郑州郊区崖沙燕(Ripariariparia)的巢址选择
2016-12-28叶淑英郭书林窦泽龙王振龙路纪琪
叶淑英,郭书林,窦泽龙,王振龙,4,*,路纪琪,4
1 河南教育学院生物系,郑州 450046 2 河南教育学院教育系,郑州 450046 3 郑州大学生命科学学院,郑州 450001 4 郑州大学生物多样性与生态学研究所,郑州 450001
郑州郊区崖沙燕(Ripariariparia)的巢址选择
叶淑英1,郭书林2,窦泽龙3,王振龙3,4,*,路纪琪3,4
1 河南教育学院生物系,郑州 450046 2 河南教育学院教育系,郑州 450046 3 郑州大学生命科学学院,郑州 450001 4 郑州大学生物多样性与生态学研究所,郑州 450001
2014年3—8月,采用样方法对分布于河南省郑州市郊区的4处崖沙燕(RipariaripariaLinnaeus 1758)营巢地的13个生境因子进行了调查,测定了巢址样方和对照样方各160组数据,并利用Mann-Whitney U检验对这些数据的显著水平进行了检验;最后采用逐步判别分析(stepwise discriminate analysis)方法确定影响崖沙燕巢址选择的主要生态因子,并建立了标准化的典则判别函数。结果表明,1)崖沙燕洞巢的洞口横径(8.46±0.79)cm,纵径(8.30±0.79)cm,巢洞深(87.80±11.97)cm,巢室长径(11.25±1.33)cm,短径(10.43±1.33)cm,巢室高(9.99±1.33)cm;2)巢址样方与对照样方在植被均高、植被密度、坡位、坡度、崖高、崖长、崖龄及干扰度等8个方面差异达到显著水平,崖沙燕偏好选择的洞巢周围植被较高(97.24±8.42)cm,植被密度较大(64.34±7.15)株/m2,坡度较大(83.78±0.35)°、上坡位(1.41±0.04)、崖较高处(7.54±0.13)m、崖壁较长(51.18±3.54)m、崖龄较短(2.41±0.05)月及人类干扰较低(1.94±0.06);3)逐步判别分析结果表明,影响崖沙燕营巢地选择的主要生境因子是坡位、崖龄和干扰度,标准化的典则判别函数为y=0.753×坡位+0.681×崖龄+0.288×干扰度。建议在城市化建设过程中针对崖沙燕的巢址选择特性建立合理的保护措施。
崖沙燕;巢址选择;逐步判别分析;城市化
鸟类的巢址选择是研究分析巢及其周围生态因子在鸟类巢址选择过程中的作用和地位,揭示鸟类选择该处筑巢的原因和主导因素[1]。巢址的质量直接影响其繁殖成效[2]。城市鸟类的巢址选择决定其是否能够与人类共生于城市生态系统中,了解鸟类不同物种的生态学需求是对其进行成功保护的先决条件[2- 3]。崖沙燕(Ripariariparia)为雀形目(Passeriformes)燕科(Hirundinidae)沙燕属(Riparia)的一种鸟类,分布于除澳大利亚外的世界各地,特点为集群繁殖,捕食双翅目、半翅目及蚊蝇等昆虫[4],在郑州地区为夏候鸟,主要分布于城郊,对消除城市环境中的害虫,维持城市生态系统稳定性起到重要作用。
