青钱柳光合作用日变化和光合响应特征
2017-01-03柏明娥方建华徐高福沈建军陈秀娟
柏明娥,方建华,徐高福,沈建军,陈秀娟
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江省淳安县千岛湖林场,浙江 淳安 311700;3. 浙江省淳安县新安江开发总公司,浙江 淳安 311700)
青钱柳光合作用日变化和光合响应特征
柏明娥1,方建华2,徐高福3,沈建军1,陈秀娟1
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 浙江省淳安县千岛湖林场,浙江 淳安 311700;3. 浙江省淳安县新安江开发总公司,浙江 淳安 311700)
以一年生青钱柳(Cyclocarya paliurus)播种苗为试材,对其叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等光合生理指标的日变化及光响应曲线进行测定,分析了光合参数间的相关性。结果表明:青钱柳苗Pn、Tr、Gs的日变化均为“双峰”曲线,两个峰值分别出现在9∶30和13∶00,Ci的日变化表现为早晚高、中午低,低谷出现在11∶30;Pn与Gs呈极显著相关(P < 0.01),与Tr呈显著相关(P < 0.05),与其它参数间的相关性不显著;光补偿点为14.63 µmol/(m2·s),光饱和点为1 200 µmol/(m2·s),最大净光合速率为8.32 µmol/(m2·s);影响青钱柳光合生理参数的主要环境因子为光合有效辐射和大气温度(Ta)。
青钱柳;光合作用;日变化;光合响应
青钱柳(Cyclocarya paliurus)系胡桃科(Juglandaceae)青钱柳属(Cyclocarya)落叶高大乔木,广泛分布于我国亚热带地区省(市),是我国特有的单种属植物[1]。青钱柳叶中含有多种对人体有益的活性有机成分如黄酮、多糖、三萜、有机酸等和钙、镁、锰、铜、铬、锌、硒、矾、锗等多种无机元素[2~4],因而具有清热解毒、抗氧化、抗肿瘤、降低血糖、血脂和血压等多种药理功能[5~8],被认为是一种具有较高开发利用价值的珍稀树种。目前对青钱柳的研究主要集中在生物学特性与资源分布、种苗繁殖、化学成分、药理作用、保健功能、有效成分提取及产品研发等方面[9~12],而对其生理生态方面的研究尚少。
光合作用是植物重要的生理活动,其强度大小受植物类型、光照、温度、CO2浓度、水、矿质元素等内外因素的影响[13~14]。已有学者对青钱柳的光合生理特征[15]、不同种源间的光合特性差异[16]等方面进行了研究,表明自然条件下生长的青钱柳光合速率日进程呈双峰曲线,不同种源间青钱柳的光合作用特征差异显著。本文以1年生青钱柳播种苗为试材,研究分析了青钱柳苗期光合生理参数的日变化规律、净光合速率(Pn)对光照强度的光合响应特征、Pn与光合有效辐射(PAR)、大气温度(Ta)、气孔导度(Gs)、蒸腾速率(Tr)等光合参数间的相关性,以期了解和掌握青钱柳对光环境的生态适应性及适应机制,为青钱柳资源的高效培育和栽培管理提供技术支撑。
1 材料与方法
1.1 试验材料
试验种源来自浙江省安吉县天荒坪镇大溪村,119° 35′ 25.9″ E,30° 27′ 13.1″ N,海拔 677 m,采种母树树龄300 ~ 500 a,胸径62 cm,树高25 m。2014年10月采集种子,种子经浓硫酸酸蚀和赤霉素浸种处理后进行室外湿砂贮藏[17],2015年3月取出进行条播。播种地位于浙江省林业科学研究院苗圃地内,120° 1′ 29″ E,30° 13′12″ N,海拔32 m,该地属亚热带湿润季风区,四季分明,雨量充沛,年平均降水量为1 400 mm,年平均气温16℃左右,圃地土壤类型为山地红壤。
