不同林龄刺槐林土壤团聚体化学计量特征及其与土壤养分的关系
2016-12-28赵发珠韩新辉杨改河白孙宝郝文芳
孙 娇,赵发珠,韩新辉,杨改河,白孙宝,郝文芳
1 西北农林科技大学生命科学学院, 杨凌 712100 2 宁夏农林科学院农业资源与环境研究所, 银川 750002 3 西北大学城市与环境学院, 西安 710127 4 西北农林科技大学农学院, 杨凌 712100 5 陕西省安塞县退耕办, 安塞 717400
不同林龄刺槐林土壤团聚体化学计量特征及其与土壤养分的关系
孙 娇1,2,赵发珠3,韩新辉4,杨改河4,白孙宝5,郝文芳1,*
1 西北农林科技大学生命科学学院, 杨凌 712100 2 宁夏农林科学院农业资源与环境研究所, 银川 750002 3 西北大学城市与环境学院, 西安 710127 4 西北农林科技大学农学院, 杨凌 712100 5 陕西省安塞县退耕办, 安塞 717400
土壤团聚体化学计量特征分析可以为土壤养分的评价提供依据,对陕北黄土丘陵区20 a、25 a、40 a、50 a刺槐林土壤团聚体有机碳、全氮、全磷化学计量比及其与土壤有机碳、全氮、全磷化学计量比的相关性采用逐步回归分析方法进行了分析。结果表明:随着林龄的增加,刺槐林各粒径土壤团聚体有机碳、全氮含量及其有机碳、全氮、全磷化学计量比显著增加(P<0.05),均表现为在0—20 cm土层高于20—40 cm土层,而刺槐林土壤团聚体全磷含量变化较小;相同林龄刺槐林在0—20 cm和20—40 cm土层中0.25—2 mm粒径土壤团聚体有机碳、全氮、全磷含量及其化学计量比最高。刺槐林0.25—2 mm粒径团聚体对土壤原土有机碳、全氮含量及其有机碳、全氮、全磷化学计量比有显著影响。营造刺槐林对各粒径土壤团聚体全效养分分配及其平衡关系存在积极的影响,主要体现在0.25—2 mm粒径土壤大团聚体中,通过影响0.25—2 mm粒径团聚体提高了土壤全效养分的供应和保持能力。
水稳性土壤团聚体;刺槐林;化学计量特征
生态化学计量学是综合生物学、化学和物理学等基本原理,利用生态过程中多重化学元素之间的平衡关系,分析各化学元素之间平衡对生态系统交互作用影响的一种理论[1]。自1958年A. Redfield首次证明了海洋浮游生物的有机碳(C)、全氮(N)、全磷(P)有特定组成以来,生态化学计量学的研究受到众多学者的广泛关注[2]。研究跨越了个体[3]、种群、群落[4]、生态系统[5]、景观[6]和区域[7]各个层次,内容主要集中在生态过程中C、N和P之间的比例关系、生态系统中限制性养分的判断以及C/N/P与生物生长率关系等[8],而研究对象主要集中于植物组织和土壤的化学计量学特征[9-10],针对土壤团聚体生态化学计量特征的研究较少。土壤团聚体是土壤结构稳定的基本单元,是土壤肥力保持的决定因素[11]。土壤团聚体粒径的变化和土壤养分含量密切相关。不同粒径土壤团聚体C、N和P的存在形态和稳定性不同[12],在土壤肥力演变中有不同的变化趋势[13]。因此,了解土壤团聚体C、N、P生态化学计量特征,对掌握土壤全效养分的供应和保持能力具有重要的指示作用。
刺槐(RobiniapseudoacaciaLinn.)根系发达、生长迅速、成活率高,具有适应性强、耐干旱、耐瘠薄的特点,是我国黄土丘陵区人工造林面积最大的树种[14]。目前关于黄土丘陵区刺槐林土壤养分的研究已有报道,但主要集中于分析不同纬度[15]、不同林龄[14]土壤养分含量或者储量的差异,刺槐与其他植被类型土壤团聚体养分含量比较[16];而从生态化学计量角度对不同林龄刺槐林土壤团聚体及其与土壤养分的相关性研究还缺乏报道。本文通过对陕北黄土丘陵区20 a、25 a、40 a、50 a刺槐林0—20 cm、20—40 cm土层土壤团聚体C、N、P化学计量特征及其与土壤C、N、P化学计量特征的关系进行研究,旨在揭示刺槐林土壤养分的演化机制,以期为了解黄土丘陵区土壤养分的演化提供一定依据。