目前,有关沙燕属鸟类的研究在国内主要集中于崖沙燕东北亚种(R.r.ijimae, Lonnberg)[4- 6],淡色崖沙燕(R.diluta)[7]的新疆亚种(R.d.diluta)[7- 8]和福建亚种(R.d.fokienensis)[7,9- 11]的洞巢生物学[6,8- 9]和繁殖行为方面的研究[5,10-11],国外主要涉及崖沙燕营巢特点的研究[12- 14],而对该物种在城市生境中生存状况的研究尚未见报道,崖沙燕的巢址选择及影响巢址选择的主要生态因子尚不明确。2014年3—8月对崖沙燕分布在郑州市郊区的营巢地进行了初步研究,旨在揭示其巢址选择的特点和影响巢址选择的主导因素,了解其对生境的基本需求,为城市建设中对该物种的保护提供基础资料。
1 研究区域和研究方法
1.1 研究地与营巢地概况
郑州市位于河南省中部偏北,北临黄河,西依嵩山,东南为广阔的黄淮平原。地理位置为112°42′—114°14′N,34°16′—34°58′ E,在动物地理区划上属古北界,东北亚界、华北区,黄淮平原亚区、豫东平原农作区[15]。该地区属于黄泛区,年均气温14.5℃,年均降雨量700—800 mm。
本研究中4处崖沙燕营巢地的植被类型均为次生及人工栽培种类,主要有:毛白杨(Populustomentosa),臭椿(Ailanthusaltissima),垂柳(Salixbabylonica),雪松(Cedrusdeodara),广玉兰(Magnoliagrandiflora),紫叶李(Prunuscerasifera),白蜡(Fraxinuschinensis),杜英(Elaeocarpussylvestris)及米蒿(Artemisiadalai-lamae),葎草(Humulusscandens),牛筋草(Eleusineindica),狗牙根(Cynodondactylon),马唐(Digitariasanguinalis),藜(Chenopodiumalbum)等。其中1号营巢地生长有芦苇(Phragmitesaustralis),菖蒲(Acoruscalamus)。土质类型为风沙土和壤土[16]。
1.2 研究方法
1.2.1 样地选取
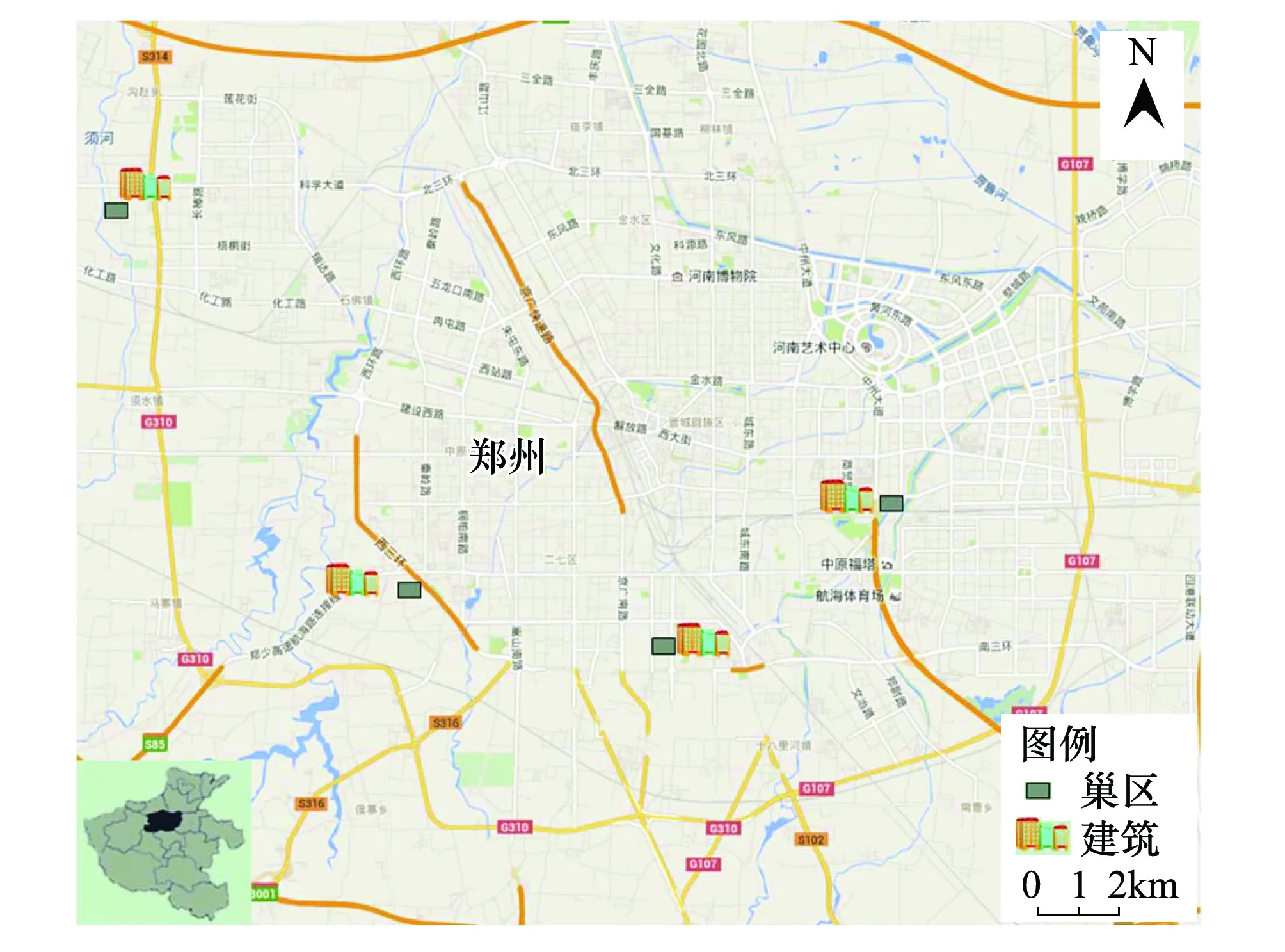
图1 崖沙燕的巢址分布示意图Fig.1 The distribution diagram of Sand Martin′s nest-sites
2014年3—8月,在郑州市郊区共发现并调查了4处崖沙燕的营巢地(图1)。1号巢区位于中州大道东500 m(34°44′ N,113°43′ E),东邻七里河;2号巢区位于南三环北500 m(34°42′ N,113°39′ E);3号巢区位于西三环西300 m(34°43′ N,113°34′ E),西邻南水北调干渠中段;4号巢区位于西四环西1 000 m(34°48′ N,113°30′ E),西邻须河。
1.2.2 数据收集
洞巢样方与对照样方的选取参照程闯等[17]的方法,在4处崖沙燕营巢地的每个营巢面的巢洞密集区域选取1个1 m×1 m 的样方,在4个角的方向上10 m的范围内随机各选取1个1 m×1 m的样方。对照样方的选取是以巢区中心为基准点,在100 m内的非营巢区选取1个1 m×1 m的样方,在4角的方向10 m的范围内随机各选取1个1 m×1 m的对照样方。测定崖长、崖龄、崖高、坡度、坡向、坡位、开阔度、明水面与否、土质、干扰度等10个巢址生境因子。植被样方与对照样方的选取参照闫永峰等[18]与吴逸群等[19]的方法,植被样方是在其觅食区域中心位置选取1个2 m×2 m的大样方,在4角做4个2 m×2 m的灌木样方,并在大样方2条对角线的4等分点及中心做5个1 m×1 m的草本样方。对照样方的选取是在其觅食区域中心为基点的任意方向上的非觅食区域内,距离崖沙燕觅食中心点100 m处作为对照样方中心点,对照样方的选取同样方。分别测定植被高度、植被盖度和植被密度等3个巢址生境因子。测量方法参照闫永峰等[18]、吴逸群等[19]与程闯等[17]的方法:
