短命植物叶片生源要素的化学计量特征及异速关系
2017-01-03刘建国刘卫国霍举颂
刘建国,林 喆,刘卫国*,霍举颂
(1 新疆大学 资源与环境科学学院,乌鲁木齐 830046;2 绿洲生态教育部重点实验室,乌鲁木齐 830046)
短命植物叶片生源要素的化学计量特征及异速关系
刘建国1,2,林 喆1,2,刘卫国1,2*,霍举颂1,2
(1 新疆大学 资源与环境科学学院,乌鲁木齐 830046;2 绿洲生态教育部重点实验室,乌鲁木齐 830046)
短命植物经长期进化形成了独特的生态化学计量特征,该研究以古尔班通古特沙漠南缘主要短命植物为研究对象,通过对研究区的66个样方、109个植物样品、42个物种的叶片生源要素的化学计量特征进行分析,以明确研究区的主要短命植物在个体尺度、种群尺度、群落尺度上的化学计量特征及异速关系,揭示制约古尔班通古特沙漠南缘短命植物生长的主要限制元素。结果显示:(1)在3个尺度上,短命植物叶片K含量均高于全国平均水平,而N、P含量均低于全国平均水平,且N∶P均值都小于14;Ca含量在个体尺度和种群尺度上低于全国平均水平,而群落尺度上高于全国平均水平。(2)个体尺度上,各生源要素间的相关性以Ca-Mg最高、K-Mg其次、Ca-S最低;各元素增长斜率显著偏离1,各元素间呈异速增长关系,增长速率为 Mg>Ca>K>P>Fe>N>S。(3)种群尺度上,Ca-Mg的增长指数为0.82(接近1),说明两元素间大致呈等速增长,各元素间的增长速率为Ca>K>P>S>N>Fe,其中Ca的增长速率最大,它在植物生物量大量积累时期,植物会获得更多的Ca为开花繁殖做准备。(4)群落尺度上,各元素间相关性的拟合优度都高于个体尺度,各生源要素间具有显著的异速增长关系,增长速率为K>Ca>Mg>S>P>N>Fe,表明短命植物对不同元素的利用速率不同。研究发现,古尔班通古特沙漠南缘短命植物在受N、P限制的同时更易受N限制,群落尺度上具有较高一致性生物学特征和相同生态适应性的物种具有更高的拟合优度。
短命植物;生源要素;化学计量;异速增长
生态化学计量学是研究生态系统能量和化学元素平衡的科学,强调的是有机体主要组成元素间的耦合关系[1]。生源要素是生命体所必须的化学元素,如氮(N)、磷(P)、钾(K)、铁(Fe)、钙(Ca)、硫(S)、镁(Mg)等化学元素是植物生命体的必要组成部分,这些元素的耦合关系不但影响着植物的生理代谢过程,而且通过生物地球化学循环作用于生态系统功能[2],如土壤微生物中N、P、K间的耦合关系通过生物地球化学循环,使得N、P、K含量在土壤中趋于较稳定的范围[3]。植物叶片的生态化学计量特征对所在环境整体养分条件具有一定的指示作用,如He等[4]研究显示植物叶片中N、P含量可表征所在环境的N、P养分条件;Campo等[5]研究揭示了陆地植物叶片 N、P 含量能反映土壤 N、P 的有效性;Koerselman等[6]研究得出植物叶片中N∶P低于14时受N限制,而高于16时受P限制,介于14~16之间则受到N、P的双重限制。因此,探究植物叶片元素间的相互关系,有助于预测生态系统对环境变化的响应,以及元素在植物生产力中的限制作用[7]。
此外,Fe、Ca、K、Mg、S等生源要素在植物生命活动中发挥着极其关键的作用[8],如Fe影响着植物叶绿素合成和众多酶活性,缺Fe会造成叶绿素合成受阻,而引起黄化病的发生;Ca能调节植物的渗透和酶活作用,在植物的选择性吸收和抗逆性方面起着关键调节作用[9];K会提升叶片的保水能力及促进气孔导度的增加,并提高叶片胞间CO2浓度含量和叶绿素含量,以及增加净光合速率获得更多的生物量[10];S对一些必需氨基酸、蛋白质、多糖、维生素和辅酶的合成起重要作用[11]。目前,化学计量特征研究大多集中在C∶N∶P及其驱动因素等方面,其中针对植物叶片N、P含量及N∶P的研究成果居多,而对植物叶片中其他生源要素含量与N、P元素间相互关系的研究相对较少。
短命植物是在短时间(3~6月)迅速完成其生活史的一类特殊植物类型,广泛分布于中国新疆北部的准噶尔盆地(约205种),作为荒漠生态系统的重要组成部分,在维持物种多样性和荒漠生态系统净初级生产力、碳循环、能量流动等方面[12]发挥着极其重要的作用。因此,本试验以古尔班通古特沙漠南缘主要短命植物为研究对象,通过对研究区的109个植物样品、42个物种、66个样方进行分析,试图从植物个体尺度、种群尺度和群落尺度探索生源要素的化学计量特征及异速关系,回答如下科学问题:(1)结合不同尺度(个体尺度、种群尺度、群落尺度)的化学计量特征,探明制约古尔班通古特沙漠南缘短命植物生长的主要限制元素是什么?(2)通过对不同尺度之间异速关系的研究,阐明元素之间的相关性和增长关系是如何表现的?对上述问题的探索将补充荒漠生态系统化学计量特征的研究资料,以期为今后开展相关研究提供基础数据和科学参考。