1.2 试验方法
1.2.1 光合作用日变化的测定 随机选取长势相近的青钱柳植株3株,于2015年9月初较晴朗天气,采用LI-6400便携式光合作用测定系统测定叶片各项光合生理参数的日变化,所测叶片位于植株上部的第5至第6片叶,每片叶中选取靠顶端的3张小叶各测1次,共测3次,结果取平均值。
测定参数为Pn(µmol /(m2·s))、Gs(mol/(m2·s))、Tr(mmol/(m2·s))、胞间CO2浓度(Ci,µmol/mol),同时记录PAR(µmol/(m2·s))、Ta(℃)、叶片温度(Tl,℃)、大气CO2浓度(Ca,µmol/mol)、空气相对湿度(RH,%)等。气孔限制值(Ls)和水分利用效率 (WUE,µmol/mmol)根据以下公式计算:Ls=( Ca- Ci)/ Ca,WUE=Pn/ Tr。测定时间从7:30到17:00。
1.2.2 光响应曲线测定 利用LI-6400便携式光响应自动测定程序测定,光强由强至弱依次设置为2 000、1 500、1 200、1 000、800、600、400、200、100、50、20、0(µmol/(m2·s)),测试时间选在9∶00-11∶00。被测叶片选用与光合作用日变化测定相同的叶片,共测3株,结果取平均值。光补偿点(LCP,µmol/(m2·s))、光饱和点(LSP,µmol/(m2·s))、最大净光合速率(Pnmax,mmol/(m2·s)),表观量子效率(AQY, mol/mol)等参数根据拟合曲线计算所得。
1.2.3 数据处理 试验数据采用Excel进行预处理和制图,用SPSS16.0进行统计分析。
2 结果与分析
2.1 环境因子的日变化
测定当日太阳PAR、Ca的日变化见图1。由图1可以看出,测定当日光合有效辐射从9∶00开始逐步上升,11∶30达最大,为1 430 µmol/(m2·s),下午逐渐下降。测定当日Ca的变化与PAR的变化相反,呈现早晚高、中午低的趋势,7∶30时Ca值为最高,达390.83 µmol/mol,随后逐渐下降,11∶30降为最低,为353.26 µmol/mol,11∶30以后逐渐回升,17∶00升至380.30 µmol/mol,但仍低于早晨的水平。
测定当日Ta、RH的日变化见图2。从图2中可以看出,Ta的日变化范围在28.04 ~ 35.33℃,早上最低,之后逐步上升,13∶00达最高,随后又逐渐降低。RH的变化趋势与Ta正好相反,早上较高,然后随着气温的上升逐渐下降,15∶00降至最低,随后又逐步上升,变化范围在18.44% ~ 23.34%。
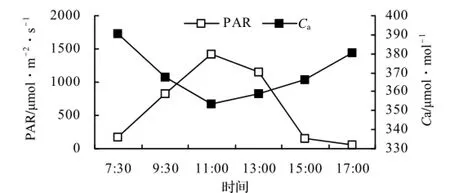
图1 PAR和Ca的日变化Figure 1 Diurnal Change of PAR and Ca
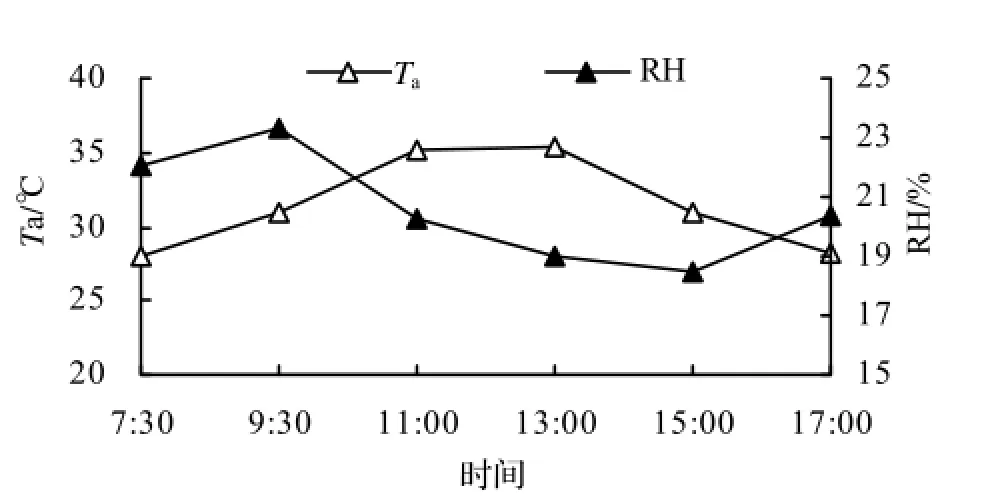
图2 Ta和RH的日变化Figure 2 Diurnal Change of Taand RH
2.2 光合生理参数的日变化