海拔高度、经纬度在采样地用GPS测定。
明水面与否,指距样地中心500 m范围内是否有溪流、河水、泉水等水源。有水源赋值为1,否则赋值为2。
植被高度(cm),用1m米尺在样方内随机选取20株植被进行测量,以其平均值为植被均高。
植被密度(株/m2),记录样方内植物的数量。
植被盖度(%),在每个样方的对角线上,设置200 cm的调查样线,以2 cm为单位,记录植物所占的格数,二者平均值为植被盖度。
依据张华等[16],4处崖沙燕营巢地的土质类型在1号样地为风沙土,其余为壤土。风沙土赋值为1,壤土赋值为2。
坡度(°)、坡向、坡位用DQL- 8型地质罗盘测定。坡向依东,南,西,北,分别赋值为1、2、3和4;坡位分为上坡位、中坡位和下坡位,分别赋值为1、2和3。没有坡的此3项均记做0。
崖高(m)、崖长(m):在样方所处的断崖处,用5 m的卷尺测量。
崖龄(月),通过访谈4处崖沙燕营巢地的建筑工人和门卫师傅收集崖壁形成的时间数据。
开阔度,以高度在10 m以上的建筑物与调查样方的距离为依据,分别对<1500 m、1500—2500 m、2500—3500 m、3500—4500 m以及>5000 m赋值为1,2,3,4和5。
干扰度,以样方中心与人类活动点的距离为依据,分别对>400 m、300—400 m、200—300 m、100—200 m和<100 m赋值为1、2、3、4和5级。其中5级为强烈干扰,4级为重度干扰,3级为中度干扰,2级为轻度干扰,1级为无人为干扰。
1.2.3 数据处理
巢址样方数据的处理与分析由SPSS 19.0统计软件完成。将百分数因子作反正弦函数转换为角度型因子,角度转化为弧度,以提高数据的正态性。采用独立样本Mann-Whitney U检验分析巢区样方和对照样方间13个变量的差异性,对于差异显著的变量使用Spearman判断变量之间的相关性,选取相关系数差异显著的生物学意义比较重要的变量进入逐步判别分析(stepwise discriminate analysis)予以筛选分析。所有数据用平均值士标准误(Mean±SE)表示。P<0.05为差异显著。
2 结果与分析
2.1 崖沙燕洞巢构造及密度相关参数测量
崖沙燕的洞巢位于开阔度较大的建筑工地新挖掘的深9 m左右地基坑的垂直断面崖壁,或者高7 m左右的堆土回填坑基所致的垂直断面上。巢洞呈坑道状,内洞低于洞口3—5 cm,且多向左或向右弯曲,洞口呈扁椭圆形,终端膨大为巢,洞口横径((8.46±0.79)cm,n=34),纵径((8.30±0.79)cm,n=34),巢洞深((87.80±11.97)cm,n=34),巢室长径((11.25±1.33)cm,n=10),短径((10.43±1.33)cm,n=10),巢室高((9.99±1.33)cm,n=10)。巢材多选择芦苇、狗牙根、马唐、牛筋草、米蒿等的茎叶,小鷿鷈(Tachybaptusruficollis)、黑水鸡(Gallinulachloropus)、白鹭(Egrettagarzetta)的羽毛及棉花、窗纱、细绒线、塑料袋、细铁丝等城区常见废弃物品。崖沙燕洞巢呈现聚集分布,未见单独营巢现象,聚集区域的平均面积为((14.34±1.23)m2,n=15),洞巢密度最高可达34个/ m2。在聚集分布区内,巢洞水平间距((12.16±0.79)cm,n=34),巢洞垂直间距((16.88±0.79)cm,n=34)(图2)。

