坡向因子对黄土高原草地群落功能多样性的影响
2016-12-28朱云云王孝安邓美皎
朱云云,王孝安,王 贤,邓美皎
陕西师范大学生命科学学院, 西安 710119
坡向因子对黄土高原草地群落功能多样性的影响
朱云云,王孝安*,王 贤,邓美皎
陕西师范大学生命科学学院, 西安 710119
研究群落水平上的植物功能性状特征及功能多样性随坡向的变化规律,对认识不同坡向上的植物群落形成过程具有重要意义。以黄土高原不同坡向上的自然草地群落为研究对象,比较研究了植物功能性状(株高和比叶面积)和功能多样性测度指标(功能丰富度、功能均匀度和功能离散度)随坡向的变化规律。研究结果显示:(1)阴坡的株高和比叶面积显著高于其他坡向;(2)一元性状的功能丰富度在不同坡向间均无显著差异;阴坡和半阴坡的多元性状功能丰富度显著高于阳坡;(3)阴坡的株高功能均匀度显著高于半阳坡,而比叶面积功能均匀度在各坡向的差异并不显著;多元性状功能均匀度在不同坡向差异显著,阴坡最高,半阳坡最低;(4)阴坡和半阴坡的株高功能离散度显著高于阳坡和半阳坡,而半阴坡的比叶面积功能离散度显著高于阳坡;半阴坡的多元性状功能离散度Rao指数显著高于阳坡。研究结果暗示了,在阴坡和半阴坡上,植物对群落内的生态位空间和资源利用更充分,种间竞争强度较低,不同物种之间生态位高度分化;而在阳坡和半阳坡上,由于水分等条件的限制植物可占据的生态位空间有限,导致其对占据的生态位空间使用不足,物种间资源竞争较强烈。在群落水平上,研究地区的植物功能性状及功能多样性随坡向的规律性变化,反映了黄土高原植被群落构建过程中坡向因子对功能性状的筛选效应。该研究结果对该区的植被恢复重建的物种选择及植被布局规划具有重要实践意义。
坡向;群落构建;功能性状;功能丰富度;功能均匀度;功能离散度
环境筛选和物种间的相互作用决定着自然群落的物种多样性和功能多样性[1- 4]。环境筛选的作用使得群落中的物种具有相似的生态特征以适应环境,而物种间的相互作用则增大了群落中物种的不相似性以实现物种的稳定共存[2]。这两种机制对群落构建具有不同的作用,一方面维持群落的结构和生物多样性,另一方面通过影响生态位的分化对群落内物种性状的分布产生影响,从而影响群落的功能多样性[3- 4]。功能多样性的测度与分析可用来判断群落对当前和将来环境筛的反应[5]。
李新娥研究发现山区不同的坡向对于植物群落构建是一个强有力的环境筛选[6]。在阴坡-阳坡地形,具有复杂多变的环境因子,例如光照、土壤湿度、土壤温度和土壤矿物质含量[7- 9]。在这个微型气候梯度上,群落结构的渐变也随之产生[7,10-11],在植物群落组成、物种多样性和生产力等方面产生了很大差异,这对造成不同坡向植物不同生活型的生物量分配差异有重要意义[11]。目前,国内外关于阴坡-阳坡植物群落的研究主要是描述性的,从功能性状以及功能多样性的角度去揭示群落构建以及环境对其影响作用的研究则较少[6]。
许多学者已提出计算功能多样性的多种指标[12-18],其中Mason等[16]由物种多样性的组成类推,将功能多样性分解成3个基本组分:功能丰富度、功能均匀度和功能离散度。每个组分均表示功能多样性的不同方面:功能丰富度测量群落中现有物种占据了多少生态位空间,功能均匀度测量物种性状在所占据性状空间的分布规律,功能离散度测量群落功能性状的多度分布在性状空间中的最大离散程度[16]。由此可见,这种分解不仅反映了性状在空间分布的互补性,也有利于剔除一些冗余指数。但国内目前尚无从这3方面探讨群落构建的研究。本文以黄土高原马栏地区不同坡向的自然草地群落为研究对象,通过分析草本植物的功能性状特征及功能丰富度、功能均匀度和功能离散度对坡向的响应,探讨坡向对草地群落功能性状和功能多样性指数的影响,阐释黄土高原草地群落对潜在的环境变化或者环境筛选的反应,回答坡向是如何对黄土高原草地群落的构建起作用的,以期预测未来草地植物群落发展模式的变化。研究结果可预测小尺度的植被动态变化,为群落构建机制的理论研究和黄土高原植被恢复重建实践提供科学依据。
1 研究方法
1.1 研究区域
