干旱胁迫下AMF对云南蓝果树幼苗生长和光合特征的影响
2016-12-28张珊珊康洪梅杨文忠向振勇
张珊珊,康洪梅,杨文忠,向振勇
云南省林业科学院珍稀濒特森林植物保护和繁育国家林业局重点实验室, 昆明 650201
干旱胁迫下AMF对云南蓝果树幼苗生长和光合特征的影响
张珊珊,康洪梅,杨文忠*,向振勇
云南省林业科学院珍稀濒特森林植物保护和繁育国家林业局重点实验室, 昆明 650201
采用盆栽试验与称重控水法,将土壤相对含水量分别控制在田间最大持水量的100%、91.68%、82.85%、60.00%、41.86%和21.28%,并在这6个不同的土壤相对含水量条件下,分别设添加苯菌灵(杀真菌剂)(低AMF)和不添加苯菌灵(高AMF)处理,研究干旱胁迫下AMF对极小种群野生植物云南蓝果树幼苗生长和光合特征的影响,揭示云南蓝果树濒危的微生物学机制,为云南蓝果树保护措施的制定与实施奠定基础。结果表明,添加苯菌灵处理显著降低了不同水分条件下的AMF侵染率,说明试验中AMF处理的实生苗在生长和光合特征上的差异是苯菌灵处理下侵染率下降导致的;随着干旱胁迫的加剧,云南蓝果树幼苗的根部AMF侵染率显著降低、叶面积等生长指标和净光合速率(Pn)等光合参数都发生显著变化;高AMF处理可以显著增加水分充足和轻度干旱胁迫条件下云南蓝果树幼苗的大部分生长指标和光合参数,而对重度胁迫下的云南蓝果树幼苗没有显著影响,说明重度干旱胁迫对其影响大于AMF的影响;另外,整合了可塑性指数分析和隶属函数分析两种方法对其抗旱性进行评价,云南蓝果树幼苗基本上无法通过调节形态和光合能力来适应水分环境的变化,但是高AMF处理可使云南蓝果树幼苗具有较强的可塑性和更强的抗旱性。实验结果为云南蓝果树的科学保育及种苗繁育提供了理论依据。
丛枝菌根真菌;干旱胁迫;生长指标;光合特征;云南蓝果树
云南蓝果树(NyssayunnanensisW.C.Yin)为蓝果树科(Nyssaceae)蓝果树属(NyssaGronov ex Linn.)植物,国级Ⅰ级重点保护野生植物,云南特有种和极危种[1]。云南蓝果树天然林内幼苗更新极其困难[1],但对云南蓝果树的研究仅见于系统分类、形态修订及种子萌发特性等方面的研究[2- 3],未见相关报道。
更新幼苗与降水特点密切相关。关于木本植物的研究发现,干旱期的长短很大程度上影响了幼苗的生长和存活,连续的降水过程或连阴雨天气才是更新幼苗发生的有利条件[4- 5]。另外,土壤水分是幼苗生长存活的关键因子,在光照强度大的森林中,由于蒸发量大,土壤含水量较低,森林内的幼苗在冬季干旱时容易大量死亡[6]。而云南蓝果树的原生境所在地西双版纳地区38 a(1974—2003年)来平均的气候情况为:从9 月份至次年2 月份,西双版纳均处于旱季[7- 8]。而云南蓝果树种子一般于每年的8 月份之前就基本成熟,错过了种子萌发的最适季节。这说明干旱已经严重影响了云南蓝果树的种群延续和扩大。然而,干旱胁迫究竟对云南蓝果树幼苗有哪些影响?云南蓝果树能否表现出相应的表型可塑性?影响这种表型可塑性的因素又有哪些?这都需进一步研究。
丛枝菌根真菌(arbuscular mycorrhizal fungi,AMF)能改善植物养分吸收[9]、植株生长状况[10- 11]等。研究表明,丛枝菌根能显著促进植物的生长[12]、提高植物养分吸收的能力[13]、增强植物的抗逆性(例如:抗干旱、耐瘠薄和耐重金属污染等)和抗病性[14]。何跃军等观察到接种AM真菌的构树(Broussonetiapapyrifera)生长量及净光合速率都显著高于未接种的对照株[15]。虽然,目前还未见AMF与云南蓝果树共生关系的相关报道,但是笔者在对云南蓝果树幼苗的根进行解剖时发现其根部可被AMF侵染,且侵染率较高(45.66%)(文章未发表)。那么,干旱胁迫条件下AMF对云南蓝果树的影响如何,以及菌根共生体功能的发挥与程度又如何呢?