图2 崖壁上崖沙燕洞巢分布(A)及巢内崖沙燕的卵(B)Fig.2 The Sand Martin nests distributed on the cliffs (A) and the eggs in their nests (B)
在郑州市郊区共调查了4处崖沙燕营巢地,其中1号样地有695个洞巢,分布于东向和北向的地基断面上;2号样地有洞巢290个,分布于在建工地开挖地基堆积土西向和南向的土崖上;3号样地有洞巢320个,分布于修筑渠道堆积土北向和东向的崖壁上;4号样地有洞巢2500个,分布于刚挖的地基断面东向和西向的崖壁上。
2.2 巢址样方和对照样方的13种变量的差异性比较
对崖沙燕巢址样方和对照样方的13个变量进行Mann-Whitney U检验,其中8个变量间的差异达到了显著水平(表1)。崖沙燕洞巢周围的植被较高(U=11109.00,P=0.041)、植被密度较大(U=9515.50,P<0.001)、坡度较大(U=2872.00,P<0.001)、上坡位(U=3387.50,P<0.001)、崖较高处(U=9915.50,P<0.001)、崖壁较长(U=9283.00,P<0.001)、崖龄较短(U=3540.00,P<0.001)及人类干扰较低(U=9300.00,P<0.001)。
2.3 巢址样方和对照样方差异显著变量的逐步判别分析结果
巢址样方和对照样方差异显著变量进行Speaman相关分析(表2)。植被均高、植被密度、坡位、坡度、崖长、崖龄、崖高和干扰度等8个变量的相关性差异显著。利用这8个变量进行逐步判别分析,共提出坡位,崖龄和干扰度等3个变量,Wilk′s λ值分别为0.588,0.530和0.406(表3)。表明这3个变量对区分崖沙燕的巢址与非巢址样方贡献力最大的变量是其巢区所在崖壁的坡位,同时说明研究区域内崖壁的崖龄和干扰度是影响崖沙燕巢址选择的重要因素。3个因子的判别函数对160组洞巢样方的判别回代结果准确率为92.50%,对160组对照样方的判别回代结果准确率为85.63%。标准化的典则判别函数为y=0.753×坡位+0.681×崖龄+0.288×干扰度。
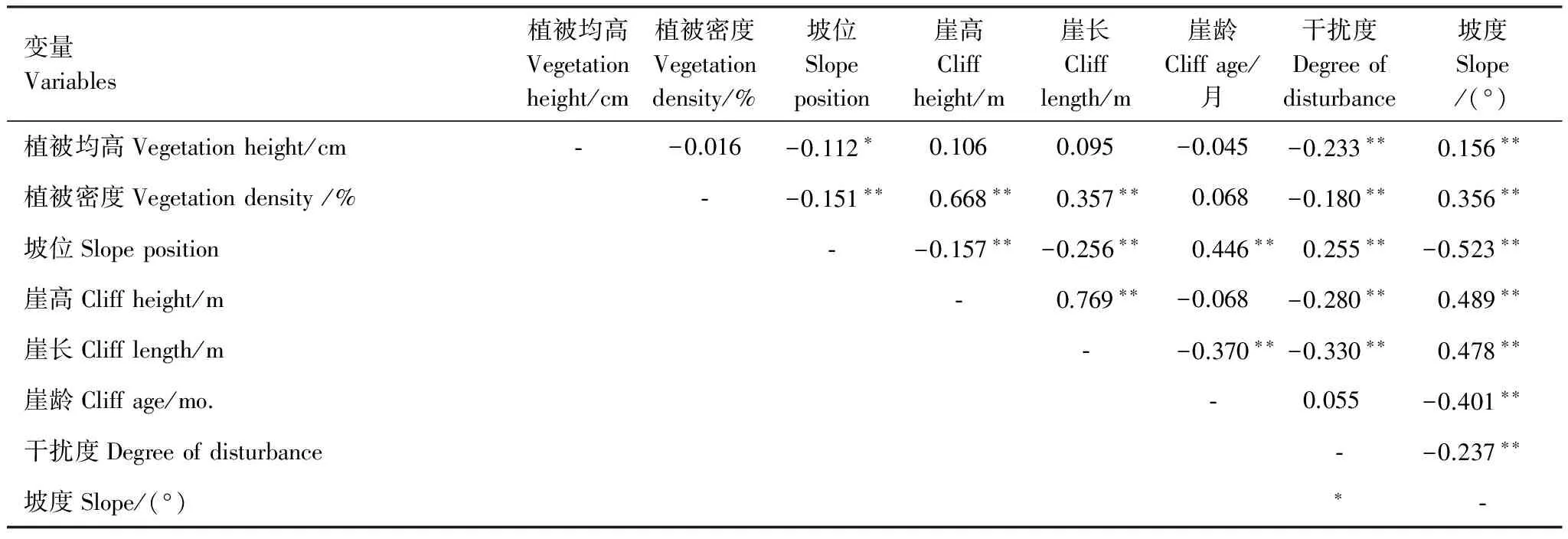
表2 崖沙燕巢址样方与对照样方变量间相关系数
*P<0.05, **P<0.001
表 3 崖沙燕巢址样方与对照样方变量的逐步判别分析结果
Table 3 Consequence of the stepwise discriminate analysis of variables between the nest-site samples and control-site samples of Sand Martin
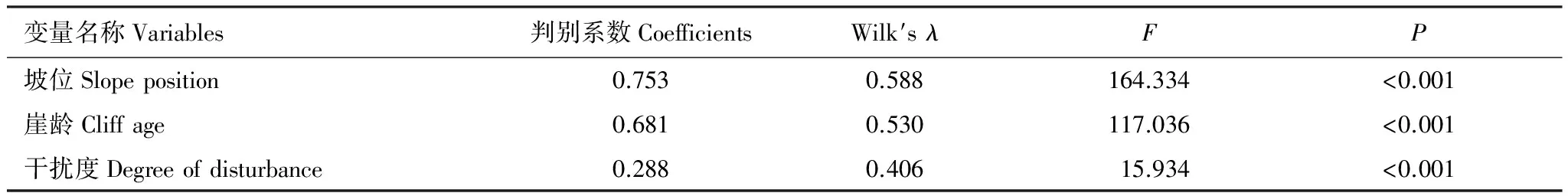
变量名称Variables判别系数CoefficientsWilk′sλFP坡位Slopeposition0.7530.588164.334<0.001崖龄Cliffage0.6810.530117.036<0.001干扰度Degreeofdisturbance0.2880.40615.934<0.001
3 讨论
3.1 筑巢崖壁与洞巢特征