该研究选在陕西省旬邑县东北部马栏林区的4个不同坡向的山坡草地进行。该林区地处黄土高原中部,地理位置为108°27′—108°52′E,35°9′—35°33′N,海拔约1200—1700m。该地区气候特征属于暖温带半湿润地区,年均气温7.4—8.5℃,12月至来年2月最低气温可达-27.7℃,7—9月最高气温可达36.7℃。年均降水量580 mm,多集中在7—9月,湿热同期,易于植物生长。阳坡草地的主要植物有细叶苔草(Carexrigescens)、蒿(Artemisiaspp.)等,种类较少;而阴坡的主要植物有百里香(Thymusmongolicus)、早熟禾(Poaannua)、萎陵菜(Potentillachinensis)、风轮草(Clinopodiumchinensis)、达乌里胡枝子(Lespedezadavurica)等,种类较多。
1.2 研究方法
1.2.1 试验设计和野外调查
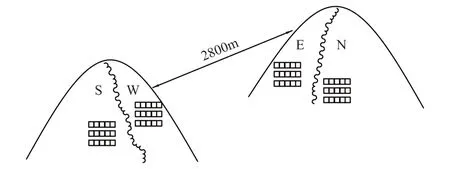
图1 样地空间分布示意图 Fig.1 Schematic Diagram of Spatial Distribution of Sampling Sets S:阳坡 Sunny Slope;W:半阳坡 Semi-sunny Slope;E:半阴坡 Semi-cloudy Slope;N:阴坡 Cloudy Slope
在对马栏地区的植被分布进行全面考察的基础上,选择不同坡向的4个样地(图1),分别在阳坡、半阳坡、半阴坡和阴坡各设置1个草本植物群落样地,在每个样地中随机选择3条1 m × 5m的样带,将每个样带分成5个1 m × 1m的样方,即每个样地设置15个1 m × 1m的样方,调查每个样方中的植物种类、数目和盖度。同时每条样带选3个点,用Hydra土壤水分仪(Microscan Electronics Pty.Ltd, USA)测温度、湿度和盐分等土壤指标[19];用pH分度计(Spectrum Technologies lnc, USA)测定土壤pH值[20];并测定土壤厚度;记录每条样带的坡度、坡向、坡位、海拔高度等环境指标。其中在坡向定值时,参考王孝安[21]的转换方法,南坡为180计,正北坡以0计,正东、正西坡计作90;坡向每偏南1度时,增加值为2;每偏北1度时,则减少2,数值越大表示越向阳、越干热。
1.2.2 样本采集和性状的选择测量
Westoby[22]提出了叶片-高度-种子植物生态策略方案,强调了叶片和高度的重要性。成熟株高表达了植物在两个干扰之间的生长数量,反映植物群落的生存方式和种群动态,与光吸收、热负荷、风速、湿度、对支持叶片的投资成本和叶片对水分的吸收相关[22]。比叶面积为分配给每单位光合干物质的捕光面积,被广泛认为是与植物的相对生长速率和资源利用相关的重要性状,反映了对快速生长机会的响应变化[22]。根据草地群落的结构特点、动态特性[1,23]、功能性状[12]和生态系统功能特性[24],选择了能很好反映植物竞争强度、响应环境变化的两个功能性状指标——株高和比叶面积。
在4个坡向12个样带的77个物种中,采集每个物种的10株成熟、完整、发育正常、无病虫害的个体的地上部分(不足10株的植物种类不予采集),为减少水分蒸发及叶形的变化,整株装入塑封袋,外罩黑色塑料袋及时带回驻扎地[25];测量其株高,在每个植株的中部各选取2片成熟、展开、无病虫害的叶子,保证每个物种至少20片叶子,进行叶性状的测量[6]。用CI- 202叶面积仪(CID, Inc., USA)扫描叶面积;然后装入纸袋中,在60℃温度下烘48 h,用电子天平(Sartorius, Beijing)称其叶片干重,计算公式为SLA = LA/LDW,式中SLA为比叶面积,LA为叶面积,LDW为叶干重[24-25]。
1.2.3 功能多样性计算公式
(1)功能丰富度
一元性状功能丰富度应用Mason等[16]提出的functional range指数计算,它计算的是一个群落中一个性状生态位空间占全部群落中该性状生态位空间的比例,这个指数范围是0—1;多元性状的功能丰富度是每个一元性状功能丰富度的平均值[16]。具体公式如下:
式中,FRci为群落i中性状c的功能丰富度,SFci为群落中物种所占据的生态位空间,Rc为所有群落中性状c占据生态位空间。
(2)功能均匀度
一元性状的功能均匀度应用Mouillot(2005)提出的functional regularity指数计算[14],它通过单一的性状值(如均值、中值等)和物种多度计算,公式如下:
