科相同。
世界分布属有68属,大多数都是草本植物包含有荠属(Capsella)、碎米荠属(Cardamine)、千里光属(Senecio)等。

表4 金盆山保护区种子植物科、属分布区类型统计
注:*表示百分比中除去世界分布型
Note:Percentage of total families or genera excludes the cosmopolitan pattern
热带分布的有301属,占金盆山总属的50.95%(除去世界分布属,下同)。泛热带分布的有133属,包括1个变型,占金盆山总属数的22.51%,是金盆山属分布的第一大类型。该分布正型包含有紫金牛属(Ardisia)、山矾属(Symplocos)、紫珠属(Callicarpa)等132属。热带亚洲、非洲和中至南美洲间断分布的凤眼蓝属(Eichhornia)1属。东亚(热带、亚热带)及热带南美间断分布的有17属,包括有樟属(Cinnamomum)、木姜子属(Litsea)、柃属(Eurya)等。旧世界热带分布的有39属,该分布区类型正型包括有野桐属(Mallotus)、楼梯草属(Elatostema)、杜茎山属(Maesa)等38属。另还包含热带亚洲、非洲(或东非,马达加斯加)和大洋洲间断分布的水蕹属(Aponogeton)。热带亚洲至热带大洋洲分布的有36属包括有新木姜子属(Neolitsea)、野牡丹属(Melastoma)、桃金娘属(Rhodomyrtus)等。热带亚洲至热带非洲分布的有17属,包括有杨桐属(Adinandra)、野茼蒿属(Crassocephalum)、香茶菜属(Rabdosia)等。热带亚洲分布的有59属,其正型分布包括有润楠属(Machilus)、青冈属(Cyclobalanopsis)、马铃苣苔属(Oreocharis)等19属。另还有7个变型,这些变型都表现出一定程度的原始性和古老性,其中爪哇(或苏门答腊)、喜马拉雅至华南、西南间断或星散分布的有蕈树属(Altingia)、古热带第三纪孑遗的木荷属(Schima)以及草珊瑚属(Sarcandra)3属。热带印度至华南(特别滇南)分布的只有伯乐树属(Bretschneidera)。越南(或中南半岛)至华南(或西南)分布的只有大血藤属(Sargentodoxa)。西马来,基本上在新华莱士线以西北可达中南半岛或印东北或热带喜马拉雅,南达苏门答腊分布的木莲属(Manglietia)、含笑属(Michelia)、楠木属(Phoebe)等26属,东马来,即新华莱士线以东,但不包括新几内亚及东侧岛屿分布的只有秤钩风属(Diploclisia),东马来,即新华来士线以东,包括新几内亚及东侧岛屿分布的苦荬菜属(Ixeris),全分布区东南达西太平洋诸岛弧,包括新喀里多尼亚和斐济分布的构属(Broussonetia)、赤车属(Pellionia)、蛇根草属(Ophiorrhiza)等6属。
温带分布的有281属,占总属比的47.55%。北温带分布的有109属,占金盆山总属数的18.44%,是金盆山属分布的第二大类型。属于这一分布类型正型的有红豆杉属(Taxus)、蓼属(Polygonum)、茜草属(Rubia)等100属。另还有北温带和南温带间断(泛温带)分布包含有栎属(Quercus)、胡颓子属(Elaeagnus)、枫属(Acer)等9属。欧亚和温带南美洲间断分布有胡桃属(Juglans)1属。东亚及北美分布这一分布区类型是最早被植物学家所认识的分布区类型,本区系中属于这一分布类型的有49属,其中该正型分布的包括有木兰属(Magnolia)、五味子属(Schisandra)、蓝果树属(Nyssa)等48属。由于它们在太平洋洋底达扩大以前就已经广泛分布于欧亚大陆和北美两大板块,这些都是古老的残遗植物。变型为东亚和墨西哥间断分布有大丁草属(Leibnitzia)1属。旧世界温带分布的有38属,其中该正型分布的包括有淫羊藿属(Epimedium)、马甲子属(Paliurus)、活血丹属(Glechoma)等37属。变型是地中海区、西亚与东亚间断分布的小窃衣属(Torilis)。温带亚洲分布本区系中只分布其正型共8属包含有虎杖属(Reynoutria)、白鹃梅属(Exochorda)、女菀属(Turczaninowia)等8属。地中海区、西亚至中亚分布的有3属,其中该正型分布的包括有黄连木属(Pistacia)、常春藤属(Hedera)共2属。另还包含有地中海至温带-热带亚洲、大洋洲和南美洲间断这一变型的牻牛儿苗属(Erodium)1属。东亚分布的有74属,其中该正型分布的包括有猕猴桃属(Actinidia)、油桐属(Vernicia)、四照花属(Dendrobenthamia)等39属。另还包括两个变型中国-喜马拉雅(SH)分布的有八月瓜属(Holboellia)、雪胆属(Hemsleya)、冠盖藤属(Pileostegia)等共4属;中国-日本(SJ)分布的有博落回属(Macleaya)、野鸦椿属(Euscaphis)、化香树属(Platycarya)等31属。