在巢址选择过程中,许多鸟类都倾向于选择那些能使其繁殖成效最大而存活代价最小的营巢生境[20]。崖沙燕对筑巢崖壁的新旧具较强的选择性,并趋于选择在新形成的2个月左右低崖龄的崖壁上筑巢。低崖龄的沙壤土质能够降低崖沙燕啄巢时的能量消耗,这和Smalley等[12]关于崖沙燕偏好选择可降低筑巢能耗的黄壤土质的研究结果类似;将洞巢建在新形成的崖壁上,可降低寄生虫的危害[17,21],崖沙燕对土崖壁的选择和栗喉蜂虎(Meropsphilippinus)[17,21]极其类似,这也许是营集群洞巢繁殖鸟类的一种生殖策略。同时,郑州郊区4处建筑工地的掘土施工期可能刚好和崖沙燕的迁入期相吻合,使得崖沙燕能够选择低崖龄的土崖筑巢;有研究指出沙燕属鸟类多筑巢于水边崖壁的沙土与黄壤交错的沙土层,且喜利用旧巢洞[4- 6,8- 9,13]。然而本研究中观察到1号营巢地尚有旧巢存在,但其却并未利用而选择3个月以内形成的土崖筑新洞巢,原因可能是旧巢经过长时间的雨水冲刷坡度远低于垂直面,巢内微生物累积过多风险过高所致,或者建筑工地由于施工需要很难有大量旧巢的土崖存在,导致样本量过低,无法真实反映崖沙燕是否选择利用旧洞巢,但是连续3a对郑州郊区崖沙燕于1,2,3号营巢地的观察发现,该物种具有选择旧巢址附近再筑新巢的偏好性。为什么郑州郊区的崖沙燕对新巢址的选择偏好优于旧洞巢,这是今后的研究中需要关注的重点。Etxezarreta和Arizaga[13]研究发现崖沙燕可将人工河墙上的建筑管道当作洞巢,我们的研究尚未发现此类现象,期待今后更丰富的观察数据充实崖沙燕的巢址选择特点。
巢洞作为崖沙燕的繁殖和栖息场所,适宜的洞巢深度和巢室大小能够保证洞巢内部环境稳定,利于孵化和育雏的正常进行[21]。郑州郊区崖沙燕的外洞口测量结果(8.46±0.79)cm×(8.30±0.79)cm 和淡色崖沙燕福建亚种[9]的(7.12±1.09)cm×(5.18±0.83)cm及崖沙燕东北亚种[6](11.16±2.254)cm×(7.02±0.740)cm的研究结果不同,推测主要原因可能是,1号巢地特殊的沙质土松散结构,育雏期间的崖沙燕亲鸟每天频繁的进出洞巢及附着于巢洞口饲喂幼雏加快了外洞口风沙土的掉落,使得外洞口纵径变大,或者不同物种(亚种)鸟类对巢洞口的大小要求不同;郑州郊区崖沙燕的巢洞深度(87.80±11.97)cm与崖沙燕东北亚种(89.35±18.27)cm[6]的差异不大,比淡色崖沙燕福建亚种[9](64.38±11.73)cm的深,这可能与物种差异或营巢地区气候有关。
3.2 土崖坡位与人类干扰
崖沙燕偏好选择在中上坡位(P<0.001),且陡峭程度高(P<0.001)的土崖坡上筑巢,可降低与天敌的接触,进而减少子代被捕食的机会[21- 22],这与韩联宪等[21]对小型鸟类营洞巢生殖策略及吴逸群等[19]对大型鸟类巢址选择的研究结果类似。坡度大且位于中上坡位,除了人类之外几乎没有天敌。人类和其他捕食动物的干扰程度相对较低,有利于崖沙燕在危险情况下紧急逃避敌害,降低捕食风险。
较低的干扰是崖沙燕巢址选择的重要生态因子之一。4处营巢地的观察发现,崖沙燕所选择的建筑工地都比较开阔,巢区上方几乎没有任何遮蔽,且巢区周围没有高大的植被,崖沙燕对周围环境能一览无余,一旦环境有变化,其能很快洞悉并及时应对。1号、2号和4号3处巢地的四周均有2 m高的密闭护栏遮挡,人类干扰小,相对环境比较安静,加之工地环境复杂非工地相关人员很难很少进入,又位于城乡结合部,远离人群入住的高大建筑,觅食生境开阔远离道路,在一定程度上更加降低了人为干扰,节省了能量成本,降低了存活代价,因而该物种更适于在城市郊区环境中生存。