式中,S为物种数,PEWi,i+1为物种i和i+1性状值权重差异的百分比,Ai为物种i的相对多度,Ci为物种i的性状值。
当性状空间是多元的时候,采用Villeger等[17]提出的FEve指数计算,公式如下:
式中,S为物种数,EW为均匀度权重,dist(i,j)为物种i和j的欧氏距离,wi为物种i的相对丰富度,I为分支长,PEWI为分支长权重。
(3)功能离散度
一元性状功能离散度的计算参考Lepš等修正的FDvar指数[12,16,23],具体公式如下:
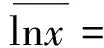
多元性状功能离散度采用Rao指数[13,15,26]计算,该指数测量种群内和种群间的多样性和差异性,计算公式如下:
式中,FDQ为Rao指数,pi和pj为物种i和j的相对多度,S为物种数,dij为物种i和j的差异,n为性状数,Xik为物种i的k性状值,Xjk为物种j的k性状值。
1.2.4 数据分析
通过Excel软件和FDiversity软件协助计算4个坡向每条样带的物种丰富度、性状平均值和每个性状的功能丰富度指数(FRci)、功能均匀度指数(FRO和FEve)和功能离散度指数(FDvar和FDQ),其中功能均匀度指数和功能离散度指数的计算在软件FDiversity中完成[27-28],功能丰富度指数在Excel中输入公式完成计算。
运用单因素方差分析以及LSD后置检验法检验坡向对物种丰富度、性状值、功能丰富度指数、功能均匀度指数和功能离散度指数影响的差异,设置95%置信区间。用SPSS 20.0进行单因素方差分析的显著性检验及逐步回归分析,用GraphPad prism5作图。
2 结果与分析
2.1 4个坡向的环境因素及物种丰富度
4个坡向的大部分环境因素差异显著(表1),阳坡和半阳坡的坡度显著比半阴坡和阴坡大,半阴坡的显著高于阴坡。阳坡的pH显著低于阴坡,半阳坡的pH显著低于半阴坡和阴坡。阳坡的土厚显著低于半阴坡和阴坡。半阳坡的土壤温度显著高于阳坡、半阴坡和阴坡。阳坡和半阳坡的土壤湿度显著低于半阴坡和阴坡。阳坡的土壤含盐量显著低于阴坡,半阳坡的土壤含盐量显著低于半阴坡和阴坡。
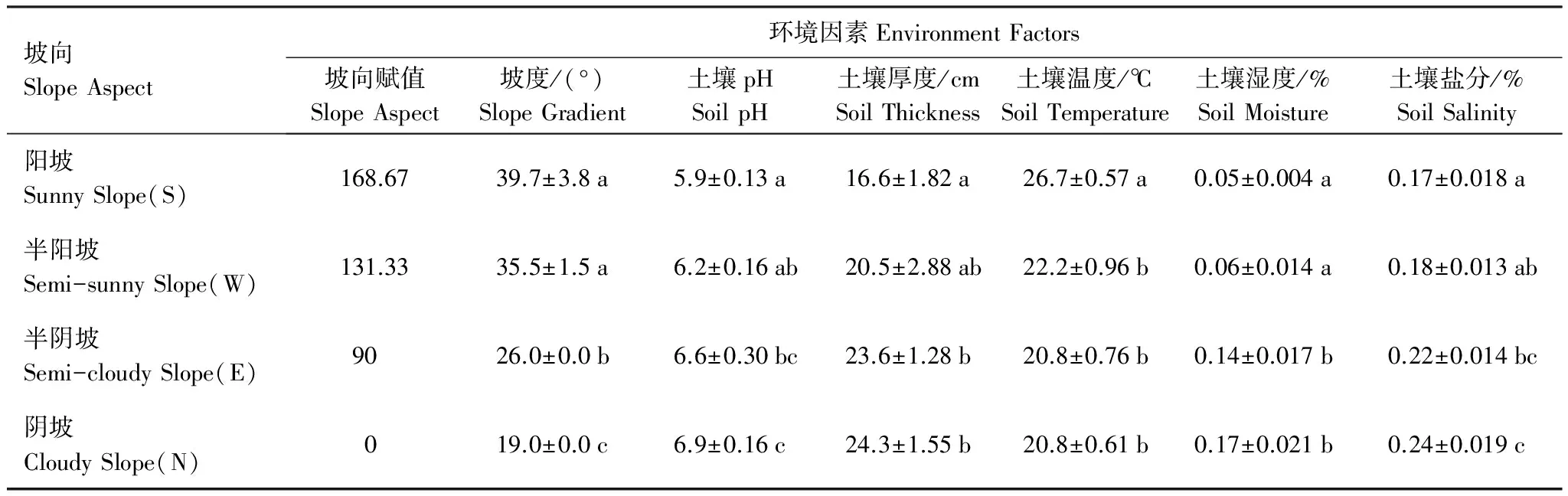
表1 不同坡向的环境因素
P<0.05;a、b、c字母相同,无显著差异;字母不同,差异显著
4个坡向植物群落的物种丰富度差异较大(图2),由于阳坡和半阳坡环境因素相似程度高,所以物种丰富度无显著差异;而阳坡和半阳坡的物种丰富度皆显著低于半阴坡和阴坡;半阴坡和阴坡之间物种丰富度也有显著差异,半阴坡的物种丰富度值在4个坡向中最高。而通过逐步回归建模发现影响物种丰富度最主要的因素是土壤湿度和坡向(表2),且土壤湿度起主导作用。
2.2 坡向与物种性状
株高和比叶面积的性状均对坡向敏感(图3)。株高表示植物对光的竞争能力,在4个坡向上的大小顺序为阴坡>半阳坡>阳坡>半阴坡,其中阴坡的株高平均值显著高于阳坡和半阴坡,半阳坡的株高平均值显著高于半阴坡。比叶面积反映植物捕获光照资源的能力,在4个坡向上的大小顺序为阴坡>半阳坡>半阴坡>阳坡,其中阴坡显著高于阳坡。通过逐步回归分析,发现没有特定的环境因素对物种性状有显著影响,不同坡向的物种性状差异是所有环境因素综合影响的结果。

表2 环境与各指数的线性回归模型
SM: 土壤湿度,SA: 坡向,ST: 土壤厚度,SG: 坡度,STE: 土壤温度,SS: 土壤盐分
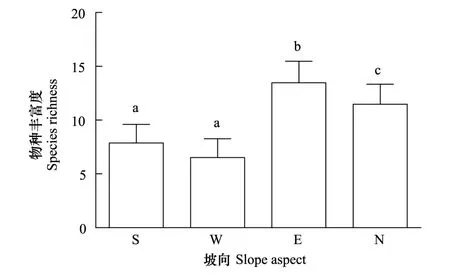
图2 阳坡、半阳坡、半阴坡和阴坡的物种丰富度 Fig.2 The Species Richness of Sunny Slope(S), Semi-sunny Slope(W), Semi-cloudy Slope(E) and Cloudy Slope(N)
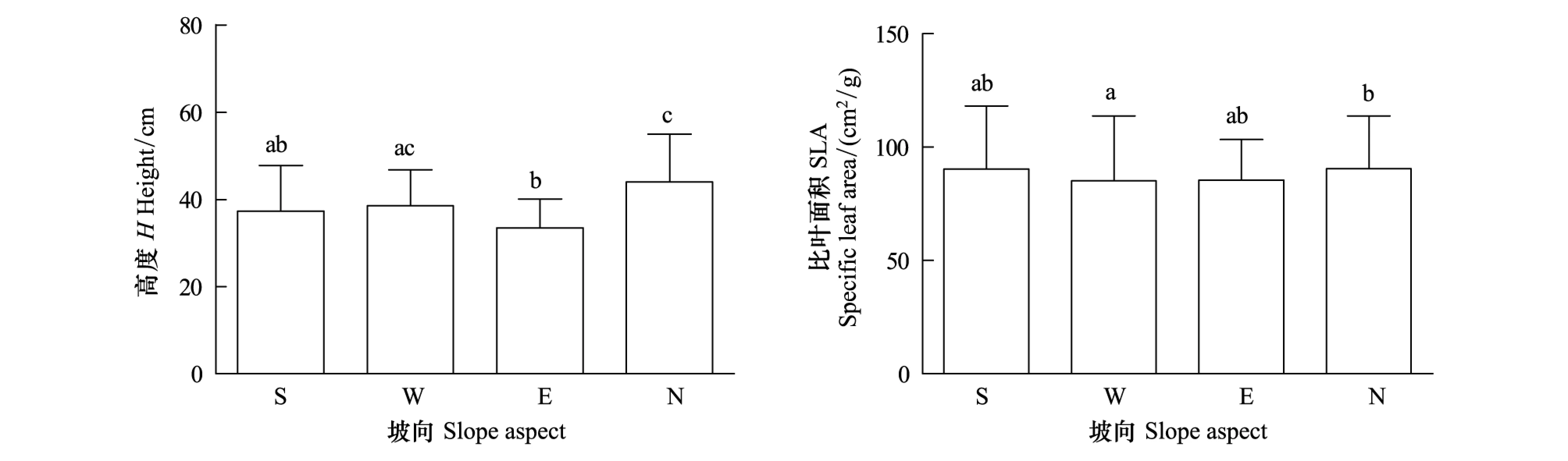
图3 阳坡、半阳坡、半阴坡和阴坡的植物功能性状Fig.3 The Plant Trait Mean of Sunny Slope(S), Semi-sunny Slope(W), Semi-cloudy Slope(E) and Cloudy Slope(N)