1 研究地区和研究方法
1.1 研究区概况
研究区域位于陕北安塞县境内(105°05′ 44″E,36°30′ 45″N),海拔997—1731 m,属于暖温带半干旱气候区。年均降雨量535 mm,干燥度1.48,年均气温8.8 ℃,无霜期160 d左右,年总辐射量552.6 kJ/cm2。地貌类型为典型丘陵沟壑区,土壤以黄土母质发育的黄绵土为主;土壤瘠薄、土质疏松、抗侵蚀性能差,是我国西北典型的生态环境脆弱区。植被带属于暖温带落叶阔叶林向干旱草原过渡的森林草原区,建群种有刺槐、侧柏(PlatycladusorientalisLinn.)、柠条(CaraganakorshinskiiKom.)和白羊草(BothriochloaischaemumLinn.)等[17]。
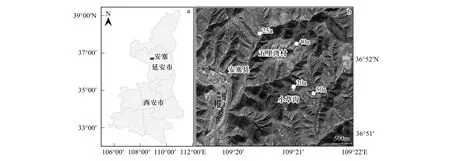
图1 样地分布图Fig.1 Map of research region (Ansai County, Shaanxi Province of China)
1.2 样地选择
通过对不同林龄刺槐林地理位置、成土母质等因素综合考虑,选择20a刺槐林地(20 a)、25a刺槐林地(25 a)、40a刺槐林地(40 a)、50a刺槐林地(50 a)为研究对象(图1)。各刺槐林自然生长,无人为管理措施,干扰较轻,其样地面积为60 m×60 m,实测样地的地理坐标、海拔、坡向、坡位和坡度等信息。样地概况见表1。
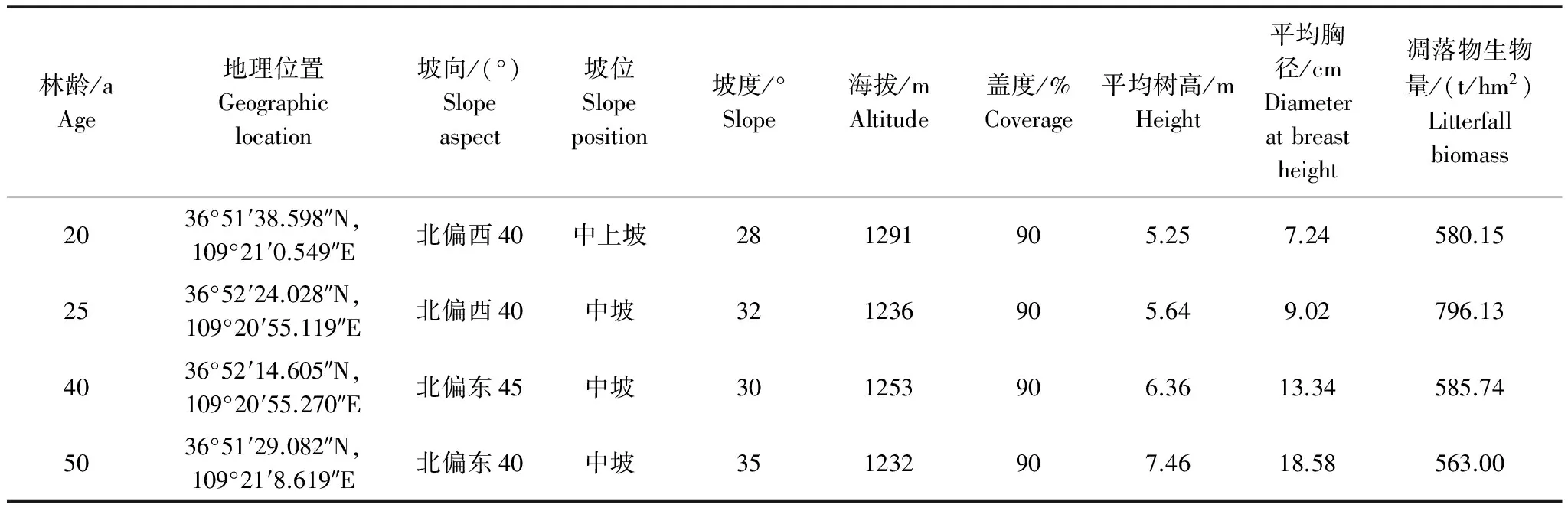
表1 样地概况
1.3 样品采集
于2013年9月份,在每个刺槐林地设置3个样方(20 m × 20 m),每个样方内沿对角线设置5个采样点,在0—20 cm、20—40 cm两个土层分别采取原状土样,存于铝盒中。将原状土样沿其自然结构轻掰成直径约为1 cm左右的小土块,剔除动植物残体和小石块,置于阴凉通风处风干。
1.4 测定方法和计算
采用湿筛法[18]测定水稳性团聚体含量,得到大于5 mm、2—5 mm、1—2 mm、0.5—1 mm、0.25—0.5 mm、0.053—0.25 mm粒径的团聚体后,在50 ℃下烘干、称重,粉碎过100目筛备用。另一部分未湿筛的土样用于土壤常规分析。土壤有机碳的测定采用重铬酸钾-浓硫酸外加热法;全氮的测定采用半微量凯氏法;全磷的测定采用NaOH碱熔-钼锑抗比色法[19]。不同林龄刺槐林土壤基本理化性质见表2,土壤团聚体的粒径分布见表3。
1.5 数据分析
利用Excel 2003和SPSS 19.0对实验数据进行统计分析,采用单因素方差分析(one-way ANOVA)和最小显著差数法(LSD)进行差异显著性检验;通过逐步回归分析和回归系数来描述各测量指标的相关程度。图表的制作采用Origin 9.0和Excel 2003软件处理。
2 结果分析
2.1 不同林龄刺槐林土壤团聚体有机碳、全氮、全磷含量分布特征
如图2所示,不同林龄刺槐林土壤团聚体有机碳含量变化范围在0.92—5.98 g/kg之间,表现为0—20 cm土层高于20—40 cm土层。0—20 cm土层各粒径土壤团聚体有机碳含量随着林龄的增加呈显著增加的趋势(P<0.05)。20—40 cm土层各粒径土壤团聚体有机碳含量的变化表现为50 a显著大于40、25 a显著大于20、25、40 a各粒径土壤团聚体有机碳含量无显著差异。