1 研究区概况与研究方法
1.1 研究区概况
古尔班通古特沙漠南缘属中温带大陆性干旱气候,冬季漫长寒冷,夏季炎热高温,年蒸发量大,全年平均气温为6.6 ℃,大于10 ℃年积温为3 000~5 000 ℃。年均无霜期为140 d左右, 年降水量为100~150 mm,且以冬季降雪为主,夏、秋季以降水和凝结水为植物主要水源[13]。该区属于中国最大固定、半固定沙漠,土壤被固定在沙丘之间的低洼地以及沙丘中下部。作为荒漠生态系统重要组成部分的短命植物广泛分布于该地区,主要有尖喙牻牛儿苗(Erodiumoxyrrhynchum)、沙蓬(Agriophyllumsquarrosum)、条叶庭芥(Alyssumlinifolium)、齿稃草(Schismusarabicus)、小花糖芥(Erysimumcheiranthoides)和粗柄独尾草(Eremurusinderiensis)等。
1.2 研究方法
1.2.1 样品采集 短命植物和类短命植物的旺盛生长期集中在每年5月份,因此,本试验于2014年5月中旬进行连续取样。样区全年降雨量小于150 mm,且主要集中在冬季,年蒸发量在2 000 mm以上,5~8月份空气相对湿度小于45%[13]。选择生境较均质的丘间平地设置样地,共设置30个典型样地(10 m×10 m),每个样地间隔不超过500 m,根据每个样地内植物的生长状况和分布状况,再设置1~5个样方(1 m×1 m),收割每个样方内所有草本植物的地上部分,按照物种分类后称量鲜重,采样时重点采集样方中比较常见的优势种,并兼顾稀少种,共采集42物种的109个植物样品。将分类好的样品装入保鲜袋中,并装入擦拭干净且带冰块的泡沫箱内迅速带回实验室。用自来水洗净灰尘和杂质,再用无离子水冲洗至少3次,置于80 ℃的烘箱烘干至恒重后取出按物种称重。将每份烘干的样品置于球磨仪中研磨成粉状,然后测定N、P、K、Mg、Fe、Ca、S元素含量。全氮含量利用凯氏定氮法测定 (GB 5009.5-2010),P含量用钼锑抗比色法测定,其余元素含量用PS-4电感耦合等离子发射光谱仪测定(Baird & Associates Ltd.,Madison,USA)。
1.2.2 数据分析 本文从个体、种群和群落3个尺度上考虑不同元素之间的相关关系,分析种群尺度关系时,将同一物种的所有个体数据平均后再进行计算。而分析群落尺度上各元素间的关系时,样方中每种植物叶片的元素含量是将其生物量占样方总生物量的比例加权后得到群落尺度的平均元素含量。即:EP=∑ES×BS,其中EP为群落尺度生源要素的平均含量,ES为个体叶片生源要素的含量,BS为各物种在样方中生物量的比例[14]。
对3个尺度(个体尺度、种群尺度、群落尺度)的数据先求算术平均值,然后对数据进行对数转换,使之服从正态分布以进行数量分析。采用y=axb来讨论不同生源要素间的关系, 线性转换为lgy= lga+blgx,其中,x代表一种生源要素含量,y代表另一种生源要素含量,a代表对数化后两生源要素间关系的截距,b表示两者对数化后的斜率,b也称为相对增长指数(当b=1时为等速增长,当b大于或小于1时为异速增长)。
文中采用多元线性回归法获得各元素之间的相关性及各元素之间的斜率(增长关系), 数据分析与作图均在R-3.2.2软件中完成。
2 结果与分析
2.1 3种尺度生源要素化学计量特征
表1结果显示,研究区短命植物叶片各元素含量在3种尺度上均呈现出不同的变化趋势,其中,N、P含量在3种尺度上都低于全国平均水平(N全国均值20.5 mg/g;P全国均值1.4 mg/g),K含量在3种尺度均高于全国平均水平(K全国均值10.3 mg/g),Ca含量在个体尺度和种群尺度上均低于全国平均水平,而群落尺度上高于全国平均水平(Ca全国8.81 mg/g)[15],其他元素含量与全国平均水平相差不大。另外,研究区短命植物叶片N∶P平均值在个体尺度上是5.03(变化范围介于0.8~16),在种群尺度上为4.36(变化范围介于1.4~13),而群落尺度上是3.7(变化范围介于0.1~7.8)。以上结果说明,在3种尺度上,研究区短命植物均受到较强的N、P限制,而K几乎不受限制。3种尺度上各元素含量的差异可能是在不同尺度上植物具有不同的养分利用策略而引起的。