2.3 功能多样性
株高和比叶面积的一元功能丰富度在4个坡向并无明显差异,但均在阳坡最低(图4)。株高的功能丰富度值在各坡向大小顺序为半阴坡>半阳坡>阴坡>阳坡。比叶面积功能丰富度在各坡向的大小顺序为半阳坡>半阴坡>阴坡>阳坡。而株高和比叶面积两个性状的多元功能丰富度在4个坡向上有显著差异,其大小顺序为阴坡>半阴坡>半阳坡>阳坡,其中阴坡和半阴坡的功能丰富度显著大于阳坡。回归分析发现高度的功能丰富度与环境因素无显著回归关系,是各环境因素综合影响的结果;比叶面积的功能丰富度受土壤厚度影响,且二者为正相关关系;株高和比叶面积的多元功能丰富度受pH影响,二者也为正相关关系(表2)。
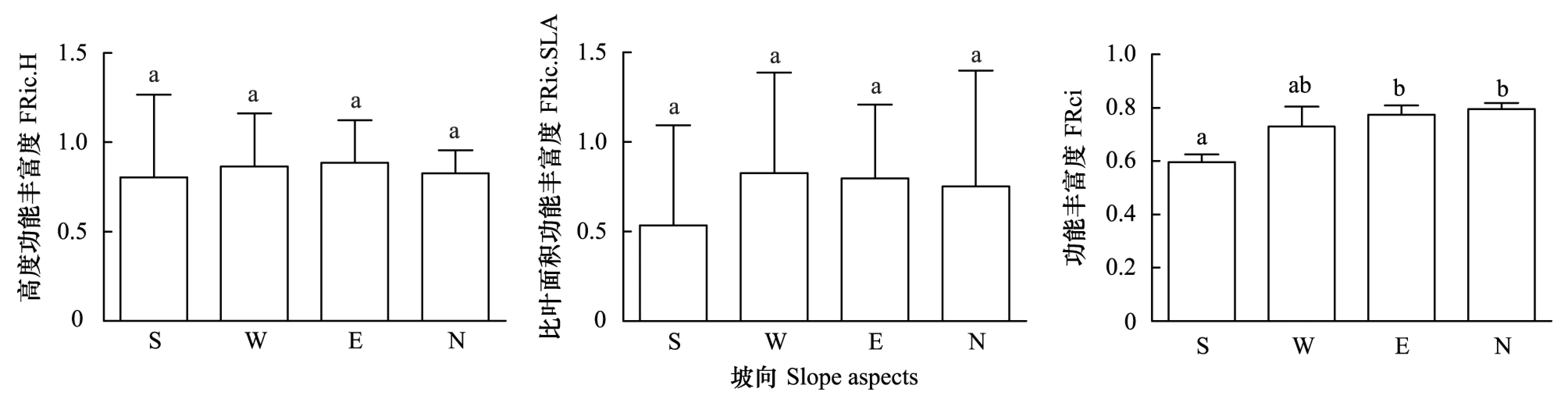
图4 阳坡、半阳坡、半阴坡和阴坡的植物功能丰富度Fig.4 Plant Functional Richness of Sunny Slope(S), Semi-sunny Slope(W), Semi-cloudy Slope(E) and Cloudy Slope(N)
不同坡向株高的功能均匀度均有显著差异(图5)。株高功能均匀度在4个坡向上的大小顺序为阴坡>阳坡>半阴坡>半阳坡,其中阴坡的株高功能均匀度值显著高于半阳坡。比叶面积功能均匀度在4个坡向上的大小顺序为阴坡>半阴坡>半阳坡=阳坡,但是差异并不显著。两个性状的多元功能均匀度指数在不同坡向差异显著(图5)。其大小顺序为阴坡>半阴坡>阳坡>半阳坡,其中阴坡的多元功能均匀度显著高于阳坡和半阳坡,半阴坡显著高于半阳坡。通过回归分析发现,并无哪个环境因子显著影响单个性状的功能均匀度,但土壤盐分显著影响多元功能均匀度,且为促进作用(表2)。
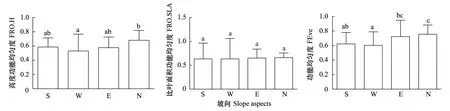
图5 阳坡、半阳坡、半阴坡和阴坡的植物功能均匀度Fig.5 Plant Functional Evenness of Sunny Slope(S), Semi-sunny Slope(W), Semi-cloudy Slope(E) and Cloudy Slope(N)
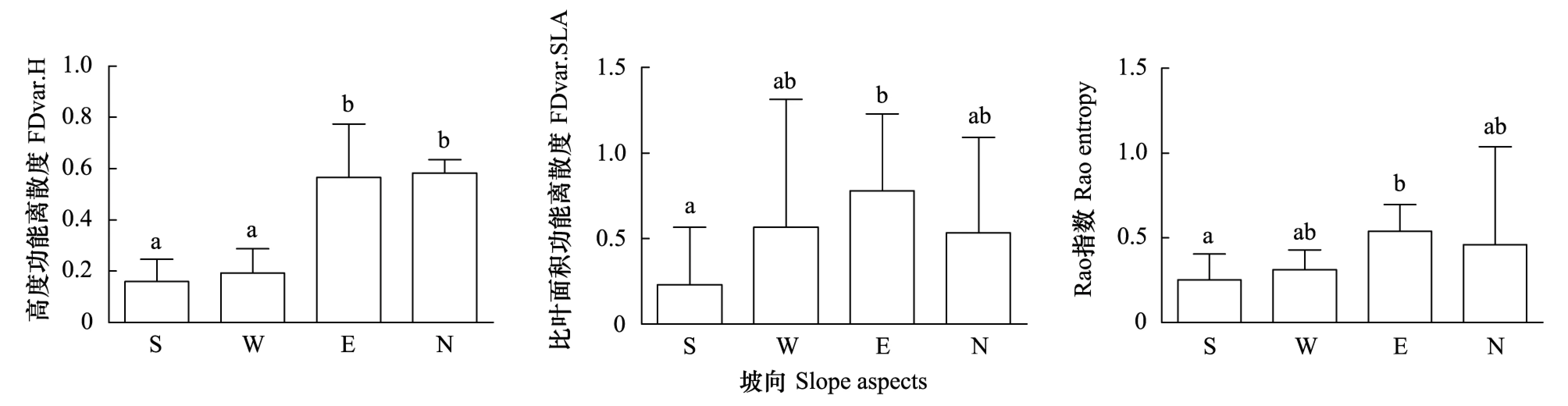
图6 阳坡、半阳坡、半阴坡和阴坡的植物功能离散度Fig.6 Plant Functional Divergence of Sunny Slope(S), Semi-sunny Slope(W), Semi-cloudy Slope(E) and Cloudy Slope(N)
不同坡向上株高和比叶面积的功能离散度均有显著差异(图6)。株高的离散度沿坡向变化顺序为阴坡>半阴坡>半阳坡>阳坡,其中阴坡和半阴坡的株高功能离散度值皆显著高于阳坡和半阳坡;比叶面积的功能离散度沿坡向的大小顺序为半阴坡>半阳坡>阴坡>阳坡,其中半阴坡的值显著高于阳坡;以Rao指数计算的多元性状功能离散度在不同坡向上的大小顺序为半阴坡>阴坡>半阳坡>阳坡,其中半阴坡与阳坡差异显著。功能离散度和环境的回归模型表明影响不同性状功能离散度的环境因子各不相同(表2):坡度影响物种高度的功能离散度,二者呈负相关;土壤温度影响物种比叶面积的功能离散度,二者为负相关关系;土壤盐分影响以Rao计算的两个性状综合离散度,且二者为正相关关系。
3 讨论
3.1 不同坡向的环境因素与物种丰富度
在山地环境条件下,植物的生存受大地形因子(海拔)和小地形因子(坡向和坡位)的共同影响[10,29]。从阴坡到阳坡,环境因子发生了明显变化。一般来讲,阳坡的坡度比阴坡陡峭,本研究中的样地也是如此(阳坡31°,阴坡21°),这容易发生营养流失,而且在雨季土壤流失更为严重,造成阳坡土壤矿化严重,土壤养分含量低于阴坡[30]。此外,坡向的差异直接影响了土壤环境条件,包括温度、湿度[31-32],土壤发育和生产[33],土壤团聚体的稳定性[34]以及土壤体积密度、孔隙度、营养利用和渗透系数[35],而且坡向和植被也影响了土壤中的有机碳和氮含量[36]以及微生物性质[37]。本研究测量结果显示(表1),阴坡和半阴坡土壤盐分含量较阳坡和半阳坡有显著提高,这与其他研究结果相似,即阴坡具有相对较高的土壤养分含量[10,38]。本研究中阴坡的土壤明显比阳坡厚,这进一步证明阳坡较陡的坡度易造成土壤流失和土壤矿化。已有多个研究证明,在不同坡向,土壤含水量是限制群落构成最主要的因素,且阴坡土壤含水量显著高于阳坡[11,39],本研究结果与此相符(表2),土壤湿度和坡向影响了各坡向的物种丰富度的差异,导致阴坡和半阴坡的物种丰富度显著高于阳坡和半阳坡,且阴坡和半阴之间的差异也有显著性(图2)。