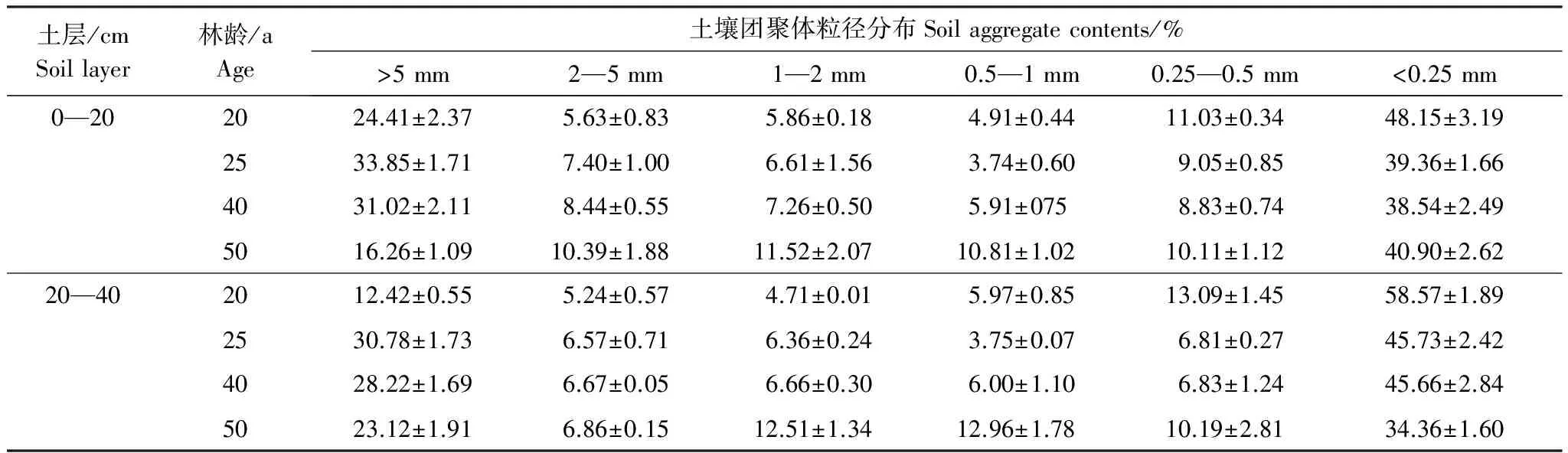
表3 不同林龄刺槐林土壤水稳性团聚体的粒径分布
不同林龄刺槐林0—20 cm和20—40 cm土层土壤团聚体全氮含量变化范围在0.15—0.48 g/kg之间(图3)。各粒径土壤团聚体全氮含量变化在0—20 cm土层中从高到低的顺序大多表现为50 a> 40 a> 25 a>20 a,其中40 a和50 a之间同粒径团聚体全氮含量无显著性差异。20—40 cm土层大于0.25 mm各粒径团聚体全氮含量的变化趋势与有机碳含量的相同,即50 a显著大于40、25 a显著大于20、25 a和40 a之间各粒径团聚体全氮含量无显著差异,小于0.25 mm粒径团聚体全氮含量从高到低的顺序为50 a>25 a>40 a>20 a。
如图4所示,不同林龄刺槐林土壤团聚体全磷含量变化范围在0.47—0.55 g/kg之间。在0—20 cm土层中,大于2 mm粒径土壤团聚体全磷含量随着林龄的增加呈增大的趋势,其中50 a土壤团聚体全磷含量显著高于其他林地,各林地土壤团聚体全磷含量变化在0.25—0.5 mm和1—2 mm粒径中从高到低的顺序为50 a>25 a>20 a>40 a,0.5—1 mm粒径中为25 a>20 a>50 a>40 a,土壤团聚体全磷含量变化在小于0.25 mm粒径中则为50 a>20 a>40 a>25 a。20—40 cm土层,刺槐林土壤团聚体全磷含量的变化趋势为:在大于5 mm粒径中随着林龄的增加呈增大的趋势,在其他粒径土壤团聚体全磷含量变化从高到低的顺序为50 a>40 a>20 a>25 a。
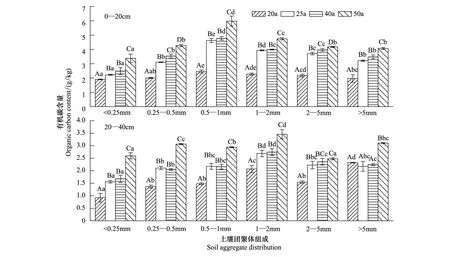
图2 土壤水稳性团聚体中有机碳的含量Fig.2 Content of organic carbon in soil water-stable aggregates不同大写字母表示不同林龄同粒径团聚体在P<0.05水平上差异显著,不同小写字母表示同林龄不同粒径团聚体在P<0.05水平上差异显著
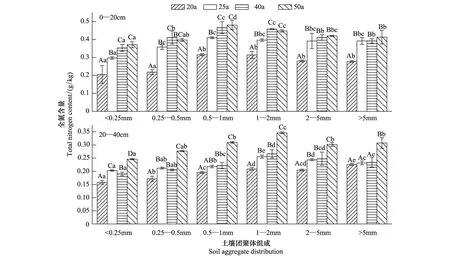
图3 土壤水稳性团聚体中全氮的含量Fig.3 Content of total nitrogen in soil water-stable aggregates
相同林龄刺槐林土壤团聚体有机碳、全氮、全磷含量的粒径分布呈两边低中间高的趋势。0—20 cm土层,除了50 a土壤团聚体全磷含量最大值出现在1—2 mm粒径中外,其他林地土壤团聚体有机碳、全氮、全磷含量最大值均出现在0.5—1 mm粒径中。20—40 cm土层,各林地土壤团聚体有机碳、全氮、全磷含量最大值多出现在1—2 mm粒径中。各林地0—20 cm和20—40 cm土层小于0.25 mm粒径团聚体的有机碳、全氮、全磷含量均显著低于其他粒径团聚体。
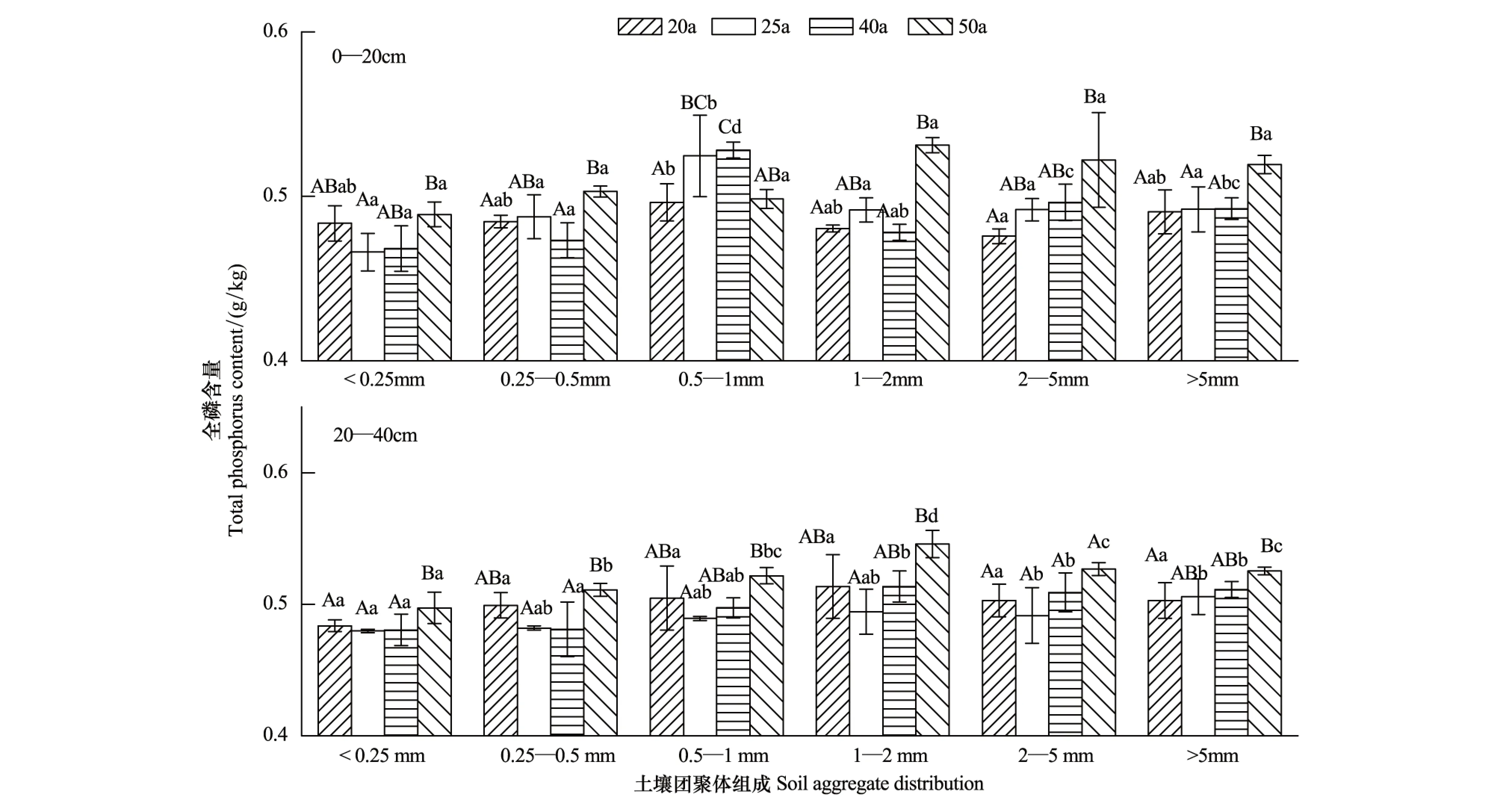
图4 土壤水稳性团聚体中全磷的含量Fig.4 Content of total phosphorus in soil water-stable aggregates
2.2 不同林龄刺槐林土壤团聚体生态化学计量特征
表4所示为不同林龄刺槐林在0—20 cm和20—40 cm土层中土壤团聚体有机碳、全氮、全磷的生态化学计量比。不同林龄刺槐林地土壤团聚体C/N、C/P、N/P变化范围分别在5.84—12.42、1.91—12.00、0.33—0.97,均表现为0—20 cm土层高于20—40 cm土层。