2.2 个体尺度各生源要素间的相关关系及相对增长指数
个体尺度上元素之间的相关系数为,Ca-Mg最高,K-Mg其次,Ca-S最低(图1)。表2显示,Mg与Ca、K、Fe 3种元素都具有显著相关性(P<0.01),K与Mg、P、S 3种元素间都存在极显著的相关性(P<0.001)。
在个体尺度上,Ca元素相对于Mg元素的增长速率最大(为0.48)但显著偏离1,表明两者呈异速增长关系,S元素相对于Mg元素的增长速率最小(为-0.02)。S元素相对于Mg和P元素的增长速率都为负值,分别为-0.02和-0.01(表2)。各元素的增长速率如下:Mg>Ca>K>P>Fe>N>S。

表1 3种尺度下各元素的平均值及变化范围

表2 个体尺度和群落尺度上各元素的增长指数与相关系数
注:下三角矩阵代表植物个体尺度(列代表y,行代表x),上三角矩阵代表植物群落尺度(列代表x,行代表y) 分隔号前代表增长斜率,分隔号后为相关系数(R)。***代表极显著(P<0.001);**代表显著(P<0.01);*代表具有显著性(P<0.05)
Note:The lower triangular matrix represents the ephemeral plant individual scale. (columns representativey, rows representativex), and the upper triangular matrix represents the ephemeral plant community scale (column representativex, rows representativey). The number of before the separator representatives the growth rate, the number after the separator is correlation coefficient (R).*** represent extreme significant(P<0.001), ** represent more significant(P<0.01), * represents significant(P<0.05)

上三角矩阵中数字代表决定系数(R2),下三角矩阵中曲线代表各元素之间的拟合优度;下同图1 个体尺度上各生源要素间的关系及拟合曲线The number represents the determination coefficient in the upper triangular matrix,and the curve represents the goodness of fit between the elements in the lower triangular matrix; The same as belowFig.1 The relationship and fitting curves of various biogenic elements in individual scale
2.3 种群尺度各生源要素间的相关关系及相对增长指数
表3结果显示,各元素间相关系数较高的为Mg-Ca(0.68),Mg-P(0.4),Mg-K(0.39),而相关系数最低的是Mg-N(0.006 4)。结合图2和表3结果可知,在种群尺度上Ca和Mg两元素具有显著的相关性,且两者的异速增长指数为0.82,大致接近于1,说明两元素间大致呈等速增长关系,其中Ca元素相对于其他元素的异速增长斜率都为正值,Fe元素相对于其他元素的异速增长斜率都为负值。各元素间的增长速率为Ca>K>P>S>N>Fe。
2.4 群落尺度各生源要素间的相关关系及相对增长指数
群落尺度上各元素间的相关性都很高,最高为1,最低也达到了0.79,群落尺度上各元素间相关性的拟合优度都高于个体尺度(图3)。从表2可知Mg与K、Ca、S、Fe 元素;S与Ca、Fe元素;Fe与N元素都具有极显著的相关性(P<0.001)。
表2结果显示,在群落尺度上K相对于Mg、S,Ca相对于Mg、S、K的增长速率都与1极为接近,说明两元素之间呈等速增长关系。而N-P的增长速率为0.74与3/4极为接近,表明N-P元素间呈异速增长趋势,而增长速率最小的Fe-P为0.66与2/3极为接近,在群落尺度上各元素的增长速率为K>Ca>Mg>S>P>N>Fe。