3.3 洞巢周围植被与水源
野生动物的生存离不开生境,其中食物、水和隐蔽条件是野生动物生境选择的3大要素[23]。植被密度、植被盖度、植被均高等是影响鸟类栖息地选择的重要因子[24],也是鸟类主要觅食地和隐蔽因子。尽管郑州郊区的崖沙燕偏好选择植被密度较大与植被均高较高的生境,然而逐步判别分析的结果并未显示其是影响崖沙燕巢址选择的主要因子。这可能与沙燕属鸟类常集大群于空中飞行捕食昆虫的习性有关[4,6,8- 9],同时,崖沙燕筑于断崖中上坡位深约1 m的洞巢对防御天敌而言本身就是一种很好的隐蔽。Etxezarretaet和Arizaga[13]及国内的研究者[4- 6,8- 9]指出不同种属的崖沙燕对巢址的选择均偏好明水面附近,但研究结果却未显示出其对大的明水面的极强的选择性(如2号和3号巢区周围3 km范围内没有明水面,2号巢区位于南水北调中线冯湾段附近,而该段仅在7月23日以后开始储水)。然而实质上崖沙燕的洞巢附近通常会有人工种植植被定时浇灌而形成的面积大小不等的明水区域,并非一定存在大的明水面。鸟类对巢址生境因子的选择偏好具有明显差异,是崖沙燕对城市生境适应的一种反映。推测明水面和适合筑巢的土崖相比,崖沙燕则更倾向于选择合适的土崖筑巢,或者环境容纳量及“先到者”的优势会使得崖沙燕的巢址选择更难反映其真实意愿[25]。
英国鸟类生态学家Lack研究指出,鸟类能通过识别环境中的某些特征来主动选择环境,如果没有这些特征,即使环境中包含动物所有必需的资源,其仍然不选择在该环境中生活[26]。可见鸟类对栖息地的选择是一个多层次的各种生态环境因子综合作用的复杂的过程[27- 28]。郑州郊区的崖沙燕之所以会选择2号和3号营巢地远离水源、土崖高度较低(约7 m)、人类干扰较大的区域筑巢,推测主要原因是该区域具有适宜其繁殖的地理环境特征,另一方面也反映出崖沙燕选择一处适宜筑巢的生境已愈来愈趋于艰难。然而由于施工方受工期制约,往往会导致工人为赶工期而于尚未出飞的崖沙燕不顾,野蛮施工使得许多幼龄崖沙燕死于非命。如郑州郊区2014年的4处建筑工地只有1号营巢地的崖沙燕幼雏最为幸运成功出飞。
4 建议
由于崖沙燕偏好选择位于城乡结合部的建筑工地新挖掘的接近垂直断面的砂、壤质崖壁上筑巢,其筑巢期在4—5月,雏鸟全部出飞弃巢时期为7—8月,在巢期约4个月。因此在城市化建设中针对崖沙燕的保护提出如下建议:(1)加强动物保护宣传力度,增强市民爱鸟护鸟意识。(2)尽量避免在崖沙燕迁入前3个月内挖掘新土,如果建筑工地必须赶工期,有必要对新挖的崖壁或土堆进行遮盖,避免崖沙燕筑巢;对已经筑巢1—2周的可以实施驱赶避免其繁殖。(3)相关部门应该积极开辟崖沙燕的适宜栖息地,如西流湖,西四环须河、新郑龙湖、中牟雁鸣湖等周围生境,既能够满足崖沙燕繁殖筑巢的需求,保护其种群,也能够满足人们观鸟识鸟的需要。
致谢:河南观鸟协会王鹏先生提供崖沙燕迁入信息,郑州东风建筑工程有限公司在数据采集中提供支持,郑州大学田军东博士和中国科学技术大学张振宇教授帮助写作,特此致谢。
[1] Cody M L. Habitat selection in birds: the roles of vegetation structure, competitors, and productivity. BioScience, 1981, 31(2): 107- 113.