0—20 cm土层,刺槐林土壤团聚体C/N在大于2 mm粒径中随着林龄增加呈显著增大的趋势,在小于2 mm粒径中从大到小的顺序为50 a>25 a>40 a>20 a。在20—40 cm土层中,大于0.25 mm粒径土壤团聚体C/N在20 a显著低于25、40、50 a,其中25、40、50 a之间无显著性差异,小于0.25 mm粒径土壤团聚体C/N从大到小的顺序为50 a>40 a>25 a>20 a。
不同林龄刺槐林同粒径土壤团聚体C/P和N/P的变化趋势与其有机碳和全氮含量的相似。在0—20 cm土层中,土壤团聚体各粒径C/P随着林龄的增加显著增大。20—40 cm土层,土壤团聚体C/P在大于5 mm粒径中从大到小的顺序为50 a>20 a>40 a>25 a,在小于5 mm各粒径中为50 a显著大于25、40 a显著大于20 a,其中25 a和40 a之间无显著差异。
在0—20 cm土层中,20 a的大于2 mm粒径土壤团聚体N/P显著低于其他林地,而25、40、50 a此粒径团聚体N/P之间无显著性差异,土壤团聚体N/P在0.5—1 mm粒径和小于0.25 mm粒径中从大到小的顺序为50 a>40 a>25 a>20 a。20—40 cm土层土壤团聚体N/P的变化趋势为:在大于0.25 mm粒径中土壤团聚体N/P随着林龄的增加呈增大的趋势,其中25、40 a之间无显著性差异,小于0.25 mm粒径团聚体N/P从高到低的顺序为50 a>25 a>40 a>20 a。
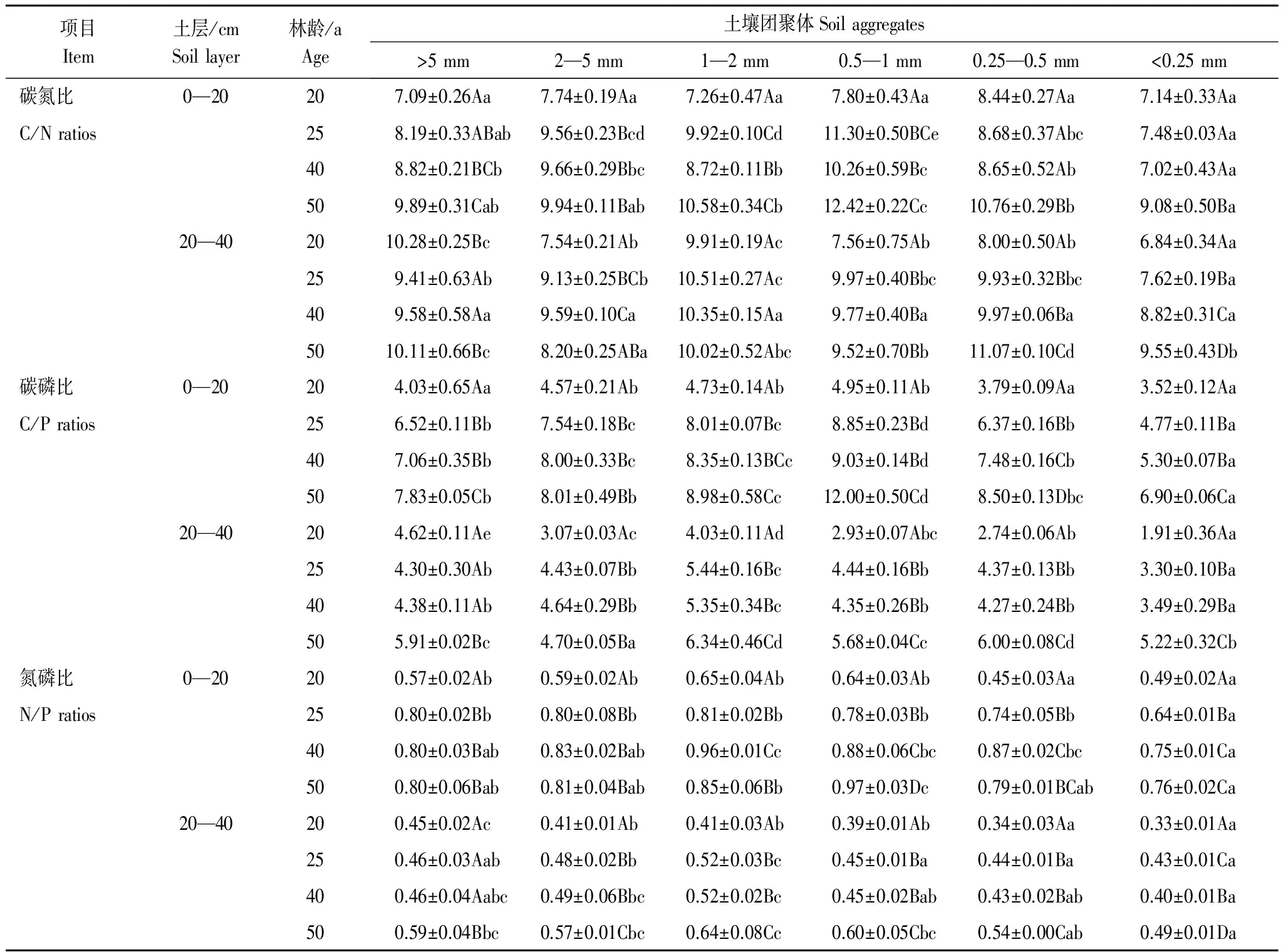
表4 土壤水稳性团聚体有机碳、全氮、全磷的化学计量特征
同一行不同小写字母表示不同粒径团聚体在P<0.05水平上差异显著,同一列不同大写字母表示同一土层不同年限相同粒径团聚体在P<0.05水平上差异显著
相同林龄刺槐林地土壤团聚体C/N、C/P、N/P在0—20 cm土层中最大值多出现在0.5—1 mm粒径中。20—40 cm土层,25、40、50 a土壤团聚体C/N、C/P、N/P最大值出现在0.25—2 mm粒径中,20 a的最大值出现在大于5 mm粒径中。各林地在0—20 cm和20—40 cm土层土壤团聚体C/N、C/P、N/P最小值均出现在小于0.25 mm粒径中。
2.3 不同林龄刺槐林土壤团聚体有机碳、全氮、全磷及其化学计量比的逐步回归分析
如表5所示,对不同林龄刺槐林土壤原土和水稳性团聚体的有机碳、全氮、全磷含量及其C/N、C/P、N/P之间关系进行逐步回归分析。结果表明,土壤原土与水稳性团聚体的有机碳、全氮、全磷含量呈显著正相关:土壤原土有机碳含量与小于0.5 mm粒径和2—5 mm粒径团聚体有机碳含量极显著正相关(P<0.01);土壤原土全氮含量与1—2 mm粒径、小于0.25 mm粒径和大于5 mm粒径团聚体全氮含量极显著正相关;土壤原土全磷含量与2—5 mm粒径团聚体全磷含量显著正相关。
土壤原土C/N受到0.25—0.5 mm粒径和2—5 mm粒径团聚体C/N极显著影响,土壤原土C/P与小于1 mm粒径团聚体C/P极显著正相关,而土壤原土N/P与0.25—1 mm粒径团聚体N/P极显著正相关。
表5 水稳性团聚体有机碳、全氮、全磷及其化学计量比的回归方程
Table 5 The stepwise regression equation of organic carbon, total nitrogen, total phosphorus and its stoichiometric ratios in water-stable aggregates