3.2 植物功能性状对不同坡向的响应
阴坡和阳坡的各种环境因素综合影响山地植物群落植物物种[10]和植物功能性状的分布[30,40]。植物的株高是一个用来表现植物内部资源分配模式和总体竞争能力(特别是对光的竞争)的性状[41-42]。当在不同群落和生态环境比较物种时,株高也与其他特征,比如生物量、根深、横向传播和叶大小相关[24]。叶性状在碳同化,水分关系和能量平衡中扮演十分重要的作用,例如叶面积与“光获取策略”和“为光的获取分配的质量数”有关[12, 22],叶面积对于叶能量收支和全株水分平衡有重要影响[24]。而比叶面积与潜在相对生长率正相关[22],在幼年期间,具有高比叶面积的物种新叶生长快速,转换率高的植物部分对光和土壤资源也有一个更灵活的响应空间[22]。本文研究结果显示阴坡的株高和比叶面积都较其他坡向高。这是由于阴坡土层更深,土壤发育更好,并且土壤微生物对枯枝落叶有更大的消化潜力[37],从而产生更高的营养利用空间,更适合植物生长。在实地观察中可以看到阴坡的总植被覆盖率较大,植物株高较高,叶片较大,这会加剧资源竞争,从而形成较高的初级成产力[22],这一点也可应用于以高产为目标的草甸管理和恢复中[43]。通常情况下,在资源有限的生境中,生长缓慢的物种比叶面积均较低,而在土壤营养资源丰富的地方,比叶面积均较高[24],这正如本研究的结果:土壤营养资源丰富的阴坡的比叶面积显著高于其他坡向。
3.3 植物功能多样性对不同坡向的响应
通过使用这3个基于单个性状的功能多样性指数[17],可以评估本研究所选的环境筛(坡向)如何形成与群落构建联系的植物功能性状。功能丰富度测量群落中现有物种占据多少生态位空间[16],低功能丰富度表示没有完全利用潜在可用的资源,这将减小生产力[44]。如果功能特征代表物种对环境的耐受性,低的功能丰富度则意味着某种环境条件下,物种无法生存,导致该生态位空间利用不足,减弱对环境波动的缓冲能力[45]。本研究结果显示株高和比叶面积的一元功能丰富度对坡向的变化不敏感,但多元功能丰富度对坡向变化敏感,且阳坡显著低于其他各坡,可见植物多元性状功能丰富度对坡向这个环境筛具有明显的响应。这说明阳坡植物的生长发育受到其生存环境的限制,并无充分利用生态空间。回归分析发现并无单个环境因子限制株高的功能丰富度,但比叶面积功能丰富度受土壤厚度影响,也就是土层越厚的土壤条件下植物的比叶面积占据的生态位空间越大。而株高和比叶面积多元功能丰富度受土壤pH影响,即土壤越偏酸性,FRci指数越低,性状在多维生态位空间的分布越“狭窄”, 这间接地也可能与土壤盐分含量和种类有关。该地区土壤整体呈弱酸性(表1),由此可见土壤偏酸程度越高,越不利于植物生长。
功能均匀度指数反映物种性状在所占据的生态位空间的分布规律,可用来指示资源的使用不足或使用过度[16]。高的功能均匀度指数意味着性状在生态位空间的分布是非常有规律的;而低的功能均匀度指数说明被占据的某部分生态位空间没有被充分利用,存在物种分布间隙,这会导致生产力和稳定性减小,入侵机会增加[16,18]。本研究结果显示,坡向对株高的功能均匀度均响应明显,对比叶面积功能均匀度响应不明显,对株高和比叶面积综合功能均匀度响应明显。与半阳坡相比,阴坡植物的株高在生态位空间的分布更规律,资源利用更充分;阴坡和半阴坡的多元功能均匀度显著比半阳坡在生态位空间的分布更规律,资源利用也更充分。可见较之阴坡和半阴坡,阳坡或者半阳坡存在物种分布间隙。由回归分析发现,土壤盐分影响了多元性状功能均匀度。也就是说,土壤养分越丰富,物种分布间隙越小,性状在多维生态位空间的分布更规律,资源利用也更充分。