3 结论与讨论
本研究显示,古尔班通古特沙漠南缘短命植物叶片的N、P含量在3种尺度上都低于全国平均水平[15],但K含量在3种尺度均高于全国平均水平;Ca含量在个体尺度和种群尺度均低于全国平均水平,而在群落尺度高于全国平均水平[15],其他元素含量与全国平均水平相差不大。

表3 种群尺度上各元素的增长指数与相关系数
注:下三角矩阵代表增长指数,上三角矩阵代表相关系数(R),***代表极显著(P<0.001);**代表显著(P<0.01);*代表具有显著性(P<0.05)
Note:The lower triangular matrix represents growth index, the upper triangular matrix represents correlation coefficient(R), *** represent extreme significant(P<0.001), ** represent more significant(P<0.01), * represents significant(P<0.05)
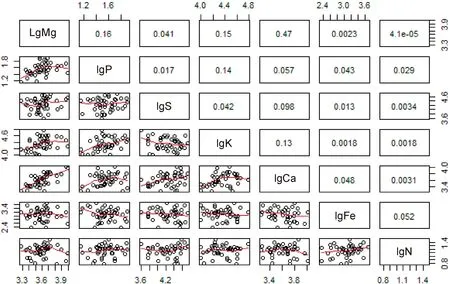
图2 种群尺度上各生源要素间的关系及拟合曲线Fig.2 The relationships and fitting curves of various biogenic elements in population scale
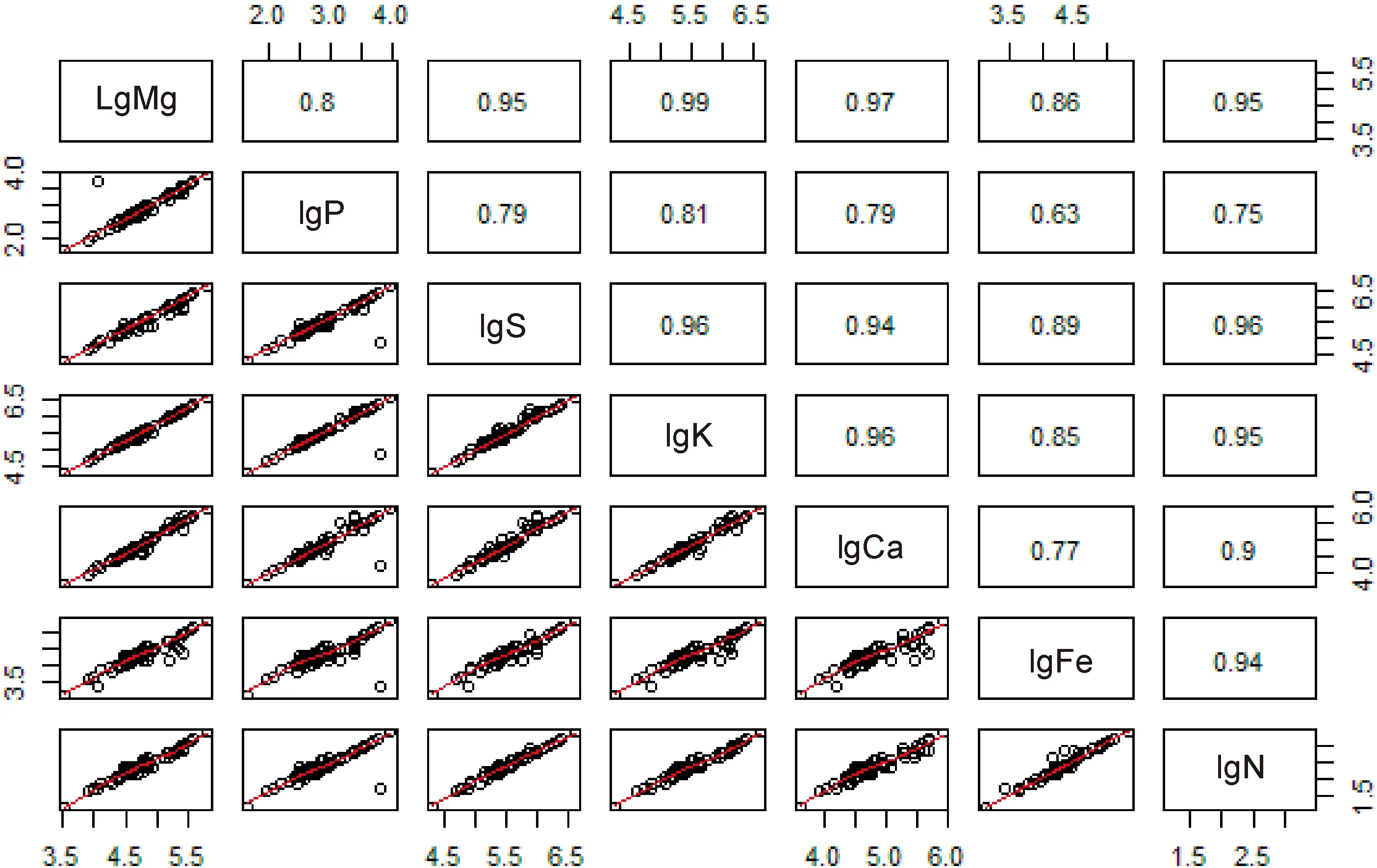
图3 群落尺度上各生源要素间的关系及拟合曲线Fig.3 The relationships and fitting curves of various biogenic elements in community scale
3.1 3种尺度生源要素化学计量特征及异速增长关系
本研究中个体尺度上N、P含量远低于全国平均水平, 表明短命植物生长受严重的N、P限制,其中N∶P平均值5.039,进一步揭示了受N、P限制的同时更易受N限制。各元素增长斜率显著偏离1,说明各元素间呈异速增长关系,其中Ca增长速率大于K、P、Fe、N、S等元素,表明荒漠短命植物在主要生长阶段对不同养分的吸收速率有所差异,相关研究表明,Ca在调节植物的渗透作用和酶活作用,以及对植物的选择性吸收和抗逆性等方面都有重要作用[9]。Ca的增长速率大于其他元素,进一步阐明了Ca在调节荒漠短命植物渗透作用和酶活作用及抗逆性等方面发挥着重要作用。
植物物种间的差异,会造成叶片化学计量特征的不同,本研究中种群尺度上N、P平均含量分别为1.51和0.39 mg/g,显著低于阎恩荣等[16]对浙江天童常绿阔叶林C∶N∶P化学计量特征研究、宋彦涛等[17]对松嫩草地 80种草本植物叶片N、P化学计量特征研究、李玉霖等[18]对北方典型荒漠及荒漠化地区植物叶片N、P化学计量特征研究,也显著低于Han等[19]对中国753种陆生植物叶片 N、P 的研究,表明不同物种之间N、P化学计量特征具有较大的差异。有研究表明Ca在植物的花芽分化、成花诱导中起着重要作用[20],本研究中Ca与Mg成等速增长且Ca的增长速率最大,表明在短命植物生物量大量积累时期,植物会获得更多的Ca为开花繁殖做准备。
群落尺度上,各生源要素的含量都高于个体尺度,表明不同尺度元素的含量具有较大差异。Ca含量在个体尺度和种群尺度上均低于全国平均水平,而在群落尺度上高于全国平均水平,这可能是由于Ca在不同尺度上对调节植物的渗透作用和酶促作用以及抗逆性等方面有所差异引起的,说明了在不同尺度上植物具有不同的养分利用策略。同时本研究显示各生源要素间具有显著的异速增长关系,表明短命植物对不同元素的利用速率不同。Reich等[21]研究表明N-P的异速增长指数为2/3,本研究N-P间的异速增长指数在群落尺度上为0.74与2/3接近。群落尺度上N素的增长速率大于个体尺度,揭示了N在短命植物群落构建以及生长过程中具有重要意义;Fe的增长速率小于K、Ca、Mg、S、P、N等元素,表明Fe在短命植物生长中增长速率相对较小。事实上,有研究表明Fe在海洋浮游生物生长过程中起到了极为重要的作用[22],但对Fe在陆地生态系统中是否具有极为重要的作用,还缺乏系统的研究。但在个体尺度上Fe元素的增长速率大于N 和 S两种元素,这可能是Fe与其他元素间的化学计量关系与植物群落相互作用具有很大的联系。