项目Items回归方程Regressionequation回归系数R2原土和水稳性团聚体有机碳CconcentrationsinbulksoilandWSAsC=0.17CWSA2—5mm+0.53CWSA0.25-0.5mm+0.43CWSA<0.25mm-0.040.99∗∗原土和水稳性团聚体全氮NconcentrationsinbulksoilandWSAsN=0.25NWSA>5mm+0.42NWSA1—2mm+0.34NWSA<0.25mm-0.030.98∗∗原土和水稳性团聚体全磷PconcentrationsinbulksoilandWSAsP=0.45PWSA2—5mm+0.280.60∗原土和水稳性团聚体碳氮比C/NratiosinbulksoilandWSAsC/N=0.32C/NWSA2—5mm+0.67C/NWSA0.25-0.5mm-0.190.99∗∗原土和水稳性团聚体碳磷比C/PratiosinbulksoilandWSAsC/P=0.14C/PWSA0.5—1mm+0.48C/PWSA0.25-0.5mm+0.37C/PWSA<0.25mm+0.490.98∗∗原土和水稳性团聚体氮磷比N/PratiosinbulksoilandWSAsN/P=0.45N/PWSA0.5—1mm+0.39N/PWSA0.25-0.5mm+0.090.97∗∗
*在0.05的显著水平(P<0.05),**0.01的显著水平(P<0.01)
3 讨论
本研究中,不同林龄刺槐林土壤团聚体有机碳、全氮含量随着林龄的增加显著增加,随着土层的加深而降低;不同林龄刺槐林0—20 cm和20—40 cm土层土壤团聚体全磷含量变化较小。林地动植物残体、根系分泌物和微生物残体及其代谢产物等新鲜有机质输入和输出的动态平衡是影响土壤有机质的重要因素[20],其输入量会随着刺槐林龄的增加而增加。因此,随着林龄的增加土壤团聚体有机碳、全氮含量会有显著增加。此外,因土壤表层(0—20 cm)直接接收新鲜有机质的输入,其土壤团聚体有机碳、全氮含量大于20—40 cm土层。土壤全磷含量主要受到土壤母质的影响[21],受有机质输入的影响较小,所以刺槐林土壤团聚体全磷含量变化较小。本研究中,刺槐林土壤团聚体有机碳、全氮的变化范围分别在0.92—5.98 g/kg、0.15—0.48 g/kg之间,低于戴全厚[22]、艾泽明[14]等的研究结果。其原因可能是各林地受到了重度侵蚀的影响,侵蚀造成土壤养分的流失,随着侵蚀的增强,土壤团聚体有机碳、全氮的含量会显著减少[23-24]。
随着林龄的增加,刺槐林地不同粒径土壤团聚体有机碳、全氮、全磷含量的增加幅度有所不同[25]。本研究中,相同林龄刺槐林0—20 cm和20—40 cm土层不同粒径土壤团聚体有机碳、全氮、全磷含量变化大多呈中间高两边低的趋势,最大值出现在0.5—1 mm粒径和1—2 mm粒径中。表明随着刺槐林龄增加,土壤团聚体有机碳、全氮、全磷含量的增加主要体现在大团聚体中。Scott Devine等[26]对3种不同农业生态系统下土壤团聚体有机碳的研究表明:同深度土壤水稳性团聚体有机碳在0.25—2 mm粒径中含量最高。由于大团聚体是由微团聚体和有机质胶结而成[27],故土壤大团聚体富含新鲜有机质并且比微团聚体具有更快的周转时间[28],因此,刺槐林土壤大团聚体有机碳、全氮、全磷含量高于土壤微团聚体。