功能离散度指数反映功能群或者功能性状在群落性状空间的离散程度[16]。性状平均值的高分歧表明群落中生态位分化程度较高,由此产生较低的资源竞争,因此具有高功能离散度的群落或许会增加生态系统功能,更有效的资源利用;低功能离散度则暗示着这些区域生态位分化程度较低,资源利用不充分,资源竞争较为剧烈[16]。本研究结果显示阳坡和半阳坡的株高的离散度显著低于阴坡和半阴坡,这说明阴坡的生态位分化程度高于阳坡且资源利用较充分,资源竞争强度较弱,而阳坡植物的生存则要面临更强的资源竞争。比叶面积的离散度为半阴坡最高,阳坡最低,这说明比叶面积在半阴坡离散程度最高,生态位空间利用充分,叶性状具有较高的适应能力。回归分析发现影响不同性状功能离散度的因素均不同,坡度影响株高在生态位空间的离散程度,土壤温度影响比叶面积在生态位空间的离散程度,而土壤盐分影响多元功能离散度,说明不同性状的功能离散度对不同的环境因子响应不同。
通过比较这3种多样性测度指标,发现山地植物的功能性状多样性对当地环境筛的强度变化具有明显的响应,环境筛过程是影响该地区的植物群落构建过程的一个重要因素。除环境筛外,物种间相互作用也影响植物群落构建的理论,这两个理论对群落构建的相对作用是相反的,而且在不同的研究尺度,起主导作用的因子不同,在坡向这个大尺度上,环境筛的作用更为显著[46]。但是通过比较影响不同指标的环境因子,发现坡向对性状的筛选作用不是由某个单一确定的因子决定的,不同的环境因素影响性状在生态位空间分布的不同方面。本研究中土壤湿度对功能多样性的影响并不显著,这与土壤湿度是影响群落中物种分布的主要限制性因子[11]的说法不同,由此可见影响物种分布和性状分布的环境因子并不一定相同。了解环境筛对群落形成的影响也可预测一个群落物种性状的大致分布,判断具有某种性状的物种是否适应生存在此环境中,为当地植被重建提供理论依据。否则一旦人工种植了一个不适应当地环境的物种就可能会造成经济和生态的双重损失,所以遵循环境与植被的适应机制,了解环境筛对群落构建的作用,对该区的植被恢复具有重要的指示作用[47]。
4 小结
群落生态学最重要的问题之一是生态环境如何影响自然群落中复杂的物种组成。基于群落构建理论,环境筛是影响群落物种分布的一个重要方面。尤其是当环境筛越过生态影响阈值的时候,植物性状和群落构(重)建过程对于环境筛的变化相当敏感。本文通过比较黄土高原马栏地区山地的各坡向植物功能性状和3种多样性测度指标的变化,发现阴坡和半阴坡的植物性状占据了更多的生态位空间,资源利用较充分且竞争较低,并且出现了高度的生态位分化;而阳坡和半阳坡由于水分等条件的限制植物性状可占据的生态位空间有限,且在占据的生态位空间资源使用不足,导致生态位分化程度较低,资源竞争比较强烈。本研究结果为环境筛与群落构建之间的联系提供了实例研究证据,揭示了坡向对草本植物群落形成的影响原因,为黄土高原草地植被恢复及重建提供了理论依据。
[1] Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741- 758.
[2] Grime J P. Trait convergence and trait divergence in herbaceous plant communities: Mechanisms and consequences. Journal of Vegetation Science, 2006, 17(2): 255- 260.
[3] De Bello F, Lepš J, Sebastià M T. Variations in species and functional plant diversity along climatic and grazing gradients. Ecography, 2006, 29(6): 801- 810.
[4] De Bello F, Thuiller W, Lepš J, Choler P, Clément J C, Macek P, Sebastià M T, Lavorel S. Partitioning of functional diversity reveals the scale and extent of trait convergence and divergence. Journal of Vegetation Science, 2009, 20(3): 475- 486.