本试验选用苯菌灵为杀真菌剂,用盆栽方法人工模拟土壤干旱条件,研究正常供水和干旱胁迫条件下AMF对云南蓝果树幼苗生长和光合特征的影响,进而揭示云南蓝果树濒危原因,为其科学保育及种苗繁育提供理论依据。苯菌灵能很好地抑制AMF的活性,降低其与植物形成共生体的机会,抑制丛枝菌根的形成。
1 材料和方法
1.1 材料
选择云南蓝果树1年实生幼苗为研究对象。2013年4月中旬将生长基本一致的幼苗移栽到容积为10 L的花盆中,每盆1株,栽培基质为云南省林业科学院苗圃红壤。试验用红壤于次氯酸钠(NaClO)中浸泡48 h,后洗净后120℃烘8 h。
1.2 试验设计
试验于云南省林业科学院温室条件下进行,自然温度和自然光照,水分人工控制,此温室主要种兰花用,需对其进行定期加湿,因此整个实验过程中的空气湿度较高。试验为2因子(AMF×水分)试验。AMF处理为施加苯菌灵(低AMF)和不加苯菌灵(高AMF),对于施加苯菌灵处理,将2 g杀真菌剂苯菌灵溶于2 L自来水,加到盆钵中,每月处理1次,获得低AMF处理的土壤,同时,在对照处理的盆钵中每次均加入相同量的自来水。水分设置6个水平,共得到2(AMF)×6(水分)=12个处理,每个处理10个重复,共有120个盆钵。正式实验前,测得栽培用土的田间持水能力(WHC),此时的实际含水率为32.32%(干土重基础)。实验处理以相对土壤含水量(RWC),即各盆的土壤含水率占WHC的百分比来区分(表1):第1个处理始终保持饱和,每天补足由于蒸散所散失的水分(100%);第2个处理保持在90%—100%(91.68%),当由于蒸散所散失的水分达到90%时,立即补充水分,使之达到饱和;以同样的方法保持第3个处理在80%—100%(82.85%);第4个处理在60%—100%(60.00%);第5个处理在40%—100%(41.86%);第6个处理在20%—100%(21.28%)。文中分别用W1,W2,W3,W4,W5和W6表示6个处理。浇水时用缓慢滴加的方法,尽量避免水分从花盆底部流失,这样可以最大限度地保证土壤养分不损失。浇水在每天下午进行,天气炎热时,W1处理每天中午补水100—200 mL。实验期限为6个月,2014年10月中旬采样,测定相应指标。
1.3 测定指标及方法
1.3.1 AMF侵染率的测定
取部分新鲜根部样品固定于FAA溶液中(37%甲醛-冰醋酸-50%乙醇溶液,体积比 9∶0.5∶0.5)用于检测AMF侵染率。先将根部的固定液清洗干净,然后浸泡在10%的KOH中,90℃水浴加热5 min,然后用1%的盐酸酸化15 min,并用酸性品红染色过夜,将根部剪成 2 cm长的根段,在显微镜下10倍物镜观察,用十字交叉法计算侵染率(gridline intersect method)(Giovannetti & Mosse,1980)。侵染率的计算公式为:侵染率= 侵染根段长度/根段总长度×100%。
1.3.2 生长指标测定
实验结束后,用直尺测量株高,用数显游标卡尺测量叶长和叶宽,并记录叶片数目。用CI- 203 型便携式激光叶面积仪(CID Inc, Wash ington, USA)测定野外叶片的面积,得到叶面积与叶长和叶宽的拟合公式:叶面积= 0.7416×叶长×叶宽-0.7477 (R2= 0.984,P< 0.001,n=120),形态测量的叶面积通过拟合公式得出。采用King 等人的方法计算株高的相对生长速率(RGR) =(lnH2-lnH1)/Δt,式中:H1和H2为前后两次测量时的株高,Δt为测量间隔时间。
将幼苗整棵挖出清洗干净,分为地上部分和地下部分,105℃杀青30 min后在80℃下烘干至恒重。称量各部分干重并计算如下参数:地上部分生物量比(AMR,地上部分重/总生物量)、地下部分生物量比(BMR,地下部分重/总生物量)、根冠比(R/S,地下部分生物量/地上生物量),结合形态测量的总叶面积计算叶面积比率(LAR,总叶面积/总生物量)和比叶面积(SLA,总叶面积/叶重)。
1.3.3 气体交换和资源利用参数测定
