森林鼠类对秦岭南坡3种壳斗科植物种子扩散的影响
2016-12-28石子俊陈晓宁李金钢
张 博,石子俊,陈晓宁,侯 祥,王 京,李金钢,常 罡,*
1 陕西师范大学生命科学学院,西安 710062 2 陕西省动物研究所,西安 710032
森林鼠类对秦岭南坡3种壳斗科植物种子扩散的影响
张 博1,石子俊1,陈晓宁2,侯 祥2,王 京2,李金钢1,常 罡2,*
1 陕西师范大学生命科学学院,西安 710062 2 陕西省动物研究所,西安 710032
种子作为森林鼠类的主要食物来源,它们的扩散和更新很大程度上依赖于鼠类的传播。在鼠类扩散种子的过程中,种子特征和食物相对丰富度是影响鼠类对种子进行何种选择策略的重要因素。2011—2012年的8—12月,采用塑料片标记法在秦岭南坡的佛坪国家级自然保护区内调查了森林鼠类对同域分布的3种壳斗科植物(锐齿槲栎Quercusalienavar.acuteserrata、栓皮栎Q.variabilis和短柄枹栎Q.serratavar.brevipetiolata)种子的扩散差异。结果表明:(1)鼠类倾向于贮藏营养价值较大的栓皮栎种子,并且其贮藏距离也最远(2011:1.52 m,2012:4.03 m),3种种子在食物相对丰富度较低年份被贮藏的距离均较远。(2)在食物相对丰富度较高的年份(2011),种子的消耗速率较慢,在种子释放10 d后种子释放点仍有67.33%的种子,贮藏量较高,至实验结束仍有29.67%的种子被贮藏。在食物相对丰富度较低的年份(2012),种子消耗速率较快,在种子释放后10 d内所有种子均被取食或搬离种子释放点,贮藏量较低,至实验结束仅有3.83%的种子仍被贮藏。(3)虽然栓皮栎种子的贮藏量最大,被贮藏后的存留量也最大,但其在实验地的分布却较小,说明种子扩散仅是植物分布与存活的第一步。以上结果表明,鼠类倾向于贮藏营养价值高的种子。在食物相对丰富度较高的年份会更多的贮藏种子,但种子被贮藏的距离较近,在食物相对丰富度较低的年份会更多的取食种子。
种子特征;贮藏行为;食物相对丰富度;种子扩散;种子选择;种子消耗
许多森林鼠类都具有贮藏植物种子的行为,贮藏行为对于鼠类度过冬眠期、哺乳期、食物匮乏期等特殊时期具有决定性作用[1]。鼠类的贮藏行为可以分为集中贮藏和分散贮藏,其中分散贮藏行为对于植物的更新和分布具有重要作用。当少数分散贮藏的种子最终逃脱捕食后,就有可能萌发并建成幼苗,实现植物的自然更新[2]。壳斗科植物是亚热带和温带地区森林群落中的优势种或建群种,在生态系统中占据着重要的地位。森林鼠类往往是这些壳斗科植物种子的重要捕食者和主要扩散者[3]。种子的物理和化学特征是影响鼠类对其取食和扩散的重要因素,已有研究表明种子大小(种子重量、长度、直径等)[4- 5]、种皮厚度[6- 7]、单宁含量[8- 9]、萌发时间[10- 11]、营养含量[10- 11]和昆虫蛀蚀[12- 13]等种子特征对鼠类的取食和贮藏策略具有重要影响。此外非种子自身特征的因素,如食物丰富度、鼠类密度等也是影响鼠类扩散种子的重要因素[14]。