中国特有分布,特有属的研究对于探讨某地区植物区系的性质、区划、起源、变迁乃至与其它植物区系的亲缘关系都有着十分重要的价值。中国种子植物特有属分散在种子植物进化的各个阶段,古老属和新生属共存。中国的种子植物特有性很高,包括有248个特有属,为总属数的7.9%。
本区系中属于这一类型的有9属,占保护区总属的1.52%,包含有杉木属(Cunninghamia)、拟单性木兰属(Parakmeria)、血水草属(Eomecon)、藤山柳属(Clematoclethra)、半枫荷属(Semiliquidambar)、喜树属(Camptotheca)、青钱柳属(Cyclocarya)等。
3.2.3 种的地理成分分析 种的区系分析能够体现出某一地区的具体区系特征,是植物区系分析的最基本单元,按照吴征镒等对中国种子植物的分布区类型划分方法,参考《Flora of China》中种的分布,将金盆山野生种子植物的1 474种划分为10个分布型,11个亚型(表4),其中热带分布种(2-7型)占金盆山总种比的33.98%;温带分布种(8-14型)占总种比的37.15%;R/T值为0.91,而中国特有种419种占总种比的28.88%。可见,该区系中种子植物地理成分比较复杂;热带分布比温带分布稍低。
世界分布种有23种,绝大部分是草本植物。主要有碎米荠(Cardaminehirsuta)、皱果苋(Amaranthusviridis)、一年蓬(Erigeronannuus)、双穗雀稗(Paspalumdistichum)等,主要借助动物或者河流进行传播。
热带分布的有493种。其中泛热带分布的有73种,占保护区总数的5.03%。其正型分布有叶下珠(Phyllanthusurinaria)、鬼针草(Bidenspilosa)、母草(Linderniacrustacea)等53种,旧世界热带分布的5种,热带亚洲、大洋洲(至新西兰)和中至南美洲(或墨西哥)间断分布的有豨莶(Sigesbeckiaorientalis)、鸭舌草(Monochoriavaginalis)、浮萍(Lemnaminor)等15种。热带亚洲、非洲和中至南美洲间断分布的有合萌(Aeschynomeneindica)、地胆草(Elephantopusscaber)、益母草(Leonurusjaponicus)等5种。旧世界热带分布的有黄花草(Arivelaviscosa)、狗尾草(Setariaviridis)、筒轴茅(Rottboelliacochinchinensis)等5种。热带亚洲和热带大洋洲分布有千斤藤(Stephaniajaponica)、牛膝(Achyranthesbidentata)、茅膏菜(Droserapeltata)等7种,中国(西南)亚热带和新西兰间断分布的有绞股蓝(Gynostemmapentaphyllum)、五月茶(Antidesmabunius)、耳草(Hedyotisauricularia)等51种。热带亚洲至热带非洲分布的有青葙(Celosiaargentea)、八角枫(Alangiumchinense)、天胡荽(Hydrocotylesibthorpioides)等8种。华南、西南至印度和热带非洲间断分布的有尼泊尔蓼(Polygonumnepalense)、鱼眼草(Dichrocephalaintegrifolia)、水蜈蚣(Kyllingapolyphylla)等10种。热带亚洲分布339种,占保护区总数的23.36%,在保护区热带分布中占主要地位。其正型分布的有阴香(Cinnamomumburmanni)、鱼腥草(Houttuyniacordata)、柏拉木(Blastuscochinchinensis)等235种,热带印度至华南(特别滇南)分布的有少花桂(Cinnamomumpauciflorum)、厚皮香(Ternstroemiagymnanthera)、灰毛大青(Clerodendrumcanescens)等14种,缅甸、泰国至华西南分布的有银木荷(Schimaargentea)、 刺果卫矛(Euonymusacanthocarpus)、青榨枫(Acerdavidii)等3种,越南(或中南半岛)至华南(或西南)分布的有黑老虎(Kadsuracoccinea)、乌药(Linderaaggregata)、杨桐(Adinandramillettii)等87种。