3.2 3种尺度化学计量特征的共性
在3种尺度上,本研究中荒漠短命植物叶片N、P含量均低于全国平均水平。有研究表明,植物叶片的N、P含量在各生长阶段具有差异[23];牛得草等[24]在旱生草本植物,吴统贵等[25]在湿生草本植物研究中得出叶片N、P含量都会随植物生物量积累而降低。由于本研究样品采于5月份,而这期间恰是荒漠短命植物生物量最大积累阶段,因此,这可能是造成研究区短命植物叶片N、P含量较低的主要原因。生长速率假说认为生命体的生长受到蛋白质与核酸的双重限制[26],而这两种物质都富含N、P元素,表明古尔班通古特沙漠南缘短命植物叶片生长受到严重的N、P限制。
K元素含量在3种尺度均高于全国平均水平(K含量10.3 mg/g),这与Rivas-Ubach等[27]认为K在干旱胁迫中的重要地位一致。古尔班通古特沙漠南缘植物生长受到严重的干热胁迫,K元素增加会提高叶片保水能力及保卫细胞的溶质浓度,促进气孔导度增加[10]使叶片气孔阻力减小,提高了叶片胞间 CO2浓度含量和叶绿素含量,从而增加净光合速率获得更多的生物量,这可能是造成K在植物体内大量集聚的重要原因。另外在群落尺度上K的增长速率最大,进一步说明了K在该地区的植物生理代谢中具有更加重要的作用。
3种尺度上,荒漠短命植物叶片N∶P均小于14,陆生植物体相对稳定的N、P含量以及N∶P是植物生存的关键适应机制[28],植物叶片中N∶P化学计量特征是确定植物受N或P限制的重要指示剂,是植物适应环境的结果,Koerselman等[6]研究得出植物叶片中N∶P低于14时受N限制,而高于16时受P限制,介于14~16之间则受到N、P的双重限制,本研究中N∶P显著低于闫邦国等[14]对金沙江干热河谷地区植物叶片生源要素的研究,年降水量小于200 mm的古尔班通古特沙漠南缘相对较低的N含量可能导致了较低的N∶P,3种尺度上N∶P均低于14,进一步阐明了此地植物生长受N、P限制的同时更易受N素限制,这与N素被认为是古尔班通古特沙漠植物生长的主要限制元素的结论相符合。此外,本研究发现K∶N、K∶P都高于全国平均水平,原因主要为N、P含量均低于全国平均值且K含量高于全国均值,从而使得K∶N、K∶P都高于全国平均水平。
关于古尔班通古特沙漠南缘植物生长主要受到N限制的原因可能是:(1)植物的营养含量在一定程度上反映了植物生长的生境条件[29],该研究区土壤养分总氮含量变化范围在0.18-0.22 mg/g[30],与其他生态系统相比该区土壤养分显得更加贫瘠,因此,可能会导致植物对N素的利用和吸收减少,从而导致植物体内N素含量明显减少;(2)植物生长所需的90%以上的N素都来自微生物对植物凋落物的分解过程所释放的N素[31],干旱的环境条件和相对较低的微生物分解速度会导致养分的可利用性比其他地区显著偏低,这会进一步影响植物对N素的吸收[32-33]。
3.3 群落尺度与个体尺度异速关系的异性
首先,群落尺度上各元素间的关系与个体尺度间的关系差异较大。这可能与植物在群落尺度上与其他植物之间在空间养分吸收上具有相互作用与相互影响有关,也可能与群落中不同植物在组织器官上的养分分配不同有关。由于本研究主要采集的是植物叶片,因此很有可能是物种之间的相互作用引起的。
其次,群落尺度上元素间的增长速率相对于个体尺度更加趋于稳定,资源比例假说[34]表明,物种在不同资源上具有不同的利用效率或竞争力时物种间更容易相互共存,因此,在群落尺度上物种间的相互竞争以及对不同资源具有不同利用效率,使得它们之间更容易共存,另外,一起共存的物种通过调整生物量和主要生源要素含量,降低了个体化学计量特征引起的差异,使得各元素间在群落尺度上比在个体尺度上更加趋于一个稳定的斜率。
另外,群落尺度上各元素间相关性的拟合优度都高于个体尺度,张金屯和焦蓉[35]认为,在同一生态种组的种,对资源的利用力和环境的适应性以及在群落中所起的作用都具有一致性,因此会呈现明显的相关性。Greig-Smith[35]认为,种间具有一致的生物学特性和生态适应性时,他们之间的相关性趋于稳定,同时他将样方大小对相关性的影响进行了详细的分析,认为如果某种植物的个体大于其他种,该种就会对其他种在空间上产生排斥,从而使整体间呈现出稳定的相关性。本研究对象均为具有一致的生物学特性和生态适应性的短命植物,且在采样时选择的样地生境均一,样方大小适中,每个样方中都有优势种存在,这可能是导致群落尺度上各元素间相关性的拟合优度都高于个体尺度上的主要原因。