锐齿槲栎、栓皮栎和短柄枹栎均为秦岭地区广泛分布的壳斗科植物,在秦岭地区的生态系统中占据重要的地位[15- 16]。在这些植物种子的成熟季节,鼠类会对其进行捕食和贮藏。尽管已有一些研究关注秦岭地区鼠类对植物种子的扩散,但是这些研究更多地集中在单一的种子,很少有研究关注同域分布的多种种子由于种子特征等因素而导致的扩散差异[17]。因此,本研究拟通过野外释放3种同域分布的壳斗科种子,探讨鼠类对种子的取食和扩散差异。同时,分析种子特征和其他因素(食物丰富度和鼠类密度)对种子扩散的影响,以期为秦岭地区壳斗科植物的自然更新和保育提供一些理论资料。
1 材料与方法
1.1 研究地点
本项研究在陕西省佛坪县秦岭南坡的佛坪国家级自然保护区内进行。该保护区位于107° 41′—107° 41′E,33°33′—33°46′N。年均气温11.4 ℃,年均降水943 mm, 海拔处于980—2904 m 之间[18]。保护区内植被表现出明显的垂直分布格局,自下向上依次为落叶阔叶林、针叶林、斑块状分布的亚高山灌丛和草甸。本研究样地位于海拔1200 m左右的落叶阔叶林带[19]。该地区分布的啮齿动物种类达28种之多,分别隶属于7科18属,其中本研究样地属于中低山落叶阔叶林区的北社鼠(Niviventerconfucianus)+中华姬鼠(Apodemusdraco)群落[20]。
1.2 研究物种及标记方法
本研究的3种植物均隶属于壳斗科(Fagaceae)栎属(Quercus),落叶乔木。其中锐齿槲栎花期3—4月,果期9—10月。在我国分布较广,秦岭南北坡均产,较为习见,生于海拔700—2000 m的山坡上,常自生成纯林,或与短柄枹栎、栓皮栎等组成落叶阔叶混交林,在南坡也常与马尾松组成松栎林[15- 16]。短柄枹栎花期4—5月,果实次年9—10月成熟。在我国分布较广,秦岭南坡较为习见,阳性树种,萌发力极强,生于海拔1300 m以下的山沟和山坡[15- 16]。栓皮栎初夏开花,单性同株,花期5月,果期9—10月。在我国分布很广,秦岭地区分布也较为广泛,常自生为纯林,分布于海拔500—1800 m的山沟和山坡上[15- 16]。
当这3种植物种子成熟时(9—10月),在野外收集新鲜种子用于当年实验。本实验采用塑料片标记法对实验种子进行标记。具体标记方法详见常罡[21]。该标记法目前在种子扩散实验中已被广泛使用,且对鼠类的影响较小[21]。
1.3 鼠类群落结构调查
2011和2012年8—10月分别在实验地区对鼠类群落结构进行了调查,具体的操作过程是在落叶阔叶林带内选择5个调查点,每个调查点随机选取3条样线,每10 m放置一个活捕笼,每条样线置20笼,样线间相隔15 m以上。以生花生做诱饵,连续捕捉3 d,对所有捕获的鼠类进行常规测量和记录后原地释放。
1.4 种子雨调查
3种壳斗科树种各选取8棵样树,每个树下设置一个种子收集框,框子的平面面积约为1 m2。每年8月20日开始每7 d调查1次,对收集到的种子进行鉴别和记录,持续到11月下旬没有种子掉落后结束[22]。
1.5 种子特征及营养测定
2011年9月底,随机选取每种树各8棵样树(共24棵),在每棵样树的树冠下捡取种子160粒。分别测定种子的重量、长度、宽度及种皮厚度。同时将完好的种子去种皮并剪碎,混匀后烘干,将烘干的样品送至农业部谷物品质监督检验测试中心测定种子营养成分。
1.6 种子扩散实验设计