[5] Díaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646- 655.
[6] 李新娥. 亚高寒草甸阳坡—阴坡梯度上植物功能性状及群落构建机制研究[D]. 兰州:兰州大学, 2011.
[7] Auslander M, Nevo E, Inbar M. The effects of slope orientation on plant growth, developmental instability and susceptibility to herbivores. Journal of Arid Environments, 2003, 55(3): 405- 416.
[8] Tomimatsu H, Hori Y. Effect of soil moisture on leaf ecophysiology ofParasenecioyatabe, a summer-green herb in a cool-temperate forest understory in Japan. Journal of Plant Research, 2008, 121(1): 43- 53.
[9] Nevo E. Evolution in action across phylogeny caused by microclimatic stresses at “Evolution Canyon”. Theoretical Population Biology, 1997, 52(3): 231- 243.
[10] Gong X, Brueck H, Giese K M, Zhang L, Sattelmacher B, Lin S. Slope aspect has effects on productivity and species composition of hilly grassland in the Xilin River Basin, Inner Mongolia, China. Journal of Arid Environments, 2008, 72(4): 483- 493.
[11] Sternberg M, Shoshany M. Influence of slope aspect on Mediterranean woody formations: Comparison of a semiarid and an arid site in Israel. Ecological Research, 2001, 16(2): 335- 345.
[12] Mason N W H, MacGillivray K, Steel J B, Wilson J B. An index of functional diversity. Journal of Vegetation Science, 2003, 14 (4): 571- 578.
[13] Ricotta C. A note on functional diversity measures. Basic and Applied Ecology, 2005, 6 (5): 479- 486.
[14] Mouillot D, Mason W H N, Dumay O, Wilson J B. Functional regularity: a neglected aspect of functional diversity. Oecologia, 2005, 142(3): 353- 359.
[15] Botta-Dukát Z. Rao′s quadratic entropy as a measure of functional diversity based on multiple traits. Journal of Vegetation Science, 2005, 16 (5): 533- 540.
[16] Mason N W H, Mouillot D, Lee W G., Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112- 118.
[17] Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290- 2301.
[18] Schleuter D, Daufresne M, Massol F, Argillier C. A user′s guide to functional diversity indices. Ecological Monographs, 2010, 80 (3): 469- 484.
[19] 刘庆生, 李集生, 刘高焕, 宋创业. Hydra土壤测试仪测定现代黄河三角洲的土壤水分与盐分试验. 现代农业科学,2009, (1):161- 162.
[20] 国家质量监督检验检疫总局. 实验室pH(酸度)计检定规程(JJG119- 2005). 北京: 中国计量出版社, 2005.
[21] 王孝安. 甘南玛曲植物群落的多元分析与环境解释. 生态学报, 1997, 17(1): 61- 65.
[22] Westoby M. A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil, 1998,199(2): 213- 227.
[23] Lepš J, de Bello F, Lavorel S, Berman S. Quantifying and interpreting functional diversity of natural communities: practical considerations matter. Preslia, 2006, 78: 481- 501.
[24] Cornelissen J H C, Lavorel S, Gamier E,. Díaz S, Buchmann N, Gurvich D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for Standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335- 380.
[25] Pérez-Harguindeguy N, Díaz S, Garnier E, Lavorel S, Poorter H, Jaureguiberry P, Bret-Harte M S, Cornwell W K, Craine J M, Gurvich D E, Urcelay C, Veneklaas E J, Reich P B, Poorter L, Wright I J, Ray P, Enrico L, Pausas J G, de Vos A C, Buchmann N, Funes G, Quétier F, Hodgson J G, Thompson K, Morgan H D, ter Steege H, van der Heijden M G A, Sack L, Blonder B, Poschlod P, Vaieretti M V, Conti G, Staver A C, Aquino S, Cornelissen J H C. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61 (3): 167- 234.
[26] Rao C R. Diversity and dissimilarity coefficients: a unified approach. Theoretical Population Biology,1982, 21 (1): 24- 43.
[27] Casanoves F, Pla L, Di Rienzo J A, Díaz S. FDiversity: a software package for the integrated analysis of functional diversity. Methods in Ecology and Evolution, 2011, 2(3): 233- 237.
[28] Casanoves F, Pla L E, Di Rienzo J A. FDiversity: an Integrated Tool to Estimate and Analyze Functional Diversity. Bulletin of the Ecological Society of America, 2011, 92(2): 147- 152.
[29] 祁建, 马克明, 张育新. 北京东灵山不同坡位辽东栎(Quercusliaotungensis)叶属性的比较. 生态学报, 2008, 28(1): 122- 128.
[30] 杜峰, 梁宗锁, 徐学选, 张兴昌, 山仑. 陕北黄土丘陵区撂荒群落土壤养分与地上生物量空间异质性. 生态学报, 2008, 28(1):13- 22.
[31] Bennie J, Huntley B, Wiltshire A, Hill M O, Baxter R. Slope aspect and climate: spatially explicit and implicit models of topographic microclimate in chalk grassland. Ecological Modelling, 2008, 216 (1): 47- 59.
[32] Sidari M, Ronzello G, Vecchio G, Muscolo A. Influence of slope aspects on soil chemical and biochemical properties in aPinuslaricioforest ecosystem of Aspromonte (Southern Italy). European Journal of Soil Biology, 2008, 44(4): 364- 372.
[33] Egli M, Mirabella A, Sartori G, Zanelli R, Bischof S. Effect of north and south exposure on weathering rates and clay mineral formation in Alpine soils. Catena, 2006, 67(3): 155- 174.
[34] Schoorl J M, Boix Fayos C, de Meijer R J, van der Graaf E R, Veldkamp A. The137Cs technique applied to steep Mediterranean slopes (Part I): the effects of lithology, slope morphology and land use. Catena, 2004, 57 (1): 15- 34.
[35] Wang L, Wang Q J, Wei S P, Shao M A, Li Y. Soil desiccation for Loess soils on natural and regrown areas. Forest Ecology and Management, 2008, 255(7): 2467- 2477.