从青钱柳Pn、Tr的日变化(图3)可以看出,Pn的日变化表现为双峰曲线,早上随着PAR的增强,Pn迅速上升,9∶30升至7.84 µmol/(m2·s),达全天中的最大值,这可能由于光照、空气温湿度等环境因子比较适宜所致;9∶30以后,由于PAR、Ta等环境因子的持续升高,Pn出现下降趋势,11∶30左右降为4.72 µmol/(m2·s),出现“午休”现象;11∶30以后随着PAR的下降,Pn又有所回升,13∶00升至5.69 µmol/(m2·s),出现第二个峰值,但比第一个峰值要小;13∶00以后随着PAR的继续下降,Pn逐渐降低,17∶00时降为1.16 µmol/(m2·s)。青钱柳的日平均净光合速率为4.37 µmol/(m2·s)。
Tr的日变化与Pn的日变化进程相似,即从早上开始逐渐上升,9∶30升至1.77 mmol/(m2·s),出现第一个峰值,然后随着Pn的下降,Tr也随之下降,11∶30降为1.48 mmol/(m2·s),11∶30以后又开始上升,13∶00升至2.19 mmol/(m2·s),出现第二个峰值,且较第一个峰值大0.42 mmol/(m2·s),之后随着PAR的降低而逐渐下降,17∶00降为0.27 mmol/(m2·s)。
从图4可以看出,Gs的日变化进程也呈双峰型,即早上逐渐上升,9∶30出现第一个峰值,达0.05 mol/(m2·s),然而开始下降,11∶30降为低谷,之后又开始上升,13∶00出现第二个峰值,为0.04 mol/(m2·s),13∶00以后逐渐下降,17∶00降为0.01 mol/(m2·s)。Gs的日变化进程与Pn、Tr有较强的相似性,说明它们之间的相关性较密切。
Ci早上最大,为188.81 µmol/mol,这可能是由于早上Ca较高所致,然后随着光合作用的增强,Ci逐渐下降,11:30降为最低,为58.79 µmol/mol,11∶30以后随着PAR逐渐减弱,Ci逐渐上升,17∶00时升至 163.22 µmol/mol,较接近于早上水平。
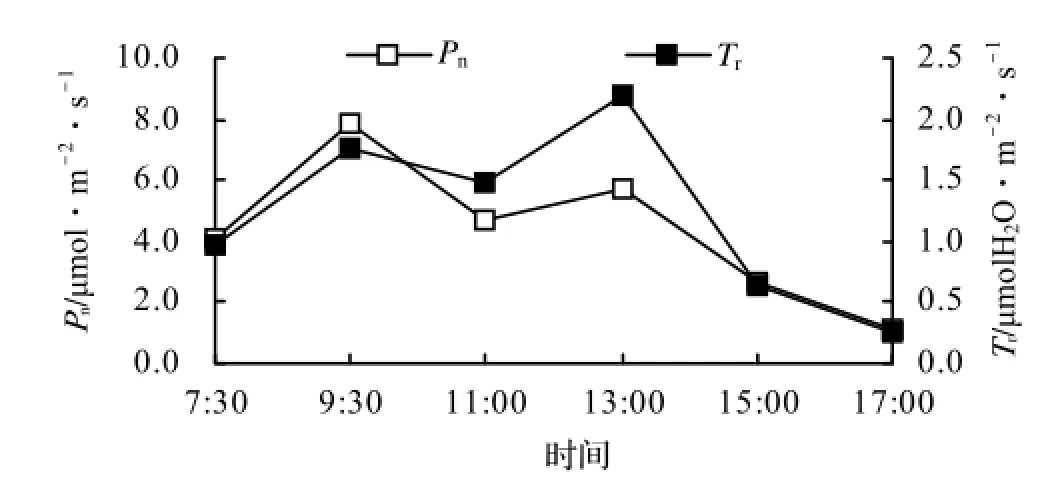
图3 青钱柳Pn、Tr的日变化Figure 3 Diurnal Change of Pnand Tr
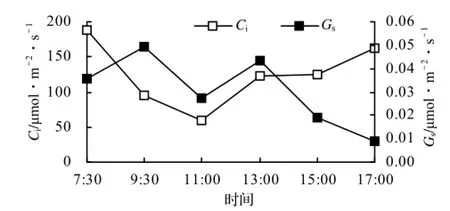
图4 青钱柳Gs和Ci的日变化Figure 4 Diurnal Change of Gsand Ci

图5 青钱柳WUE和Ls的日变化Figure 5 Diurnal Change of WUEand Ls
从图5可以看出,青钱柳叶片的WUE在9:30之前都保持较高水平,此时Pn和Tr都呈上升趋势,但Pn的增幅大于Tr,使得WUE增大;随着光强的逐渐增大和光合“午休”现象的产生,Pn降幅明显,虽有第二次峰值出现,但仍小于Tr的增幅,致使WUE下降,13:00降为最低,为2.59 µmol/mmol;13:00以后随着外界光强和温度的降低,Pn和Tr都开始下降,但Tr的降幅大于Pn,使得WUE又开始回升。
从图5可以看出,Ls为早、晚低,中午高,呈倒“V”字型,即从早上开始逐步上升,11∶30达到最大,为0.83 µmol/mmol,然后缓慢下降,说明午间叶片的气孔效应较明显,这可能是导致产生光合“午休”现象的主要原因。
2.3 光合参数间的相关性分析
表1为青钱柳光合参数间的相关性分析结果。由表1可以看出,青钱柳Pn与Tr、Gs呈正相关,相关系数分别为0.875和0.938,与Ci呈负相关,相关系数为-0.499;Pn与环境因子PAR、Ta、RH呈正相关,相关系数分别为0.643、0.453和0.519,与Ca呈负相关,相关系数为-0.379。显著性检验结果表明,Pn与Gs相关性极显著(P<0.01),与Tr相关性显著(P<0.05),而与其它参数间的相关性不显著。Tr与Gs、PAR和Ta呈显著相关,而Ci与PAR、Ta呈显著负相关,与Ca呈极显著相关。