2011年10月初,在实验样地内的落叶阔叶林带随机选取10个样点作为种子释放点,任意两个种子释放点相距20 m以上。单个种子释放点面积约为1 m2,每个样点均放置3种被标记的种子各20粒(共60粒),在释放后的第1,3,5,10,17,27,50天进行调查[12],调查面积约为以种子释放点半径为30 m的范围。2012年在相同样地重复上述实验。
调查时记录所有种子的命运及扩散距离。种子命运分为以下6种:(1)原地存留,种子在释放点完好保留,未被取食或移动;(2)原地取食,种子在释放点被鼠类取食;(3)扩散取食,种子被鼠类搬运后取食;(4)贮藏,种子被鼠类埋藏于地表;(5)拖动,种子被鼠类搬运后弃置在地表,未被取食或贮藏;(6)丢失(拖动和丢失的种子未做数据分析)[23]。在被埋藏和拖动的种子旁用红色木棍标记位置,并在其上用黑色记号笔添加种子编号。结合皮尺和步量获得扩散距离,用指南针测量并记录方向。
1.7 统计与分析
所有数据均使用OriginPro 9.0作图,其中种子命运动态图分别以贮藏累积量和单日的原地存留量、原地被取食量、扩散后被取食量为纵坐标,以天数为横坐标、标准误为误差线做折线图。使用SPSS 22.0进行数据分析。采用一般线性模型单因素方差分析(命运动态为因变量,种子类别为固定因子,天数为协变量)来比较3种种子在原地存留、被原地取食、被扩散取食和被贮藏时间动态上的差异;采用单因素方差分析比较3种种子在贮藏距离和物理特征的差异,使用Bonferroni法进行多重比较。定义P值小于0.05时为存在显著性差异,小于0.01时为存在极显著差异。
2 实验结果
2.1 鼠类种类及捕获率
两年间共捕获鼠类132只,2012年鼠类的捕获率远高于2011年(2011:3.38%,2012:16.15%)。在捕获的鼠类中,北社鼠和中华姬鼠数量较多,占到总数的92.54%,说明这两类鼠类是该地区的优势种。实验期间捕获的其他鼠类为岩松鼠(Sciurotamiasdavidianus)(2.99%)、甘肃仓鼠(Cansumyscanus)(2.99%)、苛岚绒鼠(Eothenomysinez)(1.49%)。另外,虽然岩松鼠捕获率较低,但是野外观测到的数量却较多,这主要是由于岩松鼠体型较大且比较机敏,故难以捕获。通过饲养笼喂食观察可以确定该地区5种鼠类均取食本研究中的3种壳斗科植物种子。
2.2 种子密度
通过种子雨调查发现,两年间种子降落到地表的密度(粒 d-1m-2)以锐齿槲栎(2011年: 0.90; 2012年: 0.43)为最高,其次为短柄枹栎(2011年: 0.64; 2012年: 0.30),栓皮栎(2011年: 0.16; 2012年: 0.19)种子密度最低。2011年锐齿槲栎和短柄枹栎的种子密度相对更高,达到2012年的2倍多,栓皮栎种子两年间差别很小。
2.3 种子物理和化学特征
3种植物的种子以栓皮栎体积和重量最大,短柄枹栎体积和重量最小,栓皮栎的种壳厚度厚度最大,短柄枹栎的种壳厚度最小(表1)。3种种子的重量(F2,3339=3114.49,P<0.01)、长度(F2,3339=1351.20,P<0.01)、直径(F2,3339=2839.93,P<0.01)、壳厚(F2,3339=2583.80,P< 0.01)均存在极显著差异,选用Bonferroni法进行多重比较,发现以上物理特征在3种种子两两间均为极显著差异。