实验中后期用用LI- 6400XT便携式光合仪于09:00—11:00测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)等参数, 并记录光合有效辐射(PAR)和大气CO2浓度(Ca)等环境参数, 并计算气孔限制值(Ls) = 1-Ci/Ca、瞬时水分利用效率(WUE) =Pn/Tr。
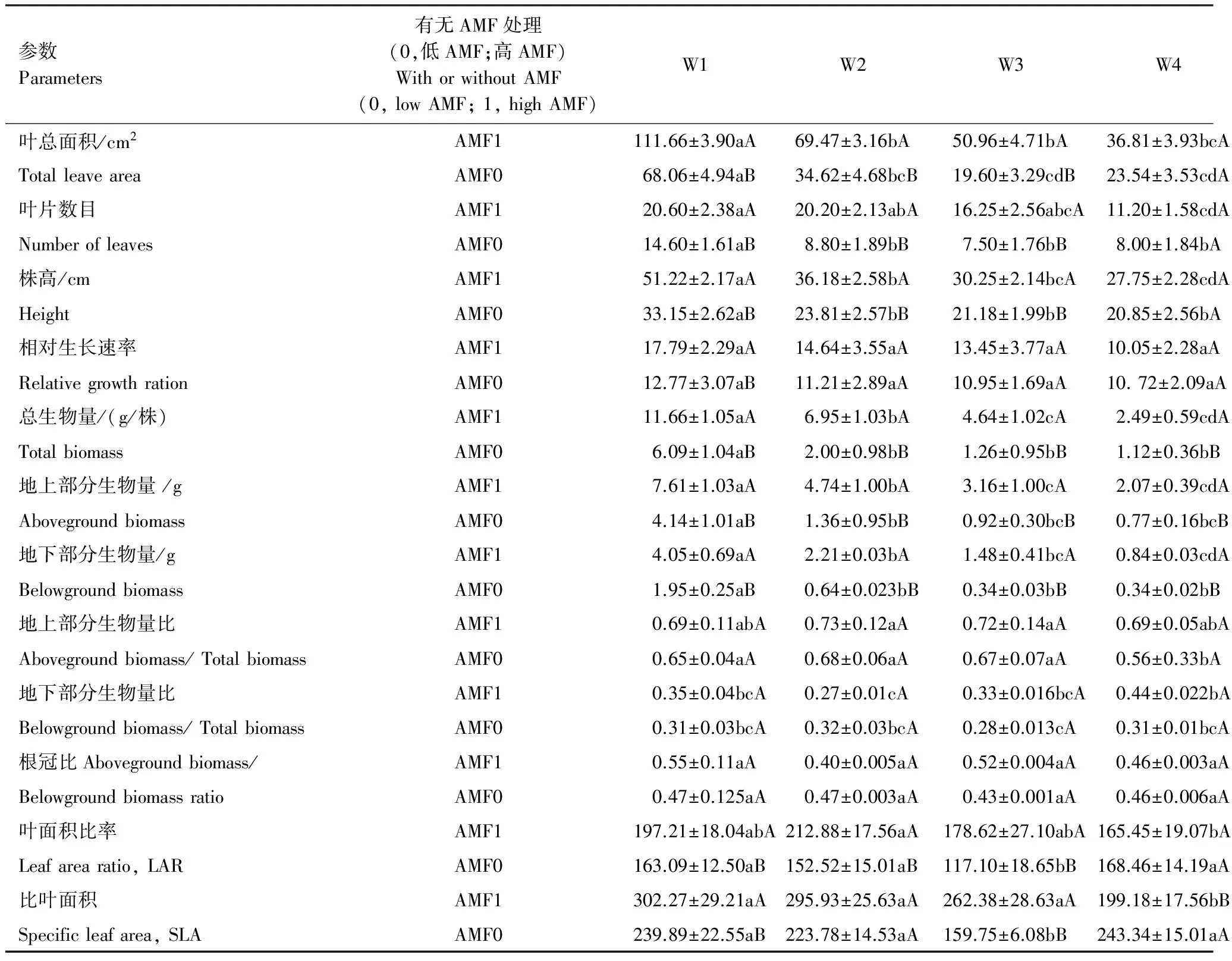
表1 不同水分条件下AMF对云南蓝果树幼苗生长指标的影响(±标准误)
表中同行不同小写字母表示不同水分条件下叶片的解剖结构指标在P<0.05水平差异显著;同列不同大写字母表示不同AMF处理下叶片的解剖结构指标在P<0.05水平差异显著
1.4 数据统计分析
试验采用SPSS16.0对数据进行单因素方差分析、Duncan多重比较、变量间的相关性分析以及主成分分析。方差分析时,不满足方差齐性检验的数据通过[arcsin]或[log(x+1)]转换以满足方差分析的要求。通常采用Post-hoc Tukey方法检验变量的显著性,如果数据不满足参数检验的条件,采用Kruskall-Wallis方法检验。5%为显著水平,1%为极显著水平。根据Valladares等的方法计算各参数的可塑性指数, 即6个水分梯度下某变量的最大值减去最小值再除以最大值。其中隶属函数值具体公式为:如果指标与抗旱性成正相关,X(μ)=(X-Xmin)/(Xmax-Xmin);如果指标与抗旱性成负相关,X(μ)=1-(X-Xmin)/(Xmax-Xmin)。式中,X表示各指标的测定值,Xmax和Xmin分别表示各处理条件下指标的最大和最小测定值。
2 结果与分析
2.1 丛枝菌根真菌侵染率
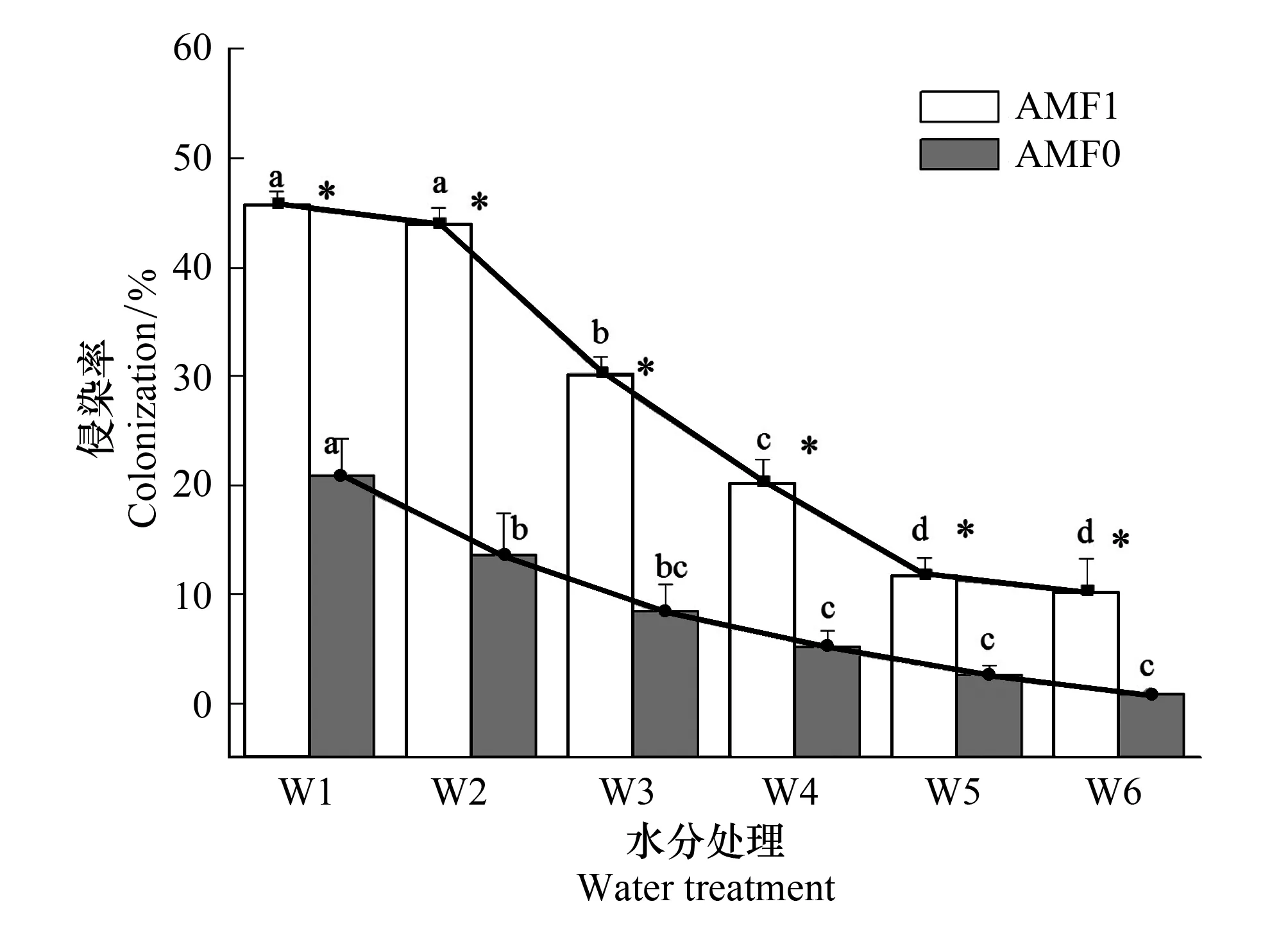
图1 不同处理条件下云南蓝果树的丛枝菌根侵染率 Fig.1 Arbuscular mycorrhizal root colonization rate under different treatments
图1显示,施加苯菌灵处理显著降低了AMF对云南蓝果树的侵染率,从而形成低AMF处理(F=38.141,P<0.01),而不施加苯菌灵的处理则形成高AMF处理。不管是高AMF处理还是低AMF处理,AMF侵染率都随着土壤水分含量的降低显著下降(F=11.093,P<0.01;F=37.175,P<0.01)。
2.2 不同水分条件下丛枝菌根真菌对云南蓝果树幼苗生长的影响
表1表明,随着土壤相对含水量的降低,不管是AMF侵染率高还是低,云南蓝果树生长指标中除了相对生长速率和根冠比2个指标上没有发生显著变化,叶片数目、叶面积、株高、总生物量、地下部分生物量、地上部分生物量、LAR和SLA8个指标呈递减状态。生物量的分配方式也发生了显著的变化(P<0.05),地上部分生物量比和地下部分生物量比都呈现先升高后降低的趋势。
在测定的10个生长指标中,AMF处理的影响也不同(表1)。W1和W2条件下AMF处理对9个生长指标有显著影响;W3条件下AMF处理对8个生长指标有显著影响;W4条件下AMF处理对4个生长指标有显著影响;W5条件下AMF处理对5个生长指标有显著影响;W6条件下AMF处理只对1个生长指标有显著影响。因此,AMF处理从土壤含水量处理为W1时便开始显著影响生长指标的特征值。当土壤过于干旱(W6)时,AMF处理对其幼苗生长的影响并不显著。
2.3 不同水分条件下丛枝菌根真菌对云南蓝果树幼苗光合特征的影响
由表2看出,随着土壤相对含水量的降低,不管是AMF侵染率高还是低,云南蓝果树光合特征指标中Pn、Tr和Gs3个指标呈显著递减状态。Ci表现为先降后升的趋势,Ls和WUE则为先升后降的趋势。
在测定的10个生长指标中,AMF处理的影响也不同(表2)。W1条件下AMF处理对4个光合特征有显著影响;W2、W3、W5和W6条件下AMF处理对3个光合特征有显著影响。W4条件下AMF处理对4个光合特征有显著影响;