温带分布的有539,占总种比的37.15%。北温带分布的有115种,其正型分布的萹蓄(Polygonumaviculare)、路边青(Geumaleppicum)、牛蒡(Arctiumlappa)等16种,北温带和南温带间断(泛温带)分布的有播娘蒿(Descurainiasophia)、石竹(Dianthuschinensis)、两栖蓼(Polygonumamphibium)等83种,欧亚和温带南美洲间断分布的有荠(Capsellabursa-pastoris)、雀舌草(Stellariaalsine)、野慈姑(Sagittariatrifolia)等16种。东亚及北美间断分布的有蔊菜(Rorippaindica)、鸡眼草(Kummerowiastriata)、蛇床(Cnidiummonnieri)等27种。
东亚分布的有397种,其正型分布的有五味子(Schisandrachinensis)、毛茛(Ranunculusjaponicus)、木通(Akebiaquinata)等309种,中国-日本分布的有新木姜子(Neolitseaaurata)、铁线莲(Clematisflorida)、博落回(Macleayacordata)等88种。
中国特有分布的有419种,占总数的28.88%。根据这些种的现代地理分布、生态适应能力、传播等,将保护区的中国特有种分为11个变型(表5)。从表5可以得出,在11个亚型中,西南-华中-华东-华南分布种有128种,占保护区特有种的30.55%,可以看出保护区与西南、华东、华南有着广泛的联系。表中可以看出金盆山与华东、华南的联系最为密切,这也与金盆山的地理位置相吻合,保护区处于华中南部,与华东、华南紧密相连。

表5 金盆山保护区中国特有种分布亚型
3.3 金盆山保护区与邻近6个自然保护区种子植物区系的比较
植物区系的形成是植物在一定的自然历史环境中发展演化和时空分布的综合反映。在一定程度上能够为历史环境变迁提供证据,反映现代自然地理条件,因此用植物区系地理成分做指标可比较不同地区植物区系的亲缘关系。金盆山保护区处于武夷山脉与南岭山脉之间的过渡区域,在中国植物区系分区系统中属于赣南-湘东丘陵亚地区,与其它地区的植物区系有着广泛的联系,与邻近地区的植物区系存在着一定的相似性。为进一步了解金盆山保护区的植物区系特征,以及与邻近地区的植物区系的关系,将金盆山保护区与江西武夷山、福建武夷山、南岭保护区、九连山、阳际峰、马头山属的分布区类型进行统计与比较,探讨金盆山保护区与武夷山脉和南岭山脉之间的联系,保护区种子植物属区系参考科考记载和文献[23-26]。
从表6可以看出金盆山保护区属的R/T值与相对其距离较远的马头山和阳际峰自然保护区相近,与之相近的九连山差异较大,与武夷山脉保护区的R/T值更接近,相对于南岭山脉则差值较大。保护区的R/T值为1.07,即热带性质和温带性质相平衡,这也与保护区所处的地理位置有关,属于武夷山脉与南岭山脉之间的过渡区域,区系性质上是热带向温带过渡的类型。

表6 金盆山保护区与相邻6个自然保护区种子植物属的分布区类型比较
根据种子植物属的分布区类型所占比例,与其他地区植物区系作比较。可以看出金盆山保护区与武夷山脉保护区属的分布区类型具有很好的一致性,与南岭保护区有一些差异。金盆山保护区植物区系特征与武夷山脉更接近。
种水平上,与邻近的阳际峰国家级自然保护区相比,热带性质更明显,其中阳际峰的热带科319科,温带科472科,R/T值为0.68[25],而金盆山保护区的R/T值为0.91。这与他们的地理位置相关,金盆山保护区比阳际峰更靠近南方,其热带性质比温带性质更显著。
4 结 论
4.1 保护区内植物物种种类丰富
金盆山保护区共有野生种子植物161科659属1474种,占江西种子植物总种数(4 452种)的33.12%;裸子植物6科7属7种,占江西裸子植物总种数(83种)的8.43%;被子植物155科652属1 467种占江西被子植物总种数(4 369种)的33.58%。