表1 青钱柳光合参数间的相关性分析Table 1 Correlation analysis on photosynthetic parameters
2.4 青钱柳Pn对光的响应
以光强为自变量,Pn为因变量,根据测得的实验数据绘制青钱柳光响应曲线见图6。从图6可以看出,PAR在0 ~ 400 µmol/(m2·s)范围内,Pn随PAR的增大,上升速度较快,Pn在PAR 400~1 000 µmol /(m2·s)上升速度逐渐减缓,而当PAR超过1 000 µmol /(m2·s)时,Pn呈现微弱的上、下波动,处于光饱和状态。经光合作用与PAR在0 ~ 200 µmol /(m2·s)进行线性回归,得线性回归方程为:y = 0.0303x-0.4393,R2= 0.9768。由此可知,青钱柳的表观量子效率为0.03,暗呼吸速率为-0.4393 µmol/(m2·s),计算得光补偿点为14.63 µmol /(m2·s)。经光合作用与PAR在0 ~ 2 000 µmol/(m2·s)进行二次多项式曲性回归,得回归方程为:y = -5×10-6x2+ 0.0125x + 1.1173,R2= 0.8385,计算得青钱柳光饱和点为1 200 µmol /(m2·s),光饱和时的最大净光合速率为8.32 µmol/(m2·s)。

图6 青钱柳Pn与PAR的响应曲线Figure 6 Response curve of Pnand PAR
3 结论与讨论
植物光合作用是一个复杂的生理过程,其变化不仅受光照、温度、湿度、大气CO2浓度等外在环境因子的综合影响,还与植物内在生理因子相关[18]。从植物光合作用的日变化规律来看,较为常见的曲线类型有双峰型(上、下午各有1个峰值)和单峰型(中午光合速率最高)。本试验结果表明一年生青钱柳播种苗Pn表现为上、下午各有1个峰值的双峰曲线,峰值出现在9:30和13:00左右,11:30左右出现光合“午休”现象。这与已有的研究结果相似,只是出现峰值的时间不同,这可能与测定时间、测试苗木的树龄和种源有关。发生光合“午休”的原因包括气孔限制和非气孔限制因素[19],本试验中青钱柳的Pn在9∶30之前随着外界PAR的升高而升高,9∶30时达到全天中的最大值,而此时PAR尚未达到最大。9∶30-11∶30,随着外界光强和气温的继续增高,Gs急速下降,Ci降低,Ls升高,说明引起青钱柳产生光合“午休”的原因主要是气孔限制因素。因此,在青钱柳的苗期管理过程中夏季温度过高,光照过强时可采取适当遮阴,减轻光合“午休”影响,并加强水分管理。
通过青钱柳Pn与PAR、Ta、Ca、RH和Tr、Gs、Ci的多元相关分析认为,青钱柳Pn与Tr呈显著相关,与Gs呈极显著相关,与Ci呈负相关,而与PAR、Ta、RH呈正相关,与Ca呈负相关,相关性均不显著。而Tr与PAR和Ta呈显著相关,说明光照和温度是影响青钱柳光合参数的主要环境因子,这为青钱柳种植环境的选择和后续管理提供了较好的理论依据。
从青钱柳光合作用及光的响应特征来看,青钱柳的光饱和点较高,为1 200 µmol/(m2·s),光饱和时的Pn为8.32 µmol/(m2·s),光补偿点较低,为14.64 µmol /(m2·s),说明青钱柳对光环境的适应范围较广,光合能力较强,这也验证了青钱柳生态适应性强、生长速度快的生物学特征。
致谢:本试验得到浙江省林业科学研究院杨华博士的大力帮助,在此谨表示谢意!