不同林龄刺槐林土壤团聚体C/N、C/P、N/P变化范围分别在5.84—12.42、1.91—12.00、0.33—0.97之间,低于中国陆地土壤的平均值(中国陆地土壤C/N、C/P、N/P的平均值为12.3、52.7、3.9[29])。本研究结果与黄土丘陵区土壤C/N、C/P、N/P相近,黄土丘陵区森林草原带的土壤C/N、C/P、N/P平均值分别为9.44、5.53、0.75[10]。气候对土壤演变、土壤生物及其与土壤养分的关系起重要的调控作用[30]。陕北黄土丘陵区属于暖温带半干旱气候,土壤有机质含量少,导致土壤团聚体有机碳、全氮含量低,而土壤通过淋溶损失的全磷少,其含量相对较高,结果土壤团聚体C/P、N/P低于中国陆地土壤的平均值。随着林龄的增加,刺槐林0—20 cm和20—40 cm土层土壤团聚体C/N、C/P、N/P显著增加;且刺槐林土壤团聚体化学计量比变化范围随林龄增加从大到小的顺序为C/P大于C/N大于N/P。这说明随着林龄的增加,黄土丘陵区刺槐林土壤团聚体有机碳含量增加速度快,土壤团聚体全氮、全磷含量增加速度缓慢,土壤团聚体全氮和全磷含量可能成为刺槐生长的制约因素。土壤团聚体C/N、C/P能反映有机质腐殖化程度及其对土壤养分的贡献能力,C/N、C/P越高表明有机质腐殖化程度越低,全氮、全磷含量增加越缓慢[31]。土壤N/P可作为植物群落养分限制的指示性指标,相对较低的土壤N/P 说明了氮素对植物群落的制约作用[32]。本研究所得刺槐林土壤团聚体N/P远低于其他地区土壤N/P[33-34]。说明了土壤团聚体全氮含量是黄土丘陵区20—50年刺槐林生长的主要制约因素[35]。
不同粒径团聚体对土壤有机碳、全氮、全磷的保持和供应能力不同,其C/N、C/P、N/P比值在各粒径团聚体中的增加幅度也不同[36]。相同林龄刺槐林0—20 cm和20—40 cm土层土壤团聚体C/N、C/P、N/P最大值多出现在0.25—2 mm粒径中。说明随着刺槐林龄增加,土壤团聚体C/N、C/P、N/P的增加作用主要体现在大团聚体中,尤其是0.25—2 mm粒径团聚体。由于土壤团聚体有机质是土壤微生物生存的来源[37],较大粒径土壤团聚体有机质含量高于微团聚体,其微生物生物量C、N及C/N高于微团聚体[38]。土壤C/N必须比土壤微生物C/N高才能维持土壤微生物C、N含量之间的平衡关系[39]。因此,刺槐林土壤大团聚体C/N、C/P高于微团聚体[40]。随着林龄的增加,刺槐林0.25—2 mm粒径土壤团聚体N/P变化范围最大。说明土壤团聚体全氮含量对刺槐生长的制约作用随着林龄的增加得到了缓解,此缓解作用主要体现在0.25—2 mm粒径土壤大团聚体中。
4 结论
随着林龄的增加,刺槐林各粒径土壤团聚体有机碳、全氮含量及其C/N、C/P、N/P显著增加,均表现为在0—20 cm土层高于20—40 cm土层,而刺槐林土壤团聚体全磷含量变化较小。0—20 cm和20—40 cm土层相同林龄刺槐林土壤团聚体有机碳、全氮、全磷含量及其C/N、C/P、N/P最大值大多出现在0.25—2 mm粒径中。刺槐林0.25—2 mm粒径团聚体对土壤原土有机碳、全氮含量及其C/N、C/P、N/P有显著影响。因此认为,陕北黄土丘陵区营造刺槐林对各粒径土壤团聚体全效养分分配及其平衡关系有积极的影响,此影响作用主要体现在0.25—2 mm粒径土壤大团聚体中,通过影响0.25—2 mm粒径团聚体提高了土壤全效养分的供应和保持能力。另外,对陕北黄土丘陵区刺槐林土壤团聚体N/P研究表明,20—50 a的生长主要受到各粒径土壤团聚体全氮含量的限制,全氮含量是黄土丘陵区刺槐林生长的主要限制因素。
[1] Elser J J, Dobberfuhl D R, MacKay N A, Schampel J H. Organism size, life history, and N: P Stoichiometry. BioScience, 1996, 46(9): 674- 684.