2.4 云南蓝果树幼苗生长指标和光合特征间的相关性分析
由表3相关分析可以看出:AMF侵染率与叶片数目、叶面积、株高和总生物量等12个指标呈显著正相关;与其他指标间虽然存在一定的或正或负的相关性,但均未达到显著水平(表3)。土壤相对含水量与叶片数目、叶面积和总生物量12个指标呈显著正相关。其余18个指标之间存在56个极显著正相关,12个显著正相关,5个极显著负相关,3个显著负相关。
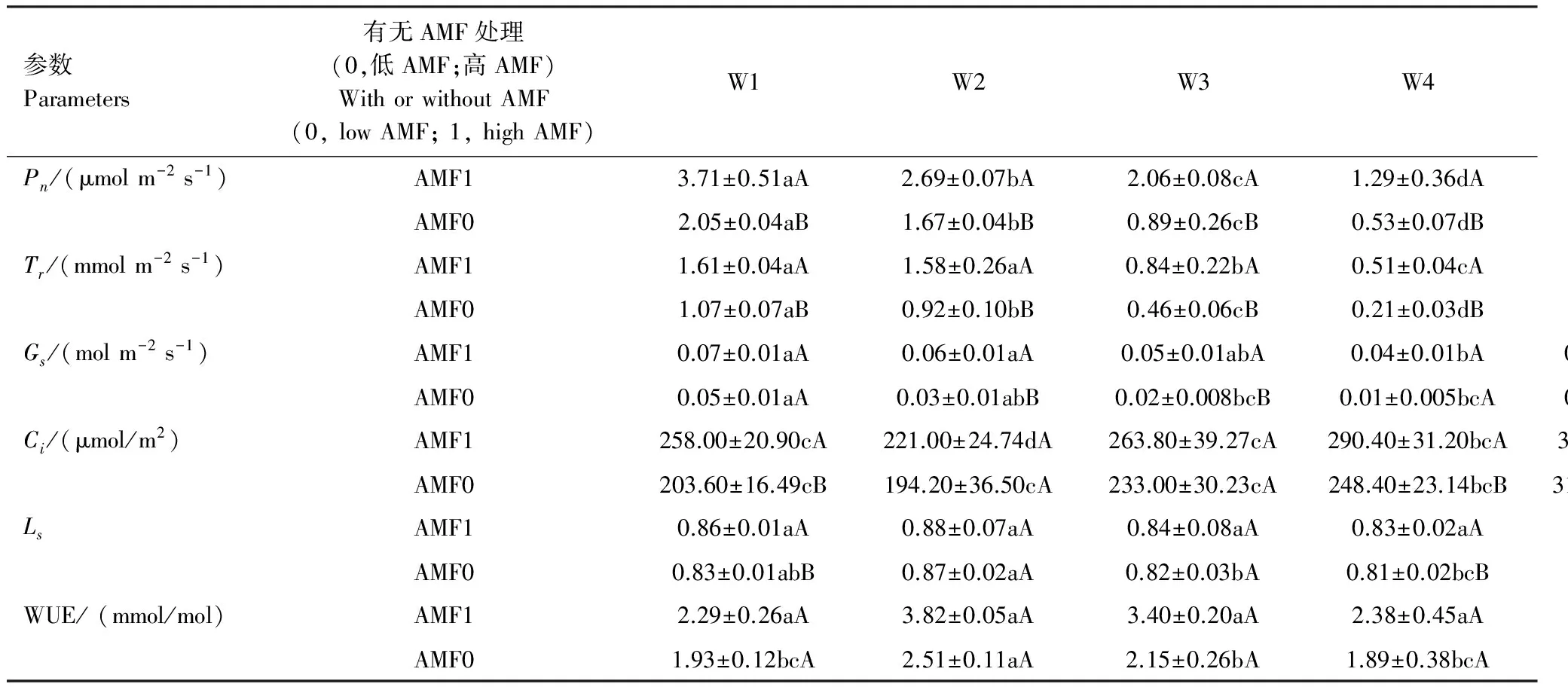
表2 干旱条件下AMF对云南蓝果树幼苗光合特征的影响(±标准误)
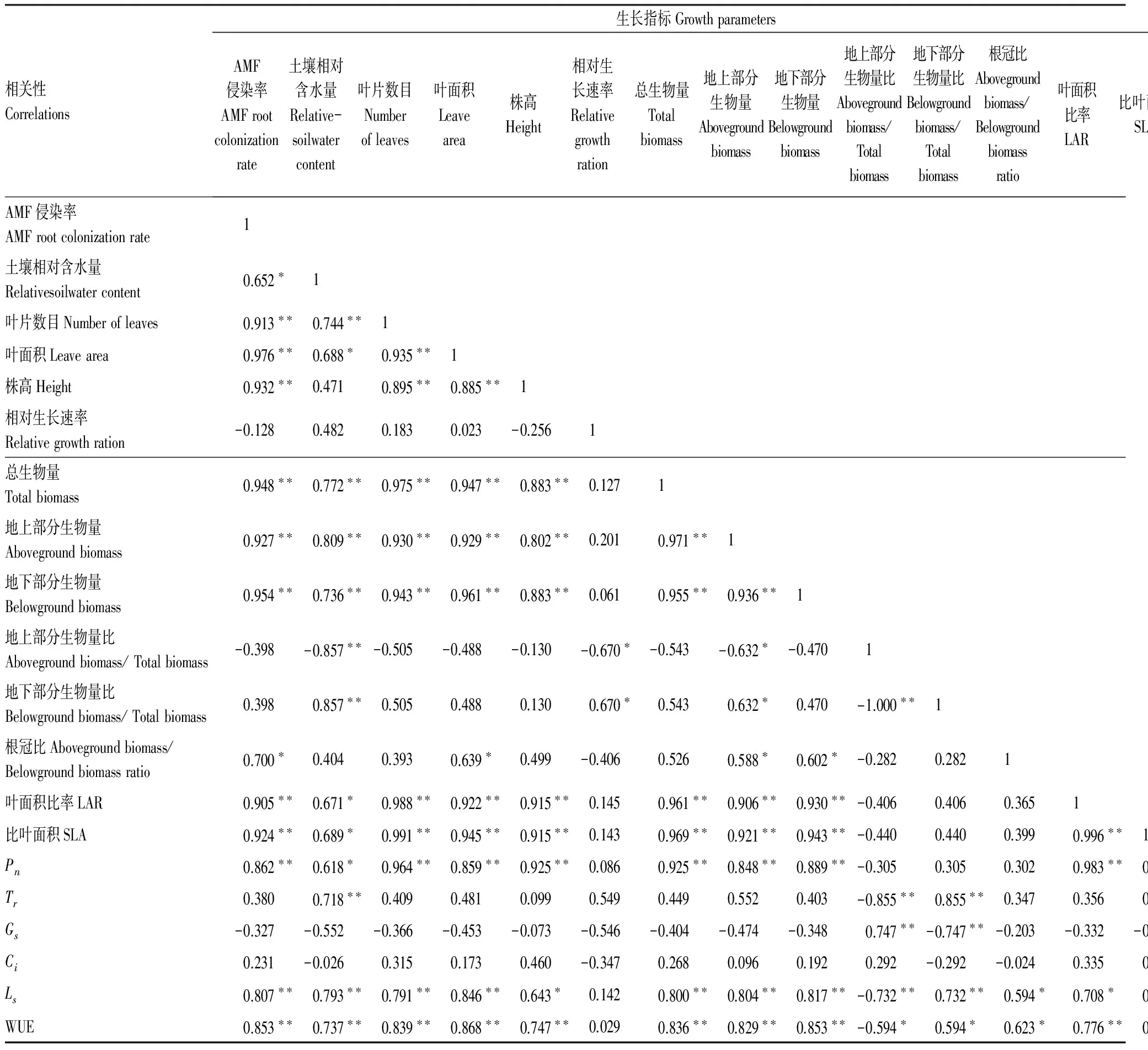
表3 干旱胁迫下云南蓝果树不同参数之间的相关性分析
*:0.05水平上相关性显著(双侧检验); **:0.001水平上相关性显著(双侧检验)
2.5 云南蓝果树幼苗生长指标和光合特征间的主成分分析
本文采用主成分分析法对18项指标进行分析,并根据各主成分中每个指标载荷量及其变异系数的大小筛选出具有代表性的指标。
从基于相关矩阵的主成分分析结果(表4和表5)可以看出,低AMF处理条件下,云南蓝果树幼苗第2个主成分的特征值为3.176,累计贡献率为87.771%,说明干旱胁迫下低AMF对云南蓝果树的前2个主因子基本上能概括18个变量的主要信息,所以共提取了2个主成分因子。在确定了主成分因子数量后,在第一主成分中,地下部分生物量、株高和总生物量7个指标得分较高,均为0.90以上;在第二主成分中,特征向量系数按照绝对值大小依次排列,得分最高的地上部分生物量比和地下部分生物量比,分别是0.908和-0.908。
从基于相关矩阵的主成分分析结果(表4和表5)可以看出,高AMF处理条件下,云南蓝果树幼苗第2个主成分的特征值为2.725,累计贡献率为93.327%,说明干旱胁迫下高AMF对云南蓝果树的前2个主因子基本上能概括18个变量的主要信息。在第一、二主成分中,将2个主成分的特征向量系数按照绝对值大小依次排列,第一主成分居得分较高的的为地下部分生物量、株高和总生物量等7个指标,得分均为0.90以上,综合反映了云南蓝果树幼苗的生长情况;第二主成分得分为0.90以上的为WUE,分值为0.931,主要反映了叶片的水分利用效率。
2.6 不同水分条件下丛枝菌根真菌对云南蓝果树幼苗光抗旱性的影响
生长指标方面,不管是高AMF还是低AMF条件下,总生物量、地上部分生物量、地下部分生物量、叶片数目,Pn、Tr和Gs具有较高的可塑性指数(表1,表2);高AMF条件下,云南蓝果树幼苗各个参数及除Ls之外的其余5个光和特征指标都表现出较高的可塑性指数(表1,表2)。通过对上述18个指标的综合分析,不同处理下云南蓝果树幼苗抗旱性的隶属函数结果列于表6。从表6可以看出,土壤水分含量在W1—W6范围时,高AMF处理都增加了云南蓝果树幼苗的隶属函数值,隶属函数值分别为0.55、0.68、0.54、0.45、0.43和0.44,增强了其在各水分条件下的抗旱性。
3 讨论
3.1 不同水分条件对云南蓝果树幼苗生长和光合特征的影响
本试验中采用苯菌灵灭菌的方法来抑制AMF对云南蓝果树实生苗根系的侵染,得到理想的结果,即杀真菌剂处理显著降低了云南蓝果树幼苗根侵染率,使得本试验中AMF处理的实生苗在生长和光合特征上的差异可以用苯菌灵处理导致的侵染差异加以解释[16]。