4.2 保护区内区系类型复杂
从科的区系类型看,保护区的161科共有9个分布区类型和10个变型(表4),其中世界广布科52科;热带科70科(2-7型),占总科数的64.22%;温带科(8-14型)39科,占总科数的35.77%。从属的区系类型看,保护区的659属共有14个分布区类型,16个变型(表4),其中热带分布属(2-7型)占金盆山总属比的50.95%;温带分布属(8-14型)占总属比的47.55%,而中国特有属9种占总属比的1.52%。其中温带分布以北温带分布为主,热带分布以泛热带分布为主。从种的区系类型看,保护区的1 474种划分为10个分布型,11个亚型(表4),其中热带分布种(2-7型)占金盆山总种比的33.98%;温带分布种(8-14型)占总种比的37.15%,而中国特有种419种占总种比的28.88%。
4.3 保护区内植物区系主要为热带性向温带性过渡的类型
金盆山科的分布区类型中热带和温带分布分别占总科数的64.22%和35.77%,R/T(热带性质科/温带性质科)约为1.80,热带分布占优势地位,得出该区系在科的分布呈现明显的热带性质,与保护区在中国植物区系分区系统中属于赣南-湘东丘陵亚地区所体现出来的典型中亚热带气候相符合。金盆山属的分布区类型中热带和温带分布分别占总属数的50.95%和49.77%,二者接近平衡,种的R/T值为0.91,由以上数据可以推断保护区的区系类型基本上是属于亚热带性质,是热带向温带过渡的类型。
该区系植物中单种属最多,占总属的56.75%;其次是寡种属,占总属数的32.17%;中属有59属,占总属的8.95%,大属(≥10)有14属,仅占总属数的2.12%。从各类型属所含种数来看,寡种属所含种数最多,其次是中属、单种属,大属所含种数最少,占总种数的12.75%。
4.4 保护区内单种属、寡种属多,植物区系起源古老
保护区种子植物中有寡种属212属,占总属比的32.17%;单种属374属,占总属比的56.75%。其中寡种属所含种数537种,占总种比的36.43%。保护区内有大量属于间断分布的属,例如,东亚及北美间断分布的木兰属、五味子属、蓝果树属等,爪哇(或苏门答腊)喜马拉雅间断或星散分布到华南、西南分布的有蕈树属、木荷属,北温带和南温带间断分布的播娘蒿属(Descurainia)、栎属、胡颓子属、枫属等,这些都是一些古老或比较古老的类群,说明了该区系起源具有一定的古老性。另该区系中包含有一些古物种裸子植物中的杉木属产于老第三纪。被子植物中起源古老的成分尤其丰富,白垩纪初期至晚期就已经出现的有木兰科、樟科、壳斗科、毛茛科、卫矛科(Celastraceae)、槭树科(Aceraceae)等,这些植物在保护区内大量分布,是本区内植物群落的主要建群种,在保护区植物区系中扮演着重要角色;在老第三纪建立的科,如八角枫科、苦木科(Simaroubaceae)、清风藤科、山茶科等在保护区很常见。
4.5 保护区与邻近保护区植物区关系
与邻近6个保护区植物区系的比较表明,金盆山保护区与武夷山脉的联系较南岭山脉更近,而且金盆山保护区表现出比邻近的阳际峰等武夷山脉地区热带性质更明显。
[1] 吴征镒. 世界种子植物科的分布区类型系统[J]. 云南植物研究,2003,25(3): 245-257.
WU Z Y. The areal-types of the world families of seed plants [J].ActaBotanicaYunnanica, 2003, 25(3): 245-257.
[2] 江西植物志编辑委员会.江西植物志,第1卷[M]. 南昌:江西科学技术出版社,1993.
[3] 吴征镒,孙 航,周浙昆,等. 中国种子植物区系地理[M]. 北京:科学出版社,2010.
[4] 樊忠良. 庐山植物的区系特征和植被分布[J]. 黄冈师专学报(自然科学版),1994,14(1): 52-55.
FAN Z L. The features of flora and the distribution of vegetation at Lushan Mountain [J].JournalofHuanggangNormalUniversity(Nat. Sci. Edi.),1994, 14(1): 52-55.