植物叶片中生源要素的增长关系与土壤间的化学循环,以及生态系统化学元素的整体循环都具有密切的联系,同时也与植物的生理代谢和环境适应策略有关[36],植物体内的化学计量元素受到多种尺度,如纬度、高温、降水、岩石分化、大气N沉降、群落演替、微生物作用等影响[23],这种不同尺度对生物地球化学计量特征的影响需要更加深入的研究去揭示,此外多数研究对N、P外的其他生源要素关注较少,因此研究N、P外其他生源要素与N、P之间的交互作用将是今后研究的重点。
综上所述,古尔班通古特沙漠南缘荒漠短命植物在受到N、P双重限制的同时更易受到N限制,说明N素是该荒漠区植物生长的主要限制性元素;研究发现群落尺度上具有较高一致的生物学特征和相同生态适应性的物种具有更高的拟合优度;物种在群落尺度上对生源要素具有不同的利用效率或竞争力时,会降低个体化学计量特征引起的差异而易促进物种间相互共存,从而使各生源要素间在群落尺度上趋于较稳定的增长速率。
[1] 杨惠敏,王冬梅. 草-环境系统植物碳氮磷生态化学计量学及其对环境因子的响应研究进展[J]. 草业学报,2011,20(2):244-252.
YANG H M,WANG D M. Advances in the study on ecological stoichiometry in grass-environment system and its response to environmental factors[J].ActaPrataculturaeSinica,2011,20(2):244-252.
[2] AERTS R,CHARPIN FS III.The mineral nutrition of wild plants revisited:A re-evaluation of processes and patterns[J].AdvancesinEcologicalResearch,2000,30(8):1-67
[3] CLEVELAND C C,LIPTZIN D. C:N:P stoichiometry in soil:Is there a“Redfield ratio”for the microbial biomass? [J].Biogeochemistry,2007,85(3):235-252.
[4] HE J S,WAN L,FLYNN D F B,etal. Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes[J].Oecologia,2008,155(2):301-310.
[5] CAMPO J,VZQUEZ YANES C.Effects of nutrient limitation on aboveground carbon dynamics during tropical dry forest regeneration in Yucatn,Mexico[J].Ecosystems,2004,7(3):311-319.
[6] KOERSELMAN W,MEULEMAN A F M. The vegetation N:P ratio:a new tool to detect the nature of nutrient limitation[J].JournalofAppliedEcology,1996,33(6):1 441-1 450.
[7] KERKHOFF A J,ENQUIST B J. Ecosystem allometry:the scaling of nutrient stocks and primary productivity across plant communities[J].EcologyLetters,2006,9(4):419-427.
[8] GLEESON S K,GOOD R E. Root allocation and multiple nutrient limitation in the New Jersey pinelands[J].EcologyLetters,2003,6(3):220-227.
[9] AMATO I. The secret life of plant crystals[J].Science&Technology. 2006,84(6):26-28
[10] 刘建祥,杨肖娥,吴良欢,等.低钾胁迫对水稻叶片光合功能的影响及其基因型差异[J].作物学报.2001,27(6):1 000-1 006.