干旱胁迫会限制植物的生长[17]。本研究表明,干旱胁迫显著抑制了云南蓝果树幼苗的10个生长指标。土壤相对含水量较高时,幼苗具有较多的叶片和较大的叶面积,从而拥有较大的光合速率和较大的生物量。随着干旱胁迫程度的增加,云南蓝果树幼苗的大部分生长指标都发生了显著的变化,如叶片数目、叶面积和株高等8个指标都显著降低,这与有些研究发现的轻度胁迫条件下某些植物可以通过形态的可塑性作出适应相反,证明云南蓝果树缺乏应对干旱胁迫的策略;生物量分配方式也发生了显著变化,水分充足时,云南蓝果树将生物量较多地分配到地上部分的生长上,可以提高光合产量,满足植物的需要;而在干旱胁迫时,大多数植物会降低地上部分生物量的分配,将更多的资源分配到地下,从而获得更多的水分和营养,获得更多的地下部分生物量,根冠比呈现先升高后降低的趋势[17- 18],然而,云南蓝果树却随着干旱胁迫的增加,根冠比没有发生显著的变化,这一定程度上揭示了云南蓝果树根部表型可塑性差的特点;生物量分配方式也发生了紊乱,例如地上部分生物量比和地下部分生物量比都呈现先升高后降低的趋势;LAR 和SLA 是调控植物功能的重要性状[19],水分充足时,云南蓝果树具有较大的LAR 和SLA,从而保证了较高的光资源捕获面积,两者共同保证了较高的生物量积累[17]。干旱胁迫导致SLA的减少是适应干旱的一种表现[20- 21],因此云南蓝果树的SLA表现出较强的可塑性。
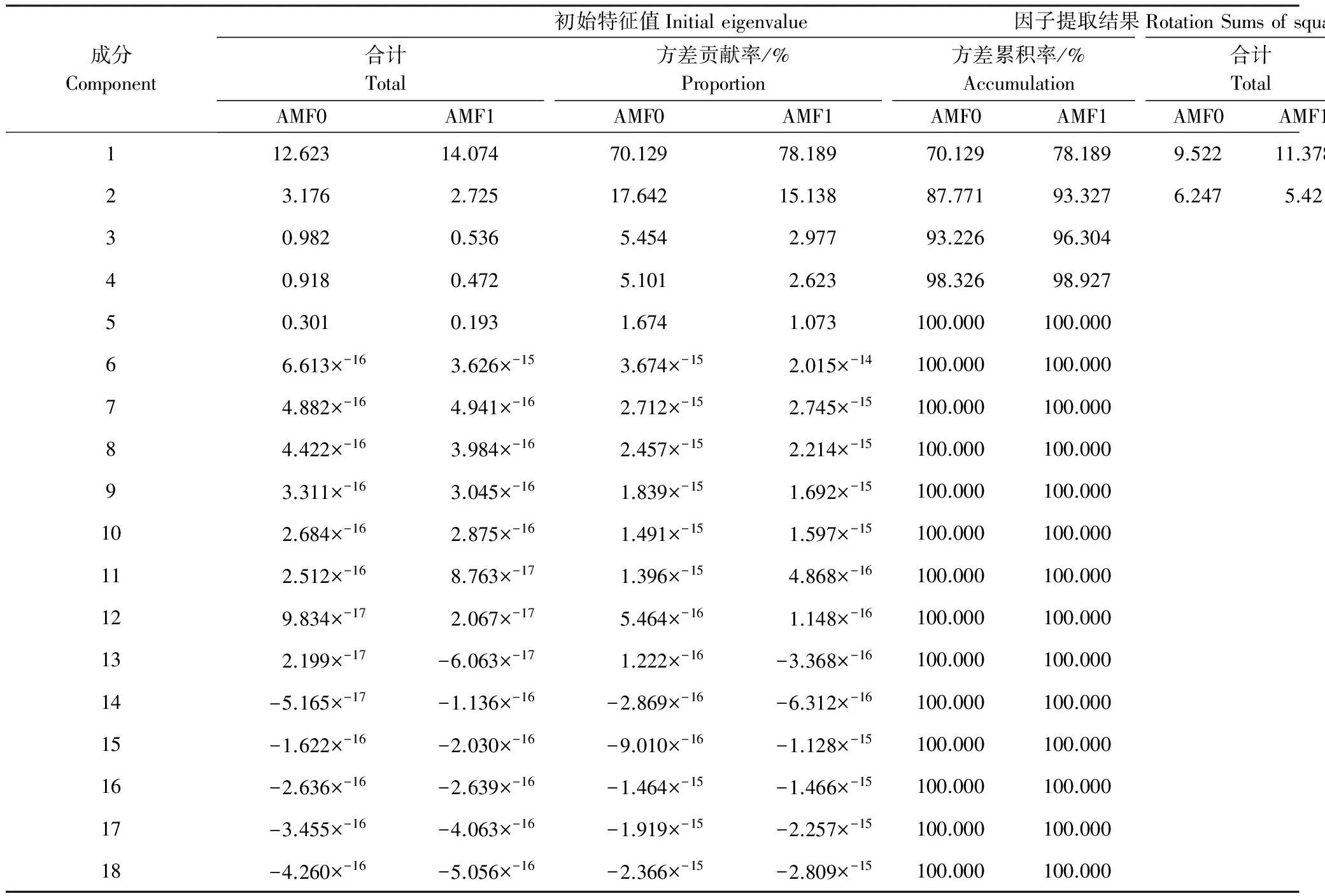
表4 不同AMF处理条件下的主成分分析表
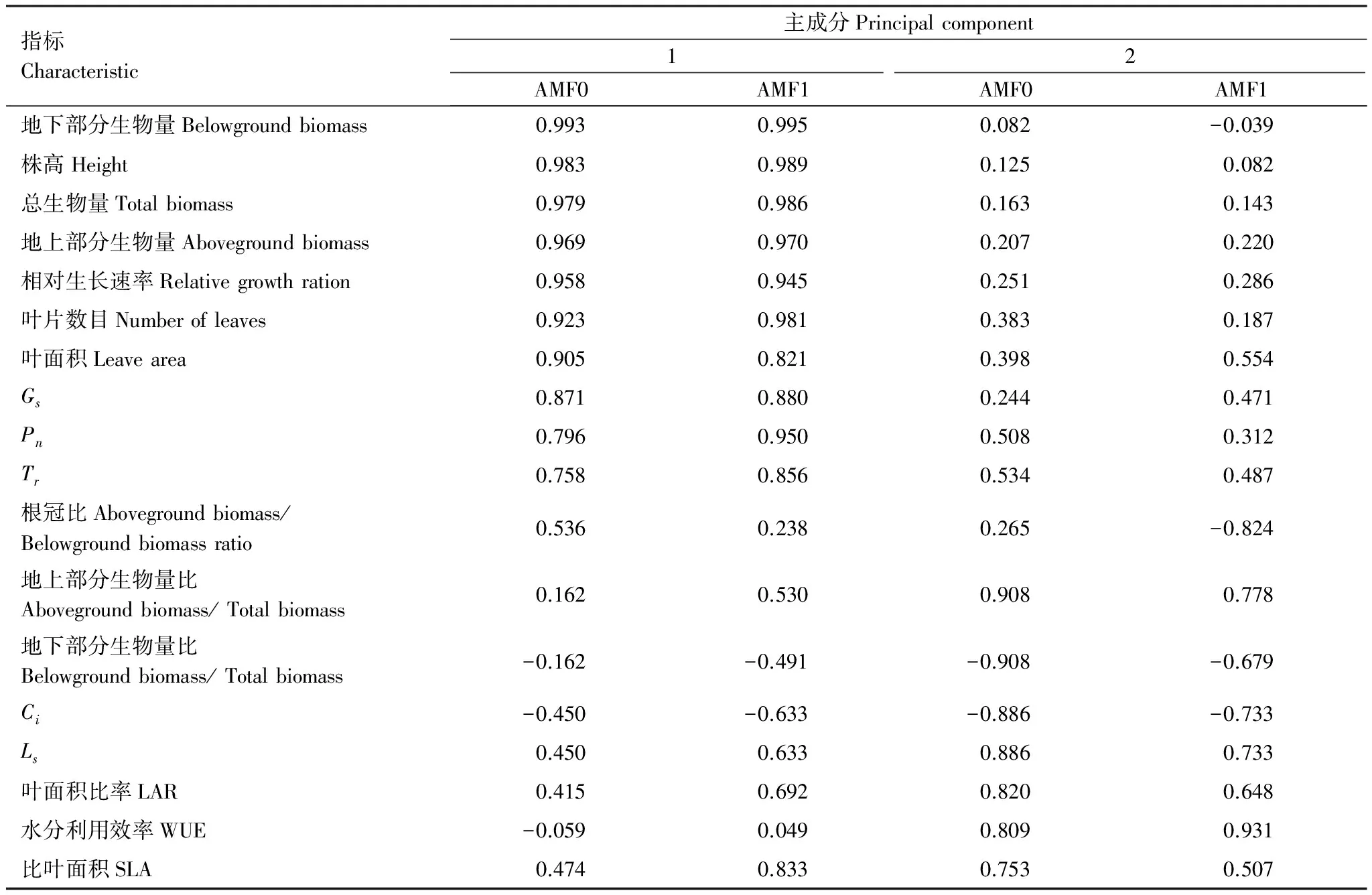
表5 不同AMF处理条件下各成分得分系数矩阵
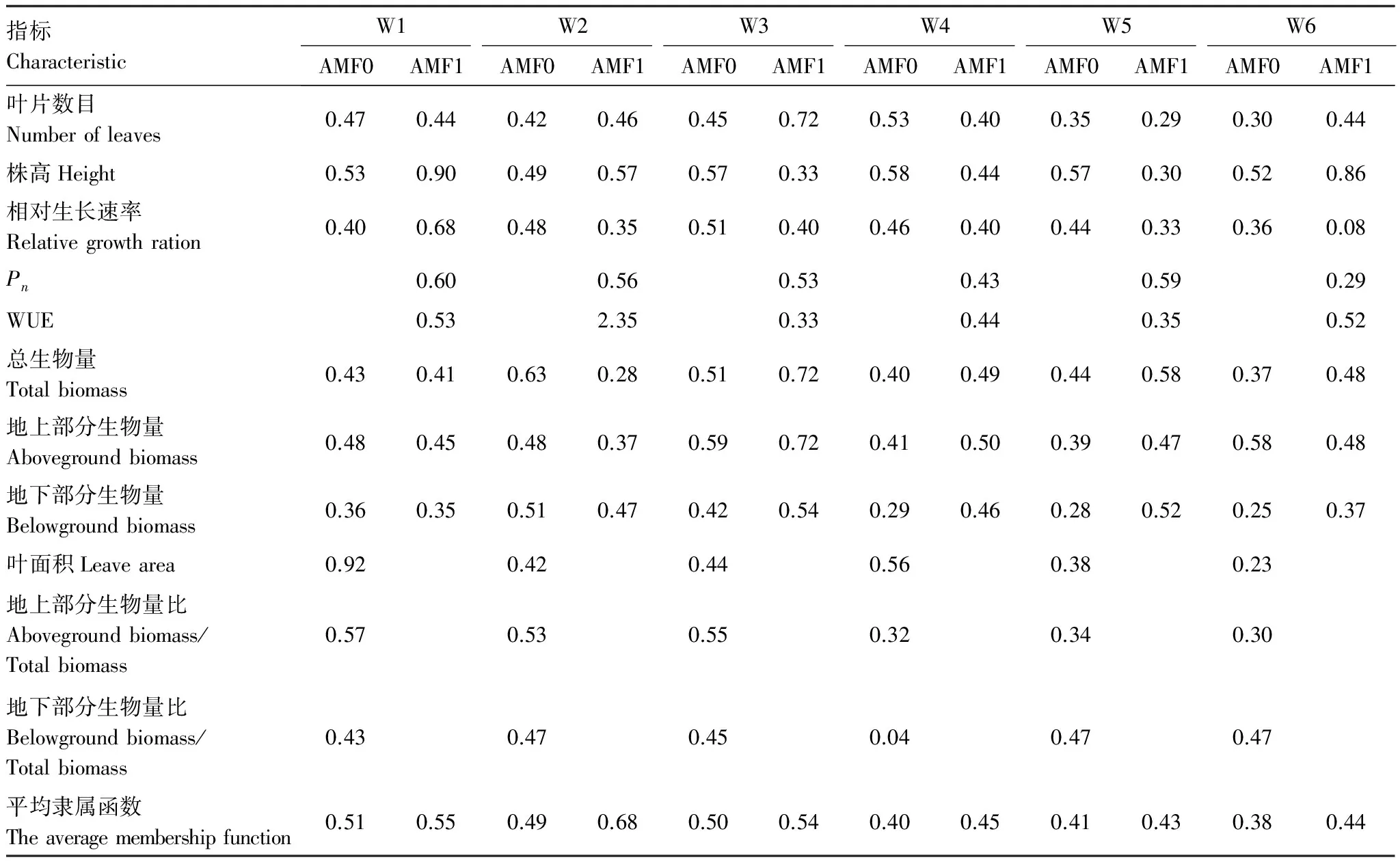
表6 云南蓝果树抗旱性综合评分值
干旱影响植物的生长在很大程度上要归结于光合作用的响应,而光合作用对缺水特别敏感[17, 22- 24]。付士磊等在研究杨树的光合作用与抗早能力的关系中指出,在一定时间内,随胁迫程度的增加,叶片的净光合速率显著降低[25]。气孔可以快速感应大气湿度或根系水势的变化,通过降低Gs可以有效防止水分散失。受旱植物为了尽量维持其Pn,叶片Gs会降低。但是,随着水分胁迫程度的增强,Ci和CO2浓度达到平衡,出现先降低再升高的趋势,Ls和WUE则为先升后降[26]。本研究中,云南蓝果树的Pn、Tr和Gs3个指标随着干旱程度的增加呈显著递减状态,Ls和WUE则为先升后降的趋势。由此表明,干旱胁迫条件下云南蓝果树幼苗不仅能获得的资源量较少,而且光合机构的伤害造成了资源利用率的低下,共同影响了云南蓝果树幼苗的生长。
3.2 不同水分条件下丛枝菌根真菌对云南蓝果树幼苗生长和光合特征的影响
干旱胁迫条件下AM真菌对植物生长的影响已有一些研究[22, 27- 30]。陈冬青等通过施加苯菌灵对云南蓝果树进行AMF处理,发现水分胁迫下AMF对黄顶菊的生物量和生理指标影响显著,AMF共生能够促进黄顶菊土壤水分和矿质营养吸收,改变植物代谢活动,提高抗旱性[16]。贺学礼和赵丽莉发现接种摩西球囊霉(Glomusmoseae)显著促进了小麦(Triticumaestivum)的营养生长和磷吸收, 提高了叶绿素含量, 增强了光合作用[29]。贺学礼和李生秀采用土培试验研究了水分胁迫条件下接种摩西球囊霉对玉米(Zeamays)营养生长和抗旱性的影响[31]。贺学礼等发现,不同水分条件下,接种AMF提高了民勤绢蒿的生长和抗旱性[32]。本试验中丛枝菌根的处理对干旱胁迫条件下云南蓝果树幼苗的生长产生了较大的影响,AMF侵染率与8个生长指标呈显著正相关关系。在胁迫前期,AMF对云南蓝果树幼苗的作用明显,但是到了胁迫后期,干旱胁迫对其影响大于AMF的影响,导致AMF对重度干旱胁迫条件下的云南蓝果树幼苗没有作用。
AMF可以提高光合作用效率,增强宿主植物的抗旱性[22, 27- 28]。很多研究发现AMF可以提高很多植株叶片的光合速率(Pn)、蒸腾速率(Tr)、增加了气孔导度(Gs)和水分利用效率(WUE), 水分胁迫下促进作用更加显著[33]。Morte等在向日葵上也得出接种菌根能提高蒸腾速率、气孔导度和净光合速率等结论[34]。本试验中丛枝菌根的处理对云南蓝果树幼苗的光合特征产生了较大的影响,AMF处理能够提高云南蓝果树幼苗的光合速率、蒸腾速率、气孔导度及水分利用效率等参数,增强其抗旱能力,这无论对菌根化育苗,还是云南蓝果树生态恢复都将具有极其重要的实践意义,而且云南蓝果树幼苗生长指标和光合参数之间的相关性比较强,意味着AMF处理导致的净光合速率等光合能力的提高,使得云南蓝果树幼苗生长指标的增加。