栓皮栎的脂肪、淀粉和纤维含量最高,3种种子的单宁含量接近。因此,综合比较而言,栓皮栎种子营养价值最大,短柄枹栎营养价值最小(表1)。
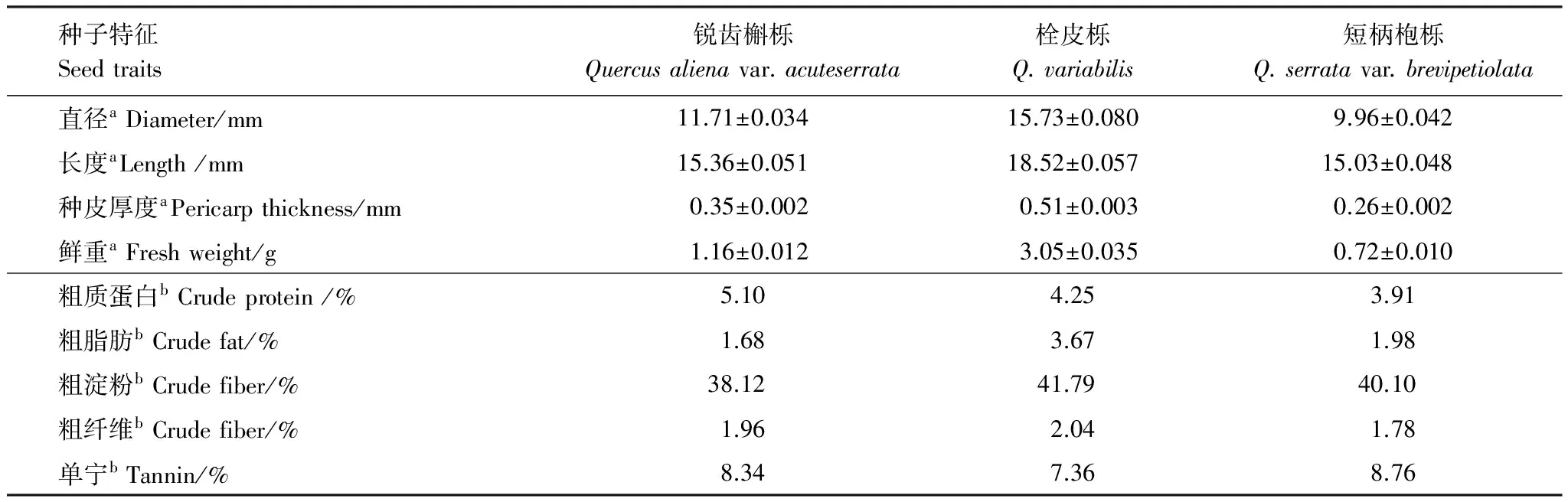
表1 种子理化特征
a 平均值±标准误(Mean ± S.E.) 样本量N=160 b 种子化学成分由农业部谷物品质监督检验测试中心测定(NO. K120041-K120047)
2.4 种子命运
2011年3种植物种子的消失速率都较慢,但是三者之间差异极显著(F2,600=32.29,P<0.01),截至实验结束后锐齿槲栎和短柄枹栎仍有少量种子存留在原地。2012年3种植物种子存留时间较短,在释放后第10天均被取食或扩散,三者之间没有表现出显著差异(F2,600=1.00,P=0.370)(图1)。
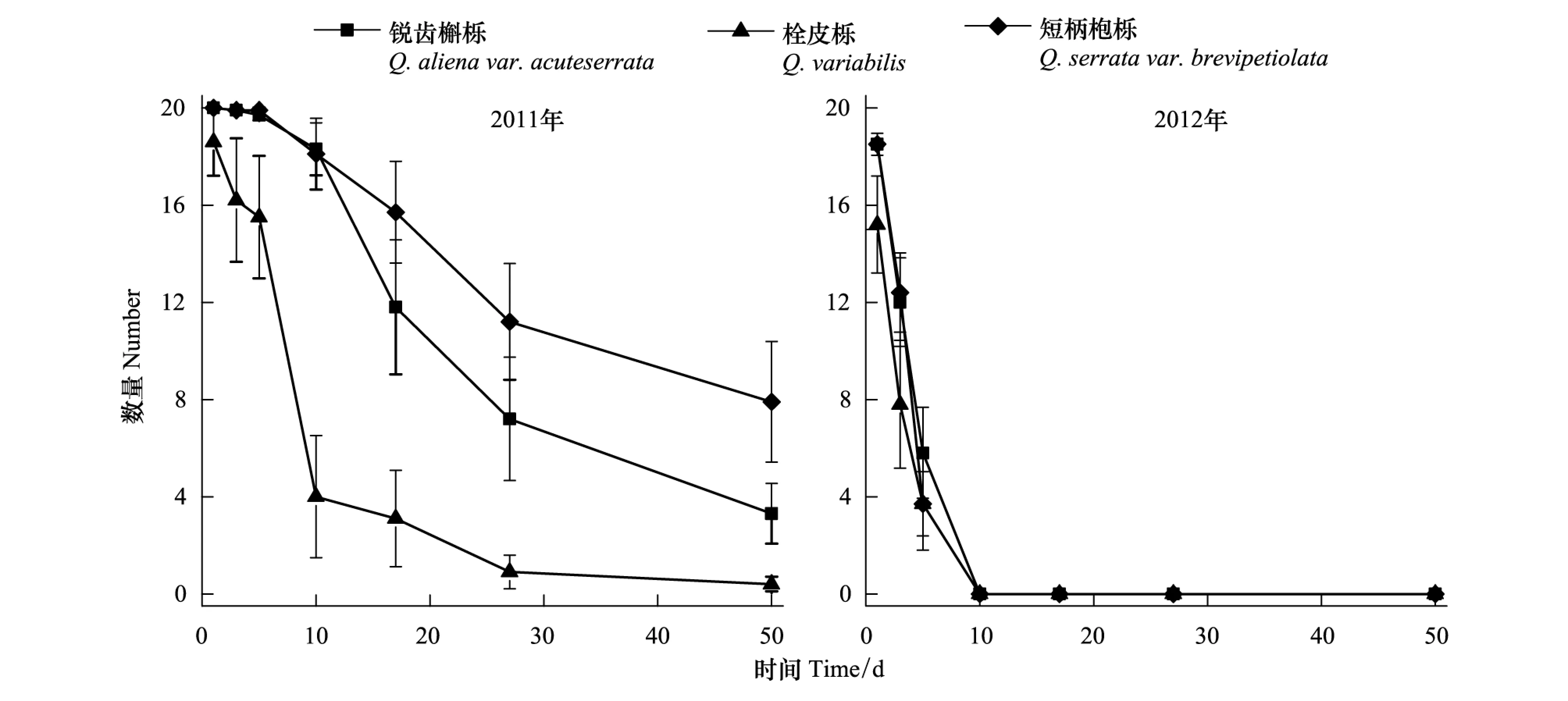
图1 种子原地存留的动态Fig.1 Dynamic of seeds that were remained in situ
3种种子的原地被取食动态在两年内均存在极显著差异(2011:F2,600=5.133,P<0.01 ; 2012:F2,600=6.038,P<0.01)。栓皮栎种子的原地被取食率均低于其余2种植物,短柄枹栎的原地被取食时间均略早于其它2种种子(图2)。
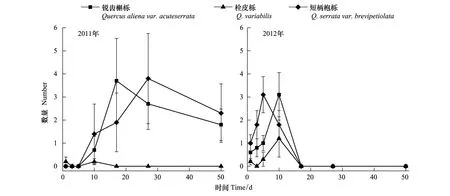
图2 种子被原地取食的动态Fig.2 Dynamic of seeds that were eaten in situ
2年中3种种子的扩散后被取食动态差异均不显著(2011:F2,600=1.662,P=0.192; 2012:F2,600=0.636,P=0.530) (图3)。
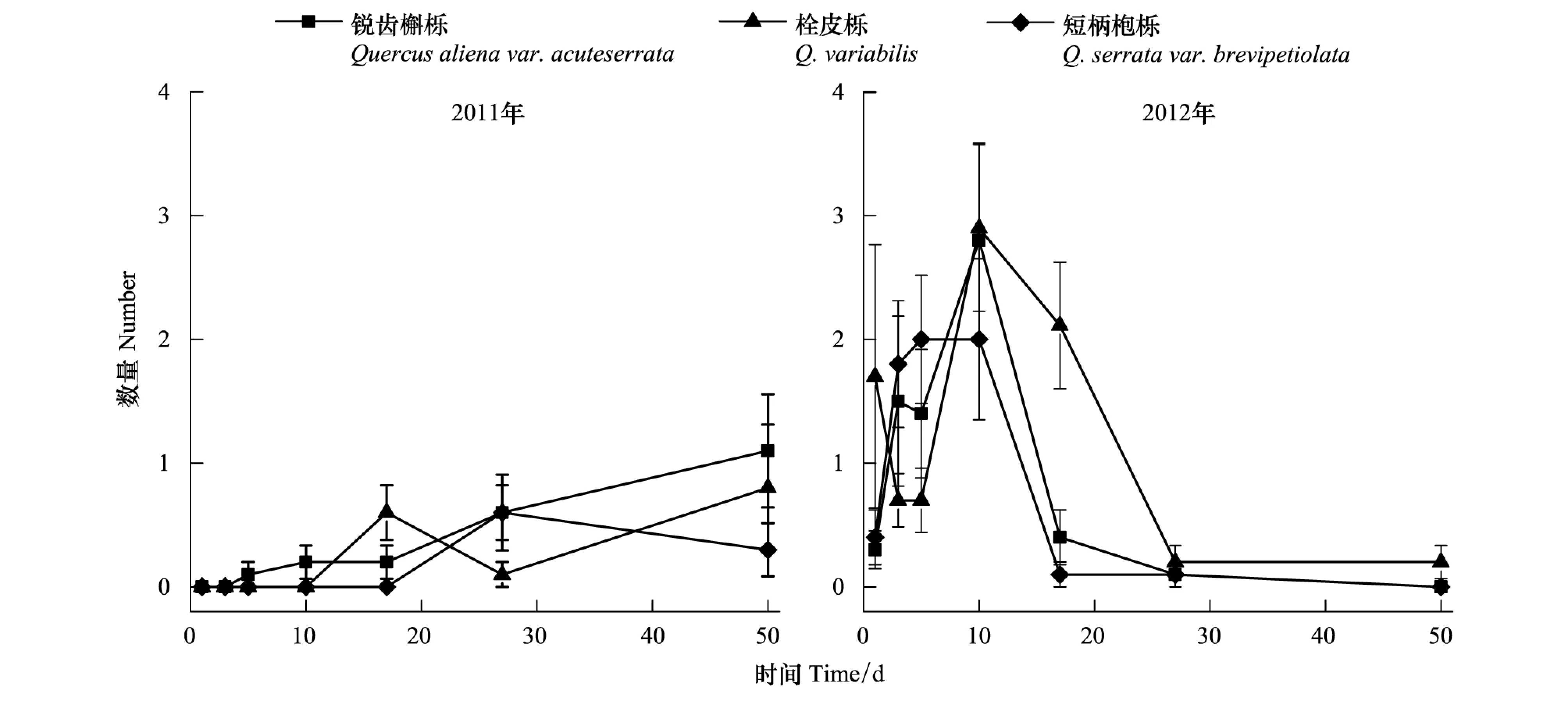
图3 种子被扩散取食的动态Fig.3 Dynamic of seeds that were eaten after removal
2011和2012年3种种子的贮藏动态均存在显著的差异(2011:F2,600=73.50,P<0.01; 2012:F2,600=19.57,P< 0.01),其中栓皮栎的贮藏量均高于其余2种种子。这些被贮藏的种子在实验期间存活了较长时期 (图4)。
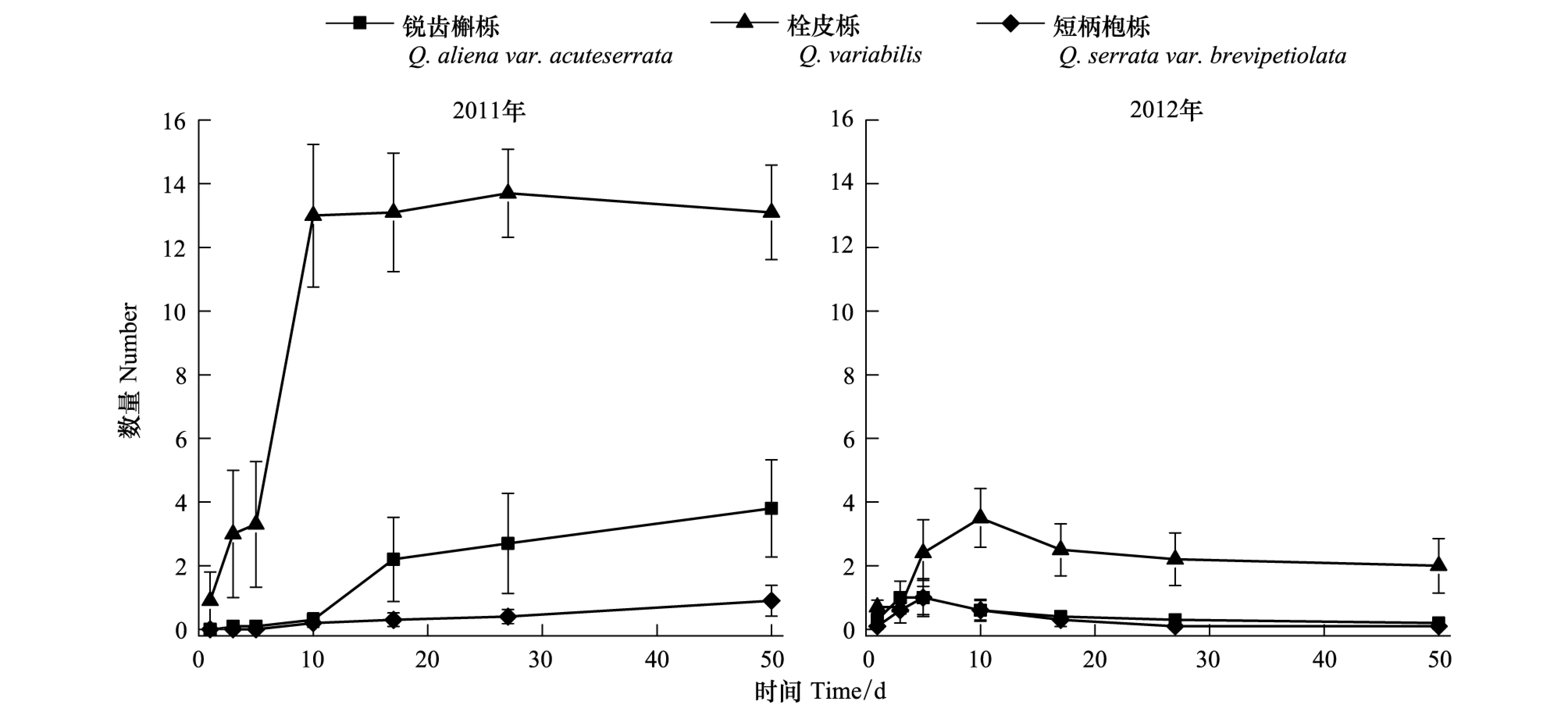
图4 种子被贮藏的动态Fig.4 Dynamic of seeds that were cached
2.5 贮藏距离
2011年3种种子的贮藏距离不存在显著差异(F2,199=0.832,P=0.437),而2012年存在极显著差异(F2,69=10.275,P< 0.01)。两年中栓皮栎种子被贮藏的距离均最远,短柄枹栎被贮藏的距离最近。2011年种子贮藏量大于2012年,但同时其被贮藏的距离则小于2012年(表2)。

表2 种子被贮藏的距离
3 讨论