[5] 裘利洪,刘仁林,施建敏,等. 马头山自然保护区种子植物区系研究——Ⅱ.属的区系分析[J]. 江西农业大学学报,2005,27(4):590-595.
QIU L H,LIU R L,SHI J M,etal. A study on the flora of seed plants in Matou Mountain Nature Reserve Ⅱ. Analysis of genus flora [J].ActaAgriculturaeUniversitisJiangxiensis,2005,27(4):590-595.
[6] 刘仁林,裘利洪,施建敏,等. 马头山自然保护区种子植物区系研究——Ⅰ.科的区系分析[J]. 江西农业大学学报,2005,27(3):361-367.
LIU R L,QIU L H,SHI J M,etal. A study on the flora of spermatophyte in the Nature Reserve of Matou Mountain Ⅰ. Flora of family [J].ActaAgriculturaeUniversitisJiangxiensis,2005,27(3): 361-367.
[7] 陈世品. 武夷山风景区种子植物区系研究[J]. 福建林业科技,2004,31(2):16-19.
CHEN S P. Seed plant flora of Wuyishan scenery spot [J].JournalofFujianForestryScienceandTechnology,2004, 31(2): 16-19.
[8] 刘仁林,唐赣成.井冈山种子植物的区系研究[J]. 武汉植物研究,1995, 13(3):210 -218.
LIU R L,TANG G C. Study on flora of spermatophyte in Jinggang Mountain [J].JournalofWuhanBatanicalResearch,1995, 13(3): 210-218.
[9] 臧 敏,黄立发.江西三清山维管束植物区系分析[J]. 亚热带植物科学,2010,39(3):55-62.
ZANG M,HUANG L F. Flora analysis on vascular plants of Sanqingshan Mountain in Jiangxi Province [J].SubtropicalPlantScience,2010, 39(3): 55-62.
[10] 涂业苟,余能富,吴能良,等. 江西铜钹山自然保护区种子植物区系初步研究[J],江西农业大学学报2012,34(4):754-761.
TU Y G,YU N F,WU N L,etal. A preliminary study on the flora of seed plants of vegetation in Tongboshan Nature Reserve of Jiangxi Province [J],ActaAgriculturaeUniversitisJiangxiensis,2012, 34(4): 754-761.
[11] 邓贤兰,吴 杨,刘玉成,等. 江西野生种子植物区系多样性及其基本特征[J].植物科学学报2012,30(1):22-30.
DENG X L, WU Y, LIU Y C,etal. Floristic diversity and fundamental features of wild seed plants in Jiangxi Province [J].PlantScienceJournal,2012, 30(1): 22-30.
[12] 邱凤英,李光运,肖复明,等. 江西金盆山林区常绿阔叶林群落生物量研究[J].江西林业科技,2011,4:1-5.
QIU F Y,LI G Y,XIAO F M,etal. Floristic diversity and fundamental features of wild seed plants in Jiangxi Province[J].JiangxiForestryScienceandTechnology,2011, 4: 1-5.
[13] 程齐来, 李洪亮, 张道英,等. 江西信丰金盆山林场药用植物资源调查研究[C]//. 全国第九届天然药物资源学术研讨会论文集,2010: 117-120.
[14] 曹展波,林 洪,罗坤水,等. 江西金盆山林区米槠生长过程与幼林生长效应[J]. 江西林业科技,2014,42(5):7-9.
CAO Z B,LIN H,LUO K S,etal. Growth process and young plantation growth effect ofCastanopsiscarlesiiin Jinpenshan forest region of Jiangxi Province [J].JiangxiForestryScienceandTechnology,2014, 42(5): 7-9.
[15] 中国植物志编辑委员会. 中国植物志(第1-80卷)[M]. 北京:科学出版社,1959-2004.
[16] Flora of China编委会. Flora of China [EB/OL]. 1989-2013. http://foc.eflora.cn/
[17] 江西植物志编辑委员会. 江西植物志,第2卷[M]. 北京:中国科学技术出版社, 2004.
[18] 江西植物志编辑委员会. 江西植物志,第3卷[M]. 南昌:江西科学技术出版社, 2013.
[19] 刘仁林,张志翔,廖为明. 江西种子植物名录[M]. 北京:中国林业出版社,2010:9.