3.3 不同水分条件下丛枝菌根真菌对云南蓝果树幼苗抗旱性的影响
可塑性指数在一定程度上可以反映植物的可塑性程度[17]。不论是水分充足还是干旱胁迫条件下,高AMF处理的云南蓝果树幼苗生长和光合参数的可塑性指数普遍较高,可以较好地适应干旱胁迫等逆境环境。
隶属函数分析提供了在多指标测定的基础上对材料进行综合评价的方法,更具科学性和可靠性[35]。本研究通过主成分分析简化了评价指标体系,确定了生长和光合特征的主要结构指标,并采用隶属函数值法进行了综合评价。结果显示,不管土壤水分条件如何(从W1到W6),AMF处理都显著影响了幼苗的抗旱性,即高AMF处理下幼苗的平均隶属函数值更高,具有更强的抗旱性。
研究证实水分显著影响丛枝菌根侵染率。Kaya等发现菌根侵染会显著提高黄瓜的产量及水分利用效率[36]。但是,从云南有气象记录的1961年以来,云南的年降水量则出现减少的趋势,半个世纪以来年降水量减少了39mm,减少速率为-8mm/10a,其中夏季和秋季减少趋势明显于春季和冬季。最显著的例子是云南蓝果树原产地西双版纳年降水日由20世纪50年代的每年270d锐减到目前的150d;年雾日由180d减少到30d,以往湿润的热带雨林气候已经发生明显变化。这有可能是导致云南蓝果树菌根侵染率降低的主要原因,而AMF侵染率的降低,影响了云南蓝果树幼苗的抗旱性。因此,云南蓝果树抗旱性不强的特点,加上逐渐干旱的原生境,种群逐渐变小,使得此物种的处境岌岌可危,如果不加以适当保护,本物种必将灭绝。
综合分析表明,干旱胁迫对云南蓝果树幼苗的生长和光合特征均产生了显著的影响,而云南蓝果树幼苗本身的抗旱性较差,但是AMF能与云南蓝果树根系形成良好的共生关系,一定程度上可以缓解干旱胁迫对植株所造成的伤害,增强其抗旱性,促进宿主植物生长。这说明自然生境下云南蓝果树幼苗数量少,天然更新缺乏很大一部分原因可以归结于当地生境的干旱,加上可以缓解其干旱胁迫的AMF也受当地气候导致水分缺乏的影响,也间接影响了云南蓝果树的抗旱性。目前对云南蓝果树开展了一系列保护措施,例如就地保护、近地保护、迁地保护和回归引种等。保护地的云南蓝果树近几年开始出现生长缓慢、部分死亡的现象。基于本研究云南蓝果树抗旱性不强的结果,在对云南蓝果树开展保护措施的过程中,建议将云南蓝果树种植在水源充足的生境,以便为云南蓝果树生长提供足够的水分;基于AMF可以增强云南蓝果树抗旱性的研究结果,建议对保护地的云南蓝果树施加AMF菌剂。
[1] 陈伟, 史富强, 杨文忠, 周云, 陈宏伟. 云南蓝果树的种群状况及生态习性. 东北林业大学学报, 2011, 39(9): 17- 19, 61- 61.
[2] 孙宝玲, 张长芹, 周凤林, 史富强, 吴之坤. 极度濒危植物——云南蓝果树的种子形态和不同处理条件对种子萌发的影响. 云南植物研究, 2007, 29(3): 351- 354.
[3] 孙宝玲, 张长芹. 极度濒危植物云南蓝果树的形态修订. 云南植物研究, 2007, 29(2): 173- 175.
[4] Poorter L, Hayashida-Oliver Y. Effects of seasonal drought on gap and understorey seedlings in a Bolivian moist forest. Journal of Tropical Ecology, 2000, 16(4): 481- 498.
[5] Engelbrecht B M J, Kursar T A. Comparative drought-resistance of seedlings of 28 species of co-occurring tropical woody plants. Oecologia, 2003, 136(3): 383- 393.