[36] Yimer F, Ledin S, Abdelkadir A. Soil organic carbon and total nitrogen stocks as affected by topographic aspect and vegetation in the Bale Mountains Ethiopia Geoderma, 2006, 135: 335- 344.
[37] Huang Y M, Dong L, An S S. Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. Catena, 2015,125: 135- 145.
[38] 潘占兵, 余峰, 王占军, 李生宝, 张源润. 宁南黄土丘陵区坡向、坡位对苜蓿地土壤含水量时空变异的影响. 水土保持研究, 2010, 17(2): 141- 144.
[39] Martínez J A, Armesto J J. Ecophysiological plasticity and habitat distribution in three evergreen sclerophyllous shrubs of the Chilean matorral. Oecologica Plantarum, 1983, 4(3): 211- 219.
[40] 刘旻霞, 马建祖. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应. 应用生态学报, 2012, 23(12): 3295- 3300.
[41] Weiher E, van der Werf A, Thompson K, Roderick M, Garnier E, Eriksson O. Challenging Theophrastus: a common core list of plant traits for functional ecology. Journal of Vegetation Science, 1999, 10 (5): 609- 620.
[42] Westoby M, Falster D S, Moles A T, Vesk P A, Wright I J. Plant ecological strategies: some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 2002, 33: 125- 159.
[43] 吕亭亭, 王平, 燕红, 廖桂项, 姜海波, 邹畅林, 盛连喜. 草甸和沼泽植物群落功能多样性与生产力的关系. 植物生态学报, 2014, 38(5): 405- 416.
[44] Petchey O L. Integrating methods that investigate how complementarity influences ecosystem functioning. Oikos, 2003, 101(2): 323- 330.
[45] Tilman D. Biodiversity: population versus ecosystem stability. Ecology, 1996, 77(2): 350- 363.
[46] Mouchet M A, Villéger S, Mason N W H, Mouillot D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 2010, 24 (4): 867- 876.
[47] 龚时慧, 温仲明, 施宇. 延河流域植物群落功能性状对环境梯度的响应. 生态学报, 2011, 31(20): 6088- 6097.
Effect of slope aspect on the functional diversity of grass communities in the Loess Plateau
ZHU Yunyun, WANG Xiao′an*, WANG Xian, DENG Meijiao
ShaanxiNormalUniversity,CollegeofLifeSciences,Xi′an710119,China
Functional diversity of a plant community, which is based on plant traits, has been proposed as a key component predicting ecosystem function. Knowledge about how plant functional traits and functional diversity change along with the different slope aspects at the community level is important for understanding the formation of plant communities under distinct slope aspects. However, to date, previous studies on plant communities in sunny and cloudy slope environments have mainly been descriptive. To our knowledge, no research has dealt with the relationship between the slope aspect and community assembly based on plant functional diversity (i.e., functional richness, evenness and divergence). In this study, we aimed to uncover the relationship between slope aspects and plant functional traits. Three sampling transects (1 m × 5 m) were established on each of four sites facing north, south, east, or west, in Malan Mountain, Loess Plateau. Each of the sampling transects were evenly divided into fifteen 1 m × 1 m plots, and a plot survey was conducted. We measured plant height (H) and specific leaf area (SLA), and then calculated three types of functional diversity indices (i.e., functional richness, evenness, and divergence), which included three one-dimensional indices based on single traits (i.e., FRci, FRO, FDvar) and three multi-dimensional indices based on two traits (i.e., FRci, FEve, FDQ). A comparison of the changing pattern of plant functional traits and the three types of functional diversity was conducted among the four slope aspects (the cloudy, sunny, semi-cloudy, and semi-sunny slope). Our results indicated that: (1) the value of H and SLA on the cloudy slope was greater than the others; (2) there was no significant difference among the four slope aspects in one-dimensional functional richness indices of H and SLA, while the multi- dimensional functional richness indices of plants on the cloudy slope and the semi-cloudy slope were greater than those of the plants on the sunny slope; (3) on the cloudy slope, the one-dimensional functional evenness of H was significantly greater compared to those on the semi-sunny slope, while that of SLA was not significantly influenced by the slope aspect; the multi-dimensional functional evenness indices of plants on the cloudy slope was greater than those of the plants on semi-sunny slope; (4) the one-dimensional functional divergence of H on the cloudy and semi-cloudy slope were significantly greater than those on the semi-sunny and sunny slope, while that of SLA on the semi-cloudy slope was significantly greater than those on the sunny slope; the multi-dimensional functional divergence index and Rao′s quadratic entropy of plants was significantly greater on the semi-cloudy slope compared with those on the sunny slope. Our results indicated that for plant traits, more niche space was occupied, more resources were sufficiently exploited, lower competition intensity occured, and a high level of niche differentiation existed on the cloudy and semi-cloudy slopes. By contrast, because of the limitation of all kinds of soil conditions and environmental factors, plant traits occupied less niche space on the sunny and semi-sunny slope, resources were exploited less sufficiently, and competition for resources tended to be stronger. At the community-level, the changing pattern of plant functional traits and functional diversity along the four different slope aspects suggests the existence of an environmental fitter effect on functional traits under the process of community assembly in the Loess region. These findings would have significant practical implications for species selection and vegetation rehabilitation layout planning in the hilly area of the Loess Plateau.
slope aspect; community assembly; functional trait; functional richness; function evenness; functional divergence
陕西师范大学中央高校基本科研业务费(GK201503044)
2015- 05- 01;
2015- 09- 21
10.5846/stxb201505010900
*通讯作者Corresponding author.E-mail: wangxa@snnu.edu.cn
朱云云,王孝安,王贤,邓美皎.坡向因子对黄土高原草地群落功能多样性的影响.生态学报,2016,36(21):6823- 6833.
Zhu Y Y, Wang X A, Wang X, Deng M J.Effect of slope aspect on the functional diversity of grass communities in the Loess Plateau.Acta Ecologica Sinica,2016,36(21):6823- 6833.