[1] 中国科学院“中国植物志”编辑委员会. 中国植物志[M]. 北京:科学出版社,2004.
[2] 易醒,石建功,周光雄,等. 青钱柳化学成分研究[J]. 中国中药杂志,2002,27(1):43-45.
[3] 何春年,彭勇,肖伟,等. 青钱柳神茶的应用历史与研究现状[J]. 中国现代中药,2012,14(5):62-68.
[4] 谢明勇,李磊. 青钱柳化学成分和生物活性研究概况[J]. 中草药,2001,32(4):365-366.
[5] 中国药材公司. 中国中药资源志要[M]. 北京:科学出版社,1994.
[6] 冷任轩. 青钱柳的基础理论研究和临床观察[J]. 江西中医药,1994,25(2):64-65.
[7] 上官新晨,陈木森,蒋艳,等. 青钱柳多糖降血糖活性的研究[J]. 食品科技,2010,35(3):82-84.
[8] 谌梦奇,梁锦业,焦志海,等. 青钱柳茶调节血脂作用的临床观察[J]. 中华实用中西医杂志,2002,2(15):863-865.
[9] 李丽红. 青钱柳人工林生长特性研究[J]. 安微农学通报,2012,(11):136-137.
[10] 宋祖祥,邱先华. 青钱柳播种育苗技术试验[J]. 江西林业科技,2004(5):10-11.
[11] 李先明,方升佐,刘晶晶. 青钱柳扦插育苗试验[J]. 林业科技开发,2014(5):124-127.
[12] 方升佐,洑香香. 青钱柳资源培育与开发利用的研究进展[J]. 南京林业大学学报(自然科学版),2007,31(1):95-100.
[13] 徐学华. 作物光能利用率的影响因素及提高途径[J]. 现代农业科技,2011(19):127-127.
[14] 刘福娟. 光合作用的影响因素分析[J]. 农机化研究,2004(4):237-237.
[15] 马金娥,金则新,张方标,等. 青钱柳光合日进程及其与环境因子的相关分析[J]. 安微农业科学,2007,35(16):4 752-4 754.
[16] 杨瑞卿,楚秀丽,杨万霞,等. 不同种源青钱柳苗期光合特征及变异分析[J]. 南京林业大学学报:自然科学版,2012,36(4):41-45.
[17] 黄宏亮,柏明娥,方建华,等. 青钱柳播种育苗及苗期生长特性研究[J]. 山东林业科技,2015(2):40-42, 13.
[18] 武维华. 植物生理学[M]. 北京:科学出版社,2003.
[19] 许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯,1997,33(4):241-244.
Diurnal Variation of Photosynthetic Parameters and Response Character of Cyclocarya paliurus Seedlings
BAI Ming-e1,FANG Jian-hua2,XU Gao-fu3,SHEN Jian-jun1,CHEN Xiu-juan1
(1. Zhejiang Forestry Academy, Hangzhou 310023, China; 2. Chun’an Qiangdaohu Forest Farm of Zhejiang, Chun’an 311700, China; 3. Chun’an Xin’anjiang Development Company of Zhejiang, Chun’an 311700, China)
Determinations were conducted on net photosynthetic rate(Pn), transpiration rate(Tr) stomatal conductance(Gs), intercellular CO2concentration(Ci), and response of 1-year Cyclocarya paliurus seedlings. The result showed that diurnal variation of Pn, Tr, Gshad double peaks occurred at 9∶30 and 13∶00, while that of Citopped in the morning and evening, the lowest at 11∶30. Pnhad extremely significant correlation with Gs(P < 0.01), significant correlation with Tr(P < 0.05). Light compensate point (LCP) was 14.63µmol/(m2·s), light saturation point (LSP) was 1 200µmol/(m2·s), maximum net photosynthetic rate (Pnmax) was 8.32 µmol/(m2·s). The experiment demonstrated that photosynthetically available radiation (PAR) and atmospheric temperature (Ta) had effect on photosynthetic parameters of tested seedlings.
Cyclocarya paliurus; photosynthesis; diurnal variation; response
S792.13
A
1001-3776(2016)01-0001-05
2015-09-23 ;
2015-10-19
浙江省科技厅公益技术研究农业项目(2013C32087);浙江省林业科研成果推广项目(2014B09)
柏明娥(1966-),女,浙江嘉兴人,研究员,从事资源植物开发研究。