[2] Redfield, A C. The biological control of chemical factors in the environment. American Scientist, 1958, 46(3): 205- 221.
[3] Frost P C, Evans-White M A, Finkel Z V, Jensen T C, Matzek V. Are you what you eat? Physiological constraints on organismal stoichiometry in an elementally imbalanced world. Oikos, 2005, 109 (1): 18- 28.
[4] Moe S J, Stelzer R S, Forman M R, Harpole W S, Daufresne T, Yoshida T. Recent advances in ecological stoichiometry: insights for population and community ecology. Oikos, 2005, 109(1): 29- 39.
[5] Cebrian J. Patterns in the fate of production in plant communities. The American Naturalist, 1999, 154(4): 449- 468.
[6] 高三平, 李俊祥, 徐明策, 陈熙, 戴洁. 天童常绿阔叶林不同演替阶段常见种叶片N、 P化学计量学特征. 生态学报, 2007, 27(3): 947- 952.
[7] 王晶苑, 王绍强, 李纫兰, 闫俊华, 沙丽清, 韩士杰. 中国四种森林类型主要优势植物的C:N:P化学计量学特征. 植物生态学报, 2011, 35(6): 587- 595.
[8] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论. 植物生态学报, 2010, 34(1): 2- 6.
[9] 周鹏, 耿燕, 马文红, 贺金生. 温带草地主要优势植物不同器官间功能性状的关联. 植物生态学报, 2010, 34(1): 7- 16.
[10] 朱秋莲, 邢肖毅, 张宏, 安韶山. 黄土丘陵沟壑区不同植被区土壤生态化学计量特征. 生态学报, 2013, 33(15): 4674- 4682.
[11] Bissonnais Y L E. Aggregate stability and assessment of soil crustability and erodibility: I. theory and methodology. European Journal of Soil Science, 1996, 47(4): 425- 437.
[12] Guggenberger G, Zech W, Thomas R J. Lignin and carbohydrate alteration in particle-size separates of an oxisol under tropical pastures following native savanna. Soil Biology and Biochemistry, 1995, 27(12): 1629- 1638.
[13] 夏建强, 章明奎, 徐建民. 林地开垦后对不同质地红壤碳氮和磷库的影响. 土壤通报, 2005, 36(2): 185- 189.
[14] 艾泽民, 陈云明, 曹扬. 黄土丘陵区不同林龄刺槐人工林碳、 氮储量及分配格局. 应用生态学报, 2014, 25(2): 333- 341.
[15] 张向茹, 马露莎, 陈亚南, 杨佳佳, 安韶山. 黄土高原不同纬度下刺槐林土壤生态化学计量学特征研究. 土壤学报, 2013, 50(4): 818- 825.
[16] 刘毅, 李世清, 李生秀. 黄土高原不同类型土壤团聚体中氮库分布的研究. 中国农业科学, 2007, 40(2): 304- 313.
[17] 吴芳, 陈云明, 于占辉. 黄土高原半干旱区刺槐生长盛期树干液流动态. 植物生态学报, 2010, 34(4): 469- 476.
[18] 中国科学院南京土壤研究所土壤物理研究室. 土壤物理性质测定法. 北京: 科学出版社, 1978: 83- 85.
[19] 鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000: 76- 79.
[20] 车升国, 郭胜利. 黄土塬区小流域深层土壤有机碳变化的影响因素. 环境科学, 2010, 31(5): 1372- 1378.
[21] 刘兴诏, 周国逸, 张德强, 刘世忠, 褚国伟, 闫俊华. 南亚热带森林不同演替阶段植物与土壤中N, P的化学计量特征. 植物生态学报, 2010, 34(1): 64- 71.
[22] 戴全厚, 刘国彬, 薛萐, 余娜, 张超. 侵蚀环境人工刺槐林土壤水稳性团聚体演变及其养分效应. 水土保持通报, 2008, 28(4): 56- 59.
[23] 杨建国, 安韶山, 郑粉莉, 宁南山区植被自然恢复中土壤团聚体特征及其与土壤性质关系. 水土保持学报, 2006, 20(1): 72- 75.
[24] 董雪, 王春燕, 黄丽, 谭文峰, 胡红青. 侵蚀程度对不同粒径团聚体中养分含量和红壤有机质稳定性的影响. 土壤学报, 2013, 50(3): 525- 533.
[25] 王晟强 郑子成 李廷轩. 植茶年限对土壤团聚体氮、 磷、 钾含量变化的影响. 植物营养与肥料学报, 2013, 19(6):1393- 1402.
[26] Devine S, Markewitz D, Hendrix P, Coleman D. Soil Aggregates and Associated Organic Matter under Conventional Tillage, No-Tillage, and Forest Succession after Three Decades. PloS one, 2014, 9(1): e84988.
[27] Elliott E T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Science Society of America Journal, 1986, 50(3): 627- 633.
[28] Puget P, Chenu C, Balesdent J. Dynamics of soil organic matter associated with particle-size fractions of water-stable aggregates. European Journal of Soil Science, 2000, 51(4): 595- 605.
[29] Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C: N: P ratios in China′s soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139- 151.
[30] Vitousek P M. Nutrient Cycling and Limitation: Hawai′i as a Model System. Princeton: Princeton University Press, 2004: 630- 631.
[31] Zhang Z S, Song X L, Lu X G, Xue Z S. Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: influences of vegetation coverage, plant communities, geomorphology, and seawalls. Journal of Soils and Sediments, 2013, 13(6): 1042- 1051.