[6] McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytologist, 2008, 178(4): 719- 739.
[7] 刘文杰, 李红梅. 西双版纳旅游气候资源. 自然资源, 1997, (2): 62- 66.
[8] 宋富强, 赵俊斌, 张一平, 许再富, 肖来云. 西双版纳区域气候变化对植物生长趋势的影响. 云南植物研究, 2010, 32(6): 547- 553.
[9] Smith S E, Read D J. Mycorrhizal Symbiosis. 2nd ed. San Diego: Academic Press, 1997: 233- 289.
[10] Varma A. Mycorrhizae-the friendly fungi: What we know, what should we know, and how do we know? // Varma A ed. Mycorrhiza Manual. Berlin: Springer, 1998: 1- 24.
[11] Rai M, Acharya D, Singh A, Varma A. Positive growth responses of the medicinal plantsSpilanthescalvaandWithaniasomniferato inoculation byPiriformosporaindicain a field trial. Mycorrhiza, 2001, 11(3): 123- 128.
[12] Wang X R, Pan Q, Chen F X, Yan X L, Liao H. Effects of co-inoculation with arbuscular mycorrhizal fungi and rhizobia on soybean growth as related to root architecture and availability of N and P. Mycorrhiza, 2011, 21(3): 173- 181.
[13] Veresoglou S D, Shaw L J, Sen R.GlomusintraradicesandGigasporamargaritaarbuscular mycorrhizal associations differentially affect nitrogen and potassium nutrition ofPlantagolanceolatain a low fertility dune soil. Plant and Soil, 2011, 340(1/2): 481- 490.
[14] Klironomos J N, McCune J, Hart M, Neville J. The influence of arbuscular mycorrhizae on the relationship between plant diversity and productivity. Ecology Letters, 2000, 3(2): 137- 141.
[15] 何跃军, 钟章成, 刘锦春, 刘济明. 石灰岩土壤基质上构树幼苗接种丛枝菌根(AM)真菌的光合特征. 植物研究, 2008, 28(4), 452- 457.
[16] 陈冬青, 皇甫超河, 刘红梅, 王楠楠, 杨殿林. 水分胁迫和杀真菌剂对黄顶菊生长和抗旱性的影响. 生态学报, 2013, 33(7): 2113- 2120.
[17] 徐飞, 郭卫华, 徐伟红, 王仁卿. 刺槐幼苗形态、生物量分配和光合特性对水分胁迫的响应. 北京林业大学学报, 2010, 32(1): 24- 30.