LIU J X,YANG X E,WU L H,etal. The effects of low potassium stress on the photosynthetic function of rice leaves and their genotypic difference[J].ActaAgronomicaSinica,2001,27(6):1 000-1 006.
[11] 谭丽菊,肖 慧,CRAIG A. CARLSON,等. 海水中天然细菌对不同生源要素有机物的矿化作用[J]. 生态学报,2016,36(1):77-85.
TAN L J,XIAO H,CRAIG A. CARLSON,etal. Mineralization of different dissolved organic matter containing various biogenic elements by natural bacteria in seawater[J].ActaEcologicaSinica,2016,36(1):77-85.
[12] GILLIAM F S. The ecological significance of the herbaceous layer in temperate forest ecosystems[J].BioScience,2007,57(10):845-858.
[13] 毛祖美,张佃民. 新疆北部早春短命植物区系纲要[J]. 干旱区研究,1994,11(3):1-26.
MAO Z M,ZHANG D M . The Conspectus of Ephemeral Flora in Northern Xinjiang[J].AridZoneResearch,1994,11(3):1-26.
[14] 闫帮国,何光熊,李纪潮,等. 金沙江干热河谷地区植物叶片中各生源要素的化学计量特征以及异速增长关系[J]. 植物生态学报,2012,36(11):1 136-1 144.
YAN B G,HE G X,LI J C,etal. Scaling relationships and stoichiometry of plant leaf biogenic elements from the arid-hot valley of Jinsha River,China[J].ChineseJournalofPlantEcology,2012,36(11):1 136-1 144.
[15] HAN W X,FANG J Y,REICH P B,etal. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate,soil and plant functional type in China[J].EcologyLetters,2011,14(8):788-796.
[16] 阎恩荣,王希华,郭 明,等. 浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C∶N∶P化学计量特征[J]. 植物生态学报,2010,34(1):48-57.
YAN E R,WANG X H,GUO M,etal. C∶N∶P stoichiometry across evergreen broad-leaved forests,evergreen coniferous forests and deciduous broad-leaved forests in the Tiantong region,Zhejiang Province,eastern China[J].ChineseJournalofPlantEcology,2010,34(1):48-57.
[17] 宋彦涛,周道玮,李 强,等.松嫩草地 80 种草本植物叶片氮磷化学计量特征[J].植物生态学报,2012,36(3):222-230.
SONG Y T,ZHOU D W,LI Q,etal. Leaf nitrogen and phosphorus stoichiometry in 80 herbaceous plant species of Songnen grassland in Northeast China[J].ChineseJournalofPlantEcology,2012,36(3):222-230.
[18] 李玉霖,毛 伟,赵学勇,等.北方典型荒漠及荒漠化地区植物叶片氮磷化学计量特征研究[J].环境科学,2010,31(8):1 716-1 725.
LI Y L,MAO W,ZHAO X Y,etal. Leaf nitrogen and phosphorus stoichiometry in typical desert and desertified regions,North China[J].EnvironmentalScience,2010,31(8):1 716-1 725.
[19] HAN W X,FANG J Y,GUO D L,etal.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J].NewPhytologist,2005,168(2):377-385.
[20] 孔海燕,贾桂霞,温跃戈. 钙在植物花发育过程中的作用[J]. 植物学通报,2003,20(2):168-177.
KONG H Y,JIA G X,WEN Y G. The role of calcium in flower development[J].ChineseBulletinofBotany,2003,20(2):168-177.
[21] REICH P B,OLEKSYN J,WRIGHT I J,etal. Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes[J].ProceedingsoftheRoyalSocietyB:BiologicalSciences,2010,277(1 683):877-883.
[22] TSUDA A,TAKEDA S,SAITO H,etal. A mesoscale iron enrichment in the western subarctic Pacific induces a large centric diatom bloom[J].Science,2003,300(5621):958-961.
[23] STERNER R W,ELSER J J,VITOUSEK P. Ecological Stoichiometry:the Biology of Elements from Molecules to the Biosphere[M]. Princeton: Princeton University Press. 2002.
[24] 牛得草,李 茜,江世高,等. 阿拉善荒漠区6种主要灌木植物叶片C∶N∶P化学计量比的季节变化[J]. 植物生态学报,2013,37(4):317-325.
NIU D C,LI Q,JIANG S G,etal. Seasonal variations of leaf C∶N∶P stoichiometry of six shrubs in desert of China’s Alxa Plateau[J].ChineseJournalofPlantEcology, 2013,37(4):317-325.
[25] 吴统贵,吴 明,刘 丽,等. 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化[J]. 植物生态学报,2010,34(1):23-28.
WU T G,WU M,LIU L,etal. Seasonal variations of leaf nitrogen and phosphorus stoichiometry of three herbaceous species in Hangzhou Bay coastal wetlands,China[J].ChineseJournalofPlantEcology,2010,34(1):23-28.
[26] ELSER J J,ACQUISTI C,KUMAR S. Stoichiogenomics:the evolutionary ecology of macromolecular elemental composition[J].TrendsinEcologyandEvolution,2011,26(1):38-44.