[2] Rands M R W. The effect of nest site selection on nest predation in Grey PartridgePerdixperdixand Red-legged PartridgeAlectorisrufa. Ornis Scandinavica, 1988, 19(1): 35- 40.
[3] 巴明廷, 李长看, 李元奇, 崔淑贞, 李学德. 郑州城区鸟类冬季集群调查分析. 河南农业大学学报, 2003, 37(3): 274- 277.
[4] 高玮. 中国东北地区鸟类及其生态学研究. 北京: 科学出版社, 2006: 176- 177.
[5] 李振营, 王学全. 灰沙燕的繁殖生态. 野生动物学报, 1985, (4): 13- 15.
[6] 傅松华, 李晓民. 灰沙燕巢. 国土与自然资源研究, 2001, (4): 74- 75.
[7] 郑光美. 中国鸟类分类与分布名录. 北京: 科学出版社, 2005: 161- 165.
[8] 侯兰新, 吕明, 马力. 灰沙燕的洞巢. 动物学杂志, 1996, 31(5): 50- 51.
[9] 周友兵, 张文广, 张璟霞, 张君, 胡锦矗, 余志伟. 四川南充崖沙燕的洞巢生物学. 动物学杂志, 2004, 39(2): 66- 69.
[10] 周昭敏, 周友兵, 胡锦矗, 庄华, 朱维, 陈涛. 崖沙燕繁殖期雄鸟的鸣叫行为. 动物学杂志, 2004, 39(6): 77- 81.
[11] 周昭敏, 周友兵, 胡锦矗, 庄华, 朱维, 程涛. 崖沙燕繁殖期雄鸟的占巢行为. 西华师范大学学报: 自然科学版, 2005, 26(3): 287- 290.
[12] Smalley I, Blake-Smalley R, O′Hara-Dhand K, Jary Z, Svircev Z. Sand martins favour loess: How the properties of loess ground facilitate the nesting of sand martins/bank swallows/uferschwalben (RipariaripariaLinnaeus 1758). Quaternary International, 2013, 296: 216- 219.
[13] Etxezarreta J, Arizaga J. Characteristics of sand martinRipariaripariacolonies in artificial river walls. Ardeola, 2014, 61(1): 127- 134.
[14] Heneberg P. Sex-specific differences in Sand MartinRipariariparianest attentiveness. Bird Study, 2011, 58(1): 107- 110.
[15] 路纪琪, 王振龙. 河南啮齿动物区系与生态. 郑州: 郑州大学出版社, 2012: 14- 43.
[16] 张华, 任天祥, 孔牧, 马生明, 蒋瑞金. 郑州地区土壤背景研究在环境评价中的应用. 物探与化探, 1993, 17(4): 241- 250.
[17] 程闯, 韩联宪. 云南怒江河谷栗喉蜂虎营巢地特征. 四川动物, 2011, 30(6): 932- 936.
[18] 闫永峰, 刘迺发. 东大山自然保护区喜马拉雅雪鸡(Tetraogallushimalayensis)的巢址选择. 生态学报, 2009, 29(8): 4278- 4284.
[19] 吴逸群, 刘迺发. 甘肃南部蓝马鸡的巢址选择. 生态学杂志, 2010, 29(7): 1393- 1397.
[20] 张正旺, 梁伟, 盛刚. 斑翅山鹑巢址选择的研究. 动物学研究, 1994, 15(4): 37- 43.
[21] 韩联宪, 程闯, 吴忠荣, 匡中帆. 栗喉蜂虎营巢行为与巢洞特征. 西南林业大学学报, 2012, 32(6): 73- 77.
[22] Velando A, Freire J. How general is the central-periphery distribution among seabird colonies? Nest spatial pattern in the European shag. The Condor, 2001, 103(3): 544- 554.
[23] 马建章, 邹红菲, 贾竞波. 野生动物管理学 (第二版). 哈尔滨: 东北林业大学出版社, 2004: 53- 57.
[24] 孟晓静, 张克勤, 邓文洪. 黑枕黄鹂巢址选择研究. 北京师范大学学报: 自然科学版, 2014, 50(2): 174- 177.