不同的植物种子具有不同的营养价值,对于鼠类而言,对这些不同价值的种子作出何种选择是非常重要的,直接决定了其与同类以及其他物种竞争时存活的可能性。一般认为鼠类会将营养价值较高的种子扩散到较远的距离后再进行取食或贮藏,而营养价值较低的种子,鼠类则更倾向于原地取食[4- 6]。本研究中,栓皮栎种子质量最重、体积最大、脂肪含量最高,其对于鼠类而言价值应该是最大的。实验结果发现鼠类对栓皮栎的原地取食量最低而扩散后的取食量较大,同时栓皮栎种子的扩散距离较远且贮藏量最高。锐齿槲栎和短柄枹栎种子的营养成分差别很小,但锐齿槲栎的重量和体积较短柄枹栎大,尽管二者的4种命运动态差别不大,但仍能反映出鼠类对锐齿槲栎的喜好程度高于短柄枹栎。以上这些结果表明鼠类更倾向于扩散和贮藏营养价值较高的种子,这些种子在单位时间内会让鼠类获得更多的回报,这一结果与现有的一些同类研究结果类似[4- 6]。
被捕食风险也是影响鼠类抉择的因素之一,一般情况下鼠类咬开较厚的种壳需要花费较多的时间,增加了取食风险,因而鼠类可能倾向于将种壳较厚的种子贮藏或者带到较安全的地方取食[6],栓皮栎种壳厚度较大、质地较硬,可能也是造成鼠类更多的对其进行贮藏的原因之一。实验期间笔者发现一些大石头底部的凹陷区是鼠类较为喜欢的取食地,这一现象说明鼠类在捕食种子期间也会考虑被捕食风险。
本研究发现种子扩散的速度、扩散取食量以及贮藏量在年际间存在显著的差异。2011年种子丰富度较大,鼠类捕获率较低。而2012年种子丰富度较低,鼠类捕获率很高。种子丰富度和鼠类密度的对比在一定程度上反映了单只个体的食物占有量,可以视作食物相对丰富度。由此可以看出,2011年的食物相对丰富度较高,鼠类对实验种子的消耗速率较慢,种子的贮藏量较高。而2012年的食物相对丰富度较低,鼠类对实验种子的消耗速率极高,在实验第10天的时候3种植物种子均已完全被鼠类取食,仅少量种子被贮藏。这一结果表明在食物相对丰富度较低的时期,鼠类为了保证自身能量的需求,会以取食种子为主,而食物相对丰富度较高的时期,鼠类可以在保证其活动能量需求的情况下对种子进行较多的分散贮藏,从而使种子存活和更新的可能性增加,该结论与该地区的同类研究结果一致[12]。
在本研究中,鼠类对不同种子的扩散距离也有所差异。对营养价值相对较高的栓皮栎种子的扩散(取食和贮藏)距离最远,对营养价值相对最低的短柄枹栎种子的扩散距离则最近,这与最优贮藏空间模型的预测是相一致的,即鼠类更倾向于把高价值的食物贮藏在远离种子释放点的地方[24- 25]。本实验结果也表明食物相对丰富度会对鼠类扩散种子的距离产生显著影响。2012年3种植物种子被贮藏的距离均大于2011年,且随着种子的营养价值越大,距离的增量也越大,这个结果暗示在食物匮乏的年份种子的价值量得到了提升。
在本研究中,栓皮栎种子的贮藏量最大,在实验后期被贮藏后的存留量也最大。一般认为鼠类在壳斗科植物的扩散与分布中扮演着重要的角色,据此推断栓皮栎应在该地区分布较广,但锐齿槲栎却是实验地分布最为广泛的壳斗科植物。锐齿槲栎自第三纪冰川纪以来,就在秦岭地区广泛分布,在众多地区自生为纯林或与栓皮栎、短柄枹栎等生成混交林[19]。这说明虽然种子扩散对于植物的分布很重要,但其仅是植物生存与分布的第一步,影响植物存活的因素还有很多。