[32] Cleveland C C, Liptzin D. C: N: P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 2007, 85(3): 235- 252.
[33] 俞月凤, 彭晚霞, 宋同清, 曾馥平, 王克林, 文丽, 范夫静. 喀斯特峰丛洼地不同森林类型植物和土壤C, N, P化学计量特征. 应用生态学报, 2014, 25(4): 947- 954.
[34] 胡启武, 聂兰琴, 郑艳明, 吴琴, 尧波, 郑林. 沙化程度和林龄对湿地松叶片及林下土壤C, N, P化学计量特征影响. 生态学报, 2014, 34(9): 2246- 2255.
[35] 陈亚南, 马露莎, 张向茹, 杨佳佳, 安韶山. 陕西黄土高原刺槐枯落叶生态化学计量学特征. 生态学报, 2014, 34(15): 4412- 4422.
[36] 李玮, 郑子成, 李廷轩. 不同植茶年限土壤团聚体碳氮磷生态化学计量学特征. 应用生态学报, 2015, 26(1): 9- 16.
[37] 徐云蕾, 卢凌霄, 曾馥平, 宋同清, 彭晚霞. 喀斯特峰丛洼地不同森林土壤微生物量碳在团聚体中的分布. 农业现代化研究, 2012, 33(5): 636- 640.
[38] 谢摇涛, 王明慧, 郑阿宝. 苏北沿海不同林龄杨树林土壤活性有机碳特征. 生态学杂志, 2012, 31(1): 51- 58.
[39] Kirkby C A, Kirkegaard J A, Richardson A E, Wade L J, Blanchard C, Batten G. Stable soil organic matter: a comparison of C:N:P:S ratios in Australian and other world soils. Geoderma, 2011, 163(3/4): 197- 208.
[40] An S, Mentler A, Mayer H, Blum W E H. Soil aggregation, aggregate stability, organic carbon and nitrogen in different soil aggregate fractions under forest and shrub vegetation on the Loess Plateau, China. Catena, 2010, 81(3): 226- 233.
Ecological stoichiometry of soil aggregates and relationship with soil nutrients of different-agedRobiniapseudoacaciaforests
SUN Jiao1,2, ZHAO Fazhu3, HAN Xinhui4, YANG Gaihe4, BAI Sunbao5, HAO Wenfang1,*
1CollegeofLifeScience,NorthwestAgricultureandForestUniversity,Yangling712100,China2InstituteofAgriculturalResourcesandEnvironment,NingxiaAcademyofAgricultureandForestrySciences,Yinchuan750002,China3CollegeofUrbanandEnvironmentalScience,NorthwestUniversity,Xi′an710127,China4CollegeofAgriculture,NorthwestAgricultureandForestUniversity,Yangling712100,China5OfficeofReturningFarmlandtoForest,Ansai717400,China
The ecological stoichiometric characteristics of soil aggregates present many interesting structural features linked to soil nutrient evaluation. However, the relationship between the ecological stoichiometry of soil aggregates and the soil nutrients is unclear. In this paper, we review the various aspects of ecological stoichiometric properties of soil aggregates, including those associated with the response of soil chemical parameters and soil structural features to environmental variations as well as the aspects related to the evolution of the forest age. Soil samples were selected among the soil profiles fromRobiniapseudoacaciaforests less than 20, 25, 40, and 50 years old in the Ansai County on the Loess Plateau (located in Shaanxi Province, China). By combining the soil chemical parameters in 0—20 cm and 20—40 cm soil layers, including soil organic carbon (C), total nitrogen (N), and total phosphorus (P), we aimed to determine the relationship of the ecological stoichiometric characteristics of soil aggregates and the soil nutrient content through stepwise multiple regression analysis. The results showed that the stoichiometric properties were limited by N content in soil aggregates from all stand ages ofRobiniapseudoacaciaforests. The C and N contents in soil aggregates and the C, N, and P stoichiometric ratios increased with increasing forest age. The C content ranged from 0.92 g/kg to 5.98 g/kg; the N content ranged from 0.15 g/kg to 0.48 g/kg; and the C/N, C/P, and N/P (C, N, and P stoichiometric ratios) ranged from 5.84 to 12.42, 1.91 to 12.00, and 0.33 to 0.97, respectively, inRobiniapseudoacaciaforest stands of different ages. The C and N contents in soil aggregates and their C, N, P stoichiometric ratios in the 0—20 cm soil layer were higher than those in the 20—40 cm soil layer. However, the content of P did not vary significantly in both soil layers, ranging from 0.47 g/kg to 0.55 g/kg in soils from the forests with different age. The highest C, N, and P contents and corresponding stoichiometric ratios in soil aggregates of the two soil layers were found in aggregate particles with sizes ranging from 0.25 mm to 2 mm; the aggregates within this size range had a significant effect on soil C, N, and P contents and their stoichiometric ratios. In summary, the total nutrient contents in soil aggregates fromRobiniapseudoacaciaforest and their balance was significantly affected by the age of forest stands in the loess hilly regions, and it was mainly reflected in the aggregate particles that were 0.25—2 mm in size. Thus,Robiniapseudoacaciaforest improved the capacity of soil nutrient conservation and supply by affecting the total nutrient contents of these aggregate particles and their balance.
soil water-stable aggregate;Robiniapseudoacaciaforest; ecological stoichiometry characteristics
国家林业部公益性行业科研专项资助项目(201304312);国家自然基金资助项目(41301601)
2015- 04- 25;
日期:2016- 03- 03
10.5846/stxb201504250852
*通讯作者Corresponding author.E-mail: haowenfang@nwsuaf.edu.cn
孙娇,赵发珠,韩新辉,杨改河,白孙宝,郝文芳.不同林龄刺槐林土壤团聚体化学计量特征及其与土壤养分的关系.生态学报,2016,36(21):6879- 6888.
Sun J, Zhao F Z, Han X H, Yang G H, Bai S B, Hao W F.Ecological stoichiometry of soil aggregates and relationship with soil nutrients of different-agedRobiniapseudoacaciaforests.Acta Ecologica Sinica,2016,36(21):6879- 6888.