[27] RIVAS-UBACH A,SARDANS J,PÉREZ-TRUJILLO M,etal. Strong relationship between elemental stoichiometry and metabolome in plants[J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2012,109(11):4 181-4 186.
[28] TAO Y,ZHANG Y M. Seasonal changes in species composition, richness and the aboveground biomass of three community types in Gurbantünggüt Desert,Northwestern China[J].ActaPrataculturaeSinica,2011,20(6):1-11.
[29] HUANG J J,WANG X H.Leaf nutrient and structural characteristics of 32 evergreen broad leaved species[J].JournalofEastChinaNormalUniversity:NaturalScience,2003,(1):92-97.
[30] 陶 冶,张元明.古尔班通古特沙漠4种草本植物叶片与土壤化学计量特征[J].应用生态学报,2015,26(3):659-665.
TAO Y,ZHANG Y M. Leaf and soil stoichiometry of four herbs in the Gurbantunggut Desert,China[J].ChineseJournalofAppliedEcology,2015,26(3):659-665.
[31] CRAINE J M,JACKSON R D.Plant nitrogen and phosphorus limitation in 98 North American grassland soils[J].PlantandSoil,2010,334(1-2):73-84.
[32] DIJKSTRA F A,PENDALL E,MORGAN J A,etal.Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland[J].NewPhytologist,2012,196(3):807-815.
[33] FERNANDZ D P,NEFF J C,BELNAP J,etal.Soil respiration in the cold desert environment of the Colorado Plateau (USA) :abiotic regulators and thresholds[J].Biogeochemistry,2006,78(3):247-265.
[34] TILMAN D. Resource Competition and Community Structure[M]. Princeton University Press,Princeton. 1982.
[35] 张金屯.数量生态学[M].第二版.北京:科学出版社,2011: 108-109.
[36] ÅGREN G I. Stoichiometry and nutrition of plant growth in natural communities[J].AnnualReviewofEcology,EvolutionandSystematics,2008,39:153-170.
(编辑:裴阿卫)
The Stoichiometry and Allometric Growth Relationship of Ephemeral Plant Leaf Biogenic Elements from the Southern Gurbantungut Desert,China
LIU Jianguo1,2,LIN Zhe1,2,LIU Weiguo1,2*,HUO Jusong1,2
(1 College of Resources and Environment Science,Xinjiang University,Urumqi 830046,China;2 Xinjiang Key Laboratory of Oasis Ecology,Urumqi 830046,China)
The ephemeral plants formed their own unique ecological stoichiometry characteristics under the extreme habitat conditions. Taking the ephemeral plants in the southern Gurbantunggut Desert as the research object,this thesis aims at analyzing the stoichiometric characteristics of leaf biogenic elements of 66 plots,109 plant samples and 42 species in the study area to clarify the stoichiometric characteristics and allometric relationships of the main ephemeral plants in the individual scale,population scale and the community scale,and reveals the main limiting elements which restrict the growth of ephemeral plants in southern Gurbantunggut Desert. The results were as follows:(1) on the 3 scales,the K content of the ephemeral plant leaves was higher than that of the national average,while the content of P and N was lower than that of the national average level and N∶P mean values were less than 14. The Ca contents of individual scale and population scale is lower than that of the national average,while the Ca content is higher than that of the national average level under community scale. (2) The correlation among the elements of the individual scale is Ca-Mg highest,K-Mg second,and Ca-S minimum,and the growth rate of each element is significantly deviated from 1,which showed the allometric growth of the various elements and the growth rate of Mg > Ca > K > P > Fe > N > S. (3) The rate of Ca-Mg in the population scale was 0.82 (close to 1),which showed that the two elements are roughly constant growth. The growth rate of each element is Ca>K>P>S>N>Fe. In addition,the Ca was the largest,which indicated that plants could get more Ca to prepare for flowering and reproduction during the period of plant biomass large accumulation. (4) The goodness of fit of each element on the community scale is higher than that of the individual scale and each elements have significant allometric growth realationship. The growth rate is K>Ca>Mg>S>P>N>Fe,which showed that the utilization rate of distinct elements is not same. The result found that the southern Gurbantunggut Desert ephemeral plants were restricted by N and P but more susceptible to N limitation. It also verified that the species with high consistent biological characteristics and the same ecological adaptability will have higher goodness of fit in the community scale.
ephemeral plants;biogenic elements; stoichiometric characteristics;allometric growth
1000-4025(2016)11-2291-09
10.7606/j.issn.1000-4025.2016.11.2291
2016-06-27;修改稿收到日期:2016-10-18
国家自然科学基金(31260112)
刘建国(1990-),男,在读硕士研究生,主要研究方向为全球变化与生态系统生态学,E-mail:2515517759@qq.com
*通信作者:刘卫国,博士,副教授,硕士生导师,主要从事干旱、半干旱区荒漠植物研究。E-mail:wgliuxj@126.com
Q946.91;Q948.113
A