本研究结果表明,鼠类倾向于贮藏营养价值高的种子。在食物相对丰富度较高的年份会更多的贮藏种子,但种子被贮藏的距离较近,在食物丰富度较低的年份会更多的取食种子。鼠类通过对种子价值的判断而采取不同的选择策略,从而影响不同种子的命运,并对这些植物种子的扩散和自然更新产生重要影响
[1] Vander Wall S B. Food Hoarding in Animals. Chicago: University of Chicago Press, 1990.
[2] Chang G, Zhang Z B. Differences in hoarding behaviors among six sympatric rodent species on seeds of oil tea (Camelliaoleifera) in Southwest China. Acta Oecologica, 2011, 37(3): 165- 169.
[3] Forget P M, Vander Wall S B. Scatter-hoarding rodents and marsupials: convergent evolution on diverging continents. Trends in Ecology & Evolution, 2001, 16(2): 65- 67.
[4] Xiao Z S, Zhang Z B, Wang Y S. Effects of seed size on dispersal distance in five rodent-dispersed fagaceous species. Acta Oecologica, 2005, 28(3): 221- 229.
[5] Perea R, Miguel A S, Gil L. Flying vs. climbing: Factors controlling arboreal seed removal in oak-beech forests. Forest Ecology and Management, 2011, 262(7): 1251- 1257.
[6] Rusch U D, Midgley J J, Anderson B. Rodent consumption and caching behaviour selects for specific seed traits. South African Journal of Botany, 2013, 84: 83- 87.
[7] Zhang H M, Zhang Z B. Endocarp thickness affects seed removal speed by small rodents in a warm-temperate broad-leafed deciduous forest, China. Acta Oecologica, 2008, 34(3): 285- 293.
[8] Xiao Z S, Zhang Z B. Nut predation and dispersal of Harland TanoakLithocarpusharlandiiby scatter-hoarding rodents. Acta Oecologica, 2006, 29(2): 205- 213.