[18] Ackerly D D, Bazzaz F A. Leaf dynamics, self-shading and carbon gain in seedlings of a tropical pioneer tree. Oecologia, 1995, 101(3): 289- 298.
[19] 王满莲, 冯玉龙. 紫茎泽兰和飞机草的形态、生物量分配和光合特性对氮营养的响应. 植物生态学报, 2005, 29(5): 697- 705.
[20] Abrams M D. Adaptations and responses to drought inQuercusspecies of North America. Tree Physiology, 1990, 7(1/4): 227- 238.
[21] 郑淑霞, 上官周平. 黄土高原油松和刺槐叶片光合生理适应性比较. 应用生态学报, 2007, 18(1): 16- 22.
[22] 宋会兴, 彭远英, 钟章成. 干旱生境中接种丛枝菌根真菌对三叶鬼针草(BidenspilosaL.)光合特征的影响. 生态学报, 2008, 28(8): 3744- 3751.
[23] 孙存华, 李扬, 杜伟, 金会丽, 王东升, 陈湘玲, 徐新娜. 干旱胁迫下藜的光合特性研究. 植物研究, 2007, 27(6): 715- 720.
[24] 陈昕, 徐宜凤, 张振英. 干旱胁迫下石灰花楸幼苗叶片的解剖结构和光合生理响应. 西北植物学报, 2012, 32(1): 111- 116.
[25] 付士磊, 周永斌, 何兴元, 陈炜. 干旱胁迫对杨树光合生理指标的影响. 应用生态学报, 2006, 17(11): 2016- 2019.
[26] Lawlor D W, Tezara W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes. Annals of Botany, 2009, 103(4): 561- 579.
[27] 田帅, 刘振坤, 唐明. 不同水分条件下丛枝菌根真菌对刺槐生长和光合特性的影响. 西北林学院学报, 2013, 28(4): 111- 115.
[28] 梁宇, 高玉葆, 陈世苹, 任安芝. 干旱胁迫下内生真菌感染对黑麦草实验种群光合、蒸腾和水分利用的影响. 植物生态学报, 2001, 25(5): 537- 543.
[29] 贺学礼, 赵丽莉. 非灭菌条件下VA菌根真菌对小麦生长发育的影响. 土壤通报, 1999, (2): 57- 59.
[30] 赵金莉, 贺学礼. AM真菌对油蒿生长和抗旱性的影响. 华北农学报, 2007, 22(5): 184- 188.
[31] 贺学礼, 李生秀. 不同VA菌根真菌对玉米生长及抗旱性的影响. 西北农业大学学报, 1999, 27(6): 49- 53.
[32] 贺学礼, 高露, 赵丽莉. 水分胁迫下丛枝菌根AM真菌对民勤绢蒿生长与抗旱性的影响. 生态学报, 2011, 31(4): 1029- 1037.
[33] Sánchez-Blanco M J, Ferrández T, Morales M A, Morte A, Alarcón J J. Variations in water status, gas exchange, and growth inRosmarinusofficinalisplants infected withGlomusdeserticolaunder drought conditions. Journal of Plant Physiology, 2004, 161(6): 675- 682.
[34] Morte A, Lovisolo C, Schubert A. Effect of drought stress on growth and water relations of the mycorrhizal associationHelianthemumalmeriense-Terfeziaclaveryi. Mycorrhiza, 2000, 10(3): 115- 119.
[35] 韩瑞宏, 卢欣石, 高桂娟, 杨秀娟. 紫花苜蓿抗旱性主成分及隶属函数分析. 草地学报, 2006, 14(2): 142- 146.
[36] Kaya C, Higgs D, KiInak H, Tas I. Mycorrhizal colonisation improves fruit yield and water use efficiency in watermelon (CitrulluslanatusThunb.) grown under well-watered and water-stressed conditions. Plant and Soil, 2003, 253(2): 287- 292.
Effects of arbuscular mycorrhizal fungi on growth and photosynthetic characteristics ofNyssayunnanensisseedlings under drought stress
ZHANG Shangshan, KANG Hongmei, YANG Wenzhong*, XIANG Zhenyong
KeyLaboratoryofRareandEndangeredForestPlantofStateForestryAdministration,YunnanAcademyofForestry,Kunming650201,China
Among various environmental stresses, drought stress has become a critical problem worldwide due to its dramatic effects on plant growth and physiology, and recently, this has attracted much attention. Arbuscular mycorrhizal fungi (AMF) have previously been reported to afford greater resistance to drought stress in some plant species. The aim of this study was to determine the impacts of AMF on growth and photosynthetic characteristics ofNyssayunnanensisseedlings through addition (Low AMF) or non-addition (High AMF) of the fungicide benomyl at each water treatment. We further aimed to elucidate drought resistance and associated microbiological mechanisms of this endangered plant species, which would help to establish and implement protection measures. In this study, a pot experiment was conducted to control the soil water content, and 100, 91.68, 82.85, 60.00, 41.86, and 21.28% of field moisture capacity were set. The seedlings were subjected to different watering regimes and measurements were made at the end of the experiment to investigate the growth and photosynthetic characteristics ofN.yunnanensisseedlings. Plasticity index analysis and principal component analysis of 18 indices ofN.yunnanensiswere integrated and analyzed using the membership function method to evaluate synthetic drought resistance. These variables provide a theoretical basis for a better understanding of the mechanism of AMF on the tolerance ofN.yunnanensisseedlings to drought stress. The results showed that the AMF colonization rate ofN.yunnanensisroots significantly decreased under “Low AMF” treatments, indicating that differences in the growth and photosynthetic characteristics of AMF-treated seedlings were caused by a reduction in the rate of AMF colonization inN.yunnanensisroots. With the intensification of drought stress, the AMF colonization rate ofN.yunnanensisroots significantly decreased and changed both growth parameters, including leaf area, and photosynthetic characteristics, including net photosynthetic rate ofN.yunnanensisseedlings. High AMF significantly enhancedN.yunnanensistolerance to the imposed drought stress by improving both growth parameters and photosynthetic characteristics under conditions of adequate moisture and mild drought stress. Nevertheless, there were no significant differences between AMF treatments under severe drought stress, indicating that this had more significant effects onN.yunnanensisseedlings than did AMF. The results also demonstrated that theN.yunnanensisseedlings could not adapt to diverse water stress through plastic responses in morphology and photosynthesis, whereas high AMF treatments led to higher plastic differences in both growth parameters and photosynthetic characteristic.N.yunnanensisseedlings showed stronger drought resistance under high AMF conditions. The experimental results provide a theoretical basis for the scientific conservation and breeding ofN.yunnanensis. In conclusion,N.yunnanensisseedlings did not show resistance to drought stress, but plants were dependent on mycorrhizae under water stress, which partially compensated its effects.
arbuscular mycorrhizal fungi (AMF); drought stress; growth parameters; photosynthetic characteristics;Nyssayunnanensis
国家自然科学基金项目(31460119);国家林业局珍稀濒危物种野外救护与繁育项目(2014YB1004, 2015YB1021);云南省应用基础研究青年项目(2013FD075)
2015- 04- 27;
日期:2016- 03- 03
10.5846/stxb201504270862
*通讯作者Corresponding author.E-mail: wzyang2004@126.com
张珊珊,康洪梅,杨文忠,向振勇.干旱胁迫下AMF对云南蓝果树幼苗生长和光合特征的影响.生态学报,2016,36(21):6850- 6862.
Zhang S S, Kang H M, Yang W Z, Xiang Z Y.Effects of arbuscular mycorrhizal fungi on growth and photosynthetic characteristics ofNyssayunnanensisseedlings under drought stress.Acta Ecologica Sinica,2016,36(21):6850- 6862.