[20] 吴征镒.世界种子植物科的分布区类型系统的修订[J].云南植物研究, 2003,27(5): 535-538.
WU Z Y. The areal-types of the world families of seed plants[J].ActaBotanicaYunnanica,2003, 27(5): 535- 538.
[21] 吴征镒,周浙昆,孙 航,等. 种子植物分布区类型及其起源和分化[M]. 昆明:云南科技出版社,2006.
[22] 李锡文. 中国种子植物区系统计分析[J]. 云南值物研究1996,18(4): 1-3.
LI X W. Floristic statistics and analyses of seed plants from China [J].ActaBotanicaYunnanica,1996, 18(4): 1-3.
[23] 姚振生,曹 岚,刘信中,等. 江西武夷山自然保护区种子植物区系研究[M]//.刘信中, 方福生. 江西武夷山自然保护区科学考察集. 北京:中国林业出版社,2001: 44-54.
[24] 姚振生,刘信中,曹 岚,等. 九连山自然保护区种子植物区系研究[M]//.刘信中,肖忠优,马建华. 江西九连山自然保护区科学考察与森林生态系统研究. 北京:中国林业出版社,2002: 106-125.
[25] 季梦成,严雄梁,吴璐璐,等. 江西阳际峰自然保护区植物资源[M]//.郭英荣,江波,王英永. 江西阳际峰自然保护区综合科学考察报告. 北京:科学出版社,2010: 46-95.
[26] 刘仁林,裘利洪,揭正平,等. 江西马头山自然保护区植物资源[M]//.刘信中,傅 清. 江西马头山自然保护区科学考察与稀有植物群落研究. 北京:中国林业出版社,2006: 49-68.
(编辑:潘新社)
Study on the Flora of Seed Plants in Jinpenshan Nature Reserve,Jiangxi Province
LI Jianxing, LIU Yizhen, GE Gang, CHEN Jiang, LI Enxiang*
(College of Life Sciences, Nanchang University, Nanchang 330031, China)
Jinpenshan nature reserve is located at the junction of Longnan County, Nanquan County and Xinfeng County, Jiangxi Province. It is the transition zone between Wuyi Mountains and Nanling Mountains. The nature reserve possesses a variety of species and complex geographical floristic elements. Based on field studies and relevant scientific literatures, this study has statistics and analysis the families, genera and species in Jinpenshan. Results showed: (1) 1 474 species of wild seed plants, distributed in 659 genera of 161 families are found in Jinpenshan. Among them, there are 7 gymnosperm species belonging to 7 genera of 6 families, accounting for 8.43% of total gymnosperm species in Jiangxi (83). Floristic analysis showed, there are 1 467 angiosperm species belonging to 652 genera of 155 families, accounting for 33.58% of total angiosperm species in Jiangxi (4 369). (2) The families of tropical and temperate distribution patterns make ups 64.22% and 35.77% of the total family number (removal Cosmopolitan, the same below), respectively. Tropical distribution is in the dominant position. While at genus level, the tropical and temperate distribution patterns make up 50.95% and 47.55%, respectively. They are nearly equal. At species level,the tropical and temperate distribution pattern makes up 33.98% and 37.15%, respectively. While 419 species endemic to China,accounting for 28.88% of total species in Jinpenshan. (3) The flora possess lots of monotypic and oligotypic genera, with an ancient origin. There are 212 genera of oligotypic genera, accounting for 32.17% of the total, 374 genera of monotypic genera, accounting for 56.75% of the total and 537 genera of minor genera, accounting for 36.43% of the total. (4) Compared with the nearby 6 plant floras, there is a closer relationship between Jinpenshan and Wuyi montains than that of Nanling mountains. More tropical elements was observed in Jinpenshen than that of Yangbiaofeng of Wuyi mountains.
flora; seed plant; Jinpenshan, Jiangxi Province
1000-4025(2016)11-2322-10
10.7606/j.issn.1000-4025.2016.11.2322
2016-07-29;修改稿收到日期:2016-10-20
江西省教育厅科学技术研究项目(150161);科技部基础性研究专项(2015FY110200)
李健星(1992-),女,硕士研究生,主要从事系统与进化植物学研究。E-mail:jxlee140820@163.com
*通信作者:李恩香,副教授,硕士导师,主要从事系统与进化植物学研究。E-mail: enxl@163.com
Q948.5
A