[9] Ancillotto L, Sozio G, Mortelliti A. Acorns were good until tannins were found: Factors affecting seed-selection in the hazel dormouse (Muscardinusavellanarius). Mammalian Biology-Zeitschrifr für Säugetierkunde, 2015, 80(2): 135- 140.
[10] Yi X F, Liu G Q, Steele M A, Shen Z, Liu C Q. Directed seed dispersal by a scatter-hoarding rodent: the effects of soil water content. Animal Behaviour, 2013, 86(4): 851- 857.
[11] Chang G, Zhang Z B. Functional traits determine formation of mutualism and predation interactions in seed-rodent dispersal system of a subtropical forest. Acta Oecologica, 2014, 55: 43- 50.
[12] 张博, 石子俊, 陈晓宁, 廉振民, 常罡. 昆虫蛀蚀对鼠类介导下的锐齿槲栎种子扩散的影响. 生态学报, 2014, 34(14): 3937- 3943.
[13] Yu F, Shi X X, Wang D X, Yi X F, Fan D F, Guo T D, Lou Y G. Effects of insect infestation onQuercusalienavar.acuteserrataacorn dispersal in the Qinling Mountains, China. New Forests, 2015, 46(1): 51- 61.
[14] Rusch U D, Midgley J J, Anderson B. Seasonal fluctuations in rodent seed caching and consumption behaviour in fynbos shrublands: Implications for fire management. South African Journal of Botany, 2014, 93: 217- 221.
[15] 中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1998.
[16] 中国科学院西北植物研究所. 秦岭植物志. 北京: 科学出版社, 1974.
[17] 常罡, 王开锋, 王智. 秦岭森林鼠类对华山松种子捕食及其扩散的影响. 生态学报, 2012, 32(10): 3177- 3181.
[18] 刘诗峰, 张坚. 佛坪自然保护区生物多样性研究与保护. 西安: 陕西科学技术出版社, 2003.