[25] 张微微, 马建章, 李金波. 骨顶鸡(Fulicaatra)的巢址选择研究. 东北师大学报: 自然科学版, 2010, 42(3): 114- 120.
[26] Lack D. Habitat selection in birds. With special reference to the effects of afforestation on the Breckland avifauna. Journal of Animal Ecology, 1933, 2(2): 239- 262.
[27] 魏辅文, 冯祚建, 王祖望. 野生动物对生境选择的研究概况. 动物学杂志, 1998, 33(4): 48- 52.
[28] 杨维康, 钟文勤, 高行宜. 鸟类栖息地选择研究进展. 干旱区研究, 2000, 17(3): 71- 78.
Nest-site selection of sand martins (Ripariariparia) in the suburbs of Zhengzhou, Henan Province
YE Shuying1, GUO Shulin2, DOU Zelong3, WANG Zhenlong3,4,*, LU Jiqi3,4
1HenanInstituteofEducation,DepartmentofBiology,Zhengzhou450046,China2HenanInstituteofEducation,DepartmentofEducation,Zhengzhou450046,China3ZhengzhouUniversity,SchoolofLifeSciences,Zhengzhou450001,China4ZhengzhouUniversity,InstituteofBiodiversityandEcology,Zhengzhou450001,China
Bird nest-site selection is an important topic in avian ecology, especially with respect to bird reproduction success. In the past decades, urban avian ecology has attracted more attention, because determination of how birds adapt to urban areas could improve the ecological functioning of urban ecosystems and help to build a better living environment for both the human being and urban wildlife. Sand martin (RipariaripariaLinnaeus 1758), a bird species belonging to Passeriformes, Hirundinidae,Riparia, is distributed throughout the world except for Australia. In recent years, sand martins have become an urban bird that can reproduce in Zhengzhou City, Henan Province, the People′s Republic of China. The sand martin predates Diptera, mosquitoes, and other insects, and therefore plays an important role in eliminating insect pests from urban environments and maintaining ecological stability in cities. However, in the Zhengzhou suburb, there is a conflict between human urbanization and sand martin nest-site selection. Sand martin nest-sites are mainly located on vertical sections of cliffs, which are formed by new mining tunnels or by the hillock formed after bulldozers backfill the tunnel base. To alleviate the conflict and promote eco-city construction, it is necessary to understand the habitat characteristics of the sand martin and the dominant factors affecting its nest-site selection. A survey was conducted to study the nest-site selection of four sand martins in the suburb of Zhengzhou, using a plot method based on field observations from March to August 2014. For either burrow nests or control areas, a total of 160 data sets were collected, and Mann-Whitney U test was adopted to analyze the data. The main ecological factors affecting sand martin nest-site selection were determined by stepwise discriminate analysis, and the Standardized Canonical Discriminant Function was formed. The results showed that 1) the nest hole diameter was (8.46±0.79) cm, longitudinal diameter was (8.30±0.79) cm, depth was (87.80±11.97) cm, nest chamber diameter was (11.25±1.33) cm, longitudinal diameter was (10.43±1.33) cm, and height was (9.99±1.33) cm. 2) There were significant differences in vegetation height, vegetation density, slope position, slope, cliff height, cliff length, cliff age, and degree of disturbance between nests and control areas. Sand martins preferred higher vegetation (97.24±8.42) cm, greater vegetation density (64.34±7.15) plant/m2, steeper slopes (83.78±0.35) °, higher slope positions (1.41±0.04), greater cliffs (7.54±0.13) m, longer cliffs (51.18±3.54) m, smaller cliffs age (2.41±0.05) month, and a lower degree of disturbance (1.94±0.06) of the habitat surrounding the nests than the control areas. 3) The results of the stepwise discriminate analysis showed that the main factors affecting sand martin nest-site selection were slope position, cliff age, and degree of disturbance. The Standardized Canonical Discriminant Function wasy=0.753×slope position+0.681×cliff age+0.288×degree of disturbance. Based on our findings, we offer several suggestions for protecting the sand martin according to the characteristics of its nest-site selection during urbanization.
Sand martin (Ripariariparia); nest-site selection; stepwise discriminate analysis; urbanization
河南省重点科技攻关基金资助项目(122102310400)
2014- 12- 12;
日期:2016- 03- 03
10.5846/stxb201412122472
*通讯作者Corresponding author.E-mail: wzl@zzu.edu.cn
叶淑英,郭书林,窦泽龙,王振龙,路纪琪.郑州郊区崖沙燕(Ripariariparia)的巢址选择.生态学报,2016,36(21):7006- 7013.
Ye S Y, Guo S L, Dou Z L, Wang Z L, Lu J Q.Nest-site selection of sand martins (Ripariariparia) in the suburbs of Zhengzhou, Henan Province.Acta Ecologica Sinica,2016,36(21):7006- 7013.