[19] 岳明, 任毅, 党高弟, 辜天琪. 佛坪国家级自然保护区植物群落物种多样性特征. 生物多样性, 1999, 7(4): 263- 269.
[20] 张凤臣, 杨兴中, 巩会生. 陕西佛坪自然保护区啮齿动物群落结构研究. 野生动物, 2007, 28(1): 13- 17.
[21] 常罡. 鼠类扩散种子的几种标签标记法的比较. 生态学杂志, 2012, 31(3): 684- 688.
[22] 石子俊, 张博, 陈晓宁, 廉振民, 常罡. 秦岭壳斗科四种植物种子雨组成及其动态变化. 广西植物, 2014, 34(1): 51- 55.
[23] 常罡, 邰发道. 季节变化对锐齿栎种子扩散的影响. 生态学杂志, 2011, 30(1): 189- 192.
[24] Smith C C, Reichman O J. The evolution of food caching by birds and mammals. Annual Review of Ecology and Systematics, 1984, 15(1): 329- 351.
[25] Stapanian M A, Smith C C. A model for seed scatter hoarding: coevolution of fox squirrels and black walnuts. Ecology, 1978, 59(5): 884- 898.
Seed dispersal of three sympatric oak species by forest rodents in the south slope of Qinling Mountains, China
ZHANG Bo1, SHI Zijun1, CHEN Xiaoning2, HOU Xiang2, WANG Jing2, LI Jingang1, CHANG Gang2,*
1CollegeofLifeSciences,ShaanxiNormalUniversity,Xi′an710062,China2ShaanxiInstituteofZoology,Xi′an710032,China
Forest rodents, as seed predators and dispersers, play important roles in the regeneration of plants. Seed characteristics and food abundance influence the eating and hoarding strategies of rodents. In order to understand the seed selection differences by rodents, in 2011 and 2012 three sympatric Fagaceae plants (Quercusalienavar.acuteserrata,Q.variabilisandQ.serratavar.brevipetiolata) were used to study seed dispersal in Foping National Nature Reserve,located in the Qinling Mountains, China. Prior to the experiment, rodent density was surveyed from August to October of each year. Cage traps (27.0 cm×14.0 cm×14.0 cm), baited with peanuts, were set 10 m apart along two transects in each plot and were deployed from 17:00 to 17:30 daily. Traps were checked after 14 hours and all captured rodents were processed and immediately released. The seed mast was also surveyed from mid-August to late November of each year. 36 collection boxes were set up under 36 trees (12 trees of each species). The collection boxes were approximately 1 m2and were monitored every 7 days. Physical properties of the collected seeds were measured by technicians in the lab and chemical properties were determined by the Center of Grain Quality at the Ministry of Agriculture, China. Seed dispersal was surveyed in a deciduous broadleaved forest each year. Ten sites (1 m×1 m) were established as seed stations with a minimum distance of 15 m from one another. 20 tagged seeds of each of each of the three tree species (n=60) were placed at each station. Seed counts at each box occurred on days 1, 3, 5, 10, 17, 27 and 50 following presentation. Missing seeds were located by visual search and their fates followed. The results showed that: (1) rodents preferred to cacheQ.variabilisseeds (with higher energy) at a greater distance, and eatQ.serratavar.brevipetiolata(with lower energy) in situ.Q.variabiliswere cached farther than Q.variabilis(2011:1.52 m,2012:4.03 m). All three seed types were cached further when food was scarce. (2) In a mast seeding year (2011), rodents consumed seeds slowly and cached more seeds. 67.33% seeds remained at each station on the 10thday, and 29.67% seeds remained in caches on the 50thday. In the year that food resources were relatively poor, rodents tended to eat more seeds. All seeds were removed by the 10thday and only 3.83% seeds remained in the caches on the 50thday. When food was most scarce, all three kinds of seeds were cached less. (3) Although moreQ.variabilisseeds cached by rodents survived in later experiments, it had a narrow range of distribution. This phenomenon indicates that seed dispersal is just the first step of plant distribution and survival. These results indicate that rodents primarily select seeds of high nutritional value to cache. More seeds were cached in the year with abundant food resources and at closer cache distances, while more seeds were consumed when food resources were relatively poor.
seed traits; hoarding behavior; relative food abundance; seed dispersal; seed selection; seed consumption
国家自然科学基金项目(31100283);陕西省自然科学基金项目(2014JM3066);“西部之光”人才培养计划项目(2012DF04)
2015- 03- 05;
日期:2016- 03- 03
10.5846/stxb201503050428
*通讯作者Corresponding author.E-mail: snow1178@snnu.edu.cn
张博,石子俊,陈晓宁,侯祥,王京,李金钢,常罡.森林鼠类对秦岭南坡3种壳斗科植物种子扩散的影响.生态学报,2016,36(21):6750- 6757.
Zhang B, Shi Z J, Chen X N, Hou X, Wang J, Li J G, Chang G.Seed dispersal of three sympatric oak species by forest rodents in the south slope of Qinling Mountains, China.Acta Ecologica Sinica,2016,36(21):6750- 6757.
