苗龄、光照强度和施氮量对抗褐飞虱水稻品种主要防御酶活性的影响
2016-12-27吴碧球孙祖雄黄所生周君雷黄凤宽蒋显斌龙丽萍
吴碧球, 李 成, 孙祖雄,黄所生, 周君雷, 黄凤宽*,凌 炎, 蒋显斌, 黄 芊, 龙丽萍
(1. 广西农业科学院植物保护研究所, 广西作物病虫害生物学重点实验室, 南宁 530007;2. 广西防城港市植保站, 防城港 538001;3. 广西大学农学院, 南宁 530005; 4. 广西农业科学院水稻研究所, 南宁 530007)
苗龄、光照强度和施氮量对抗褐飞虱水稻品种主要防御酶活性的影响
吴碧球1, 李 成1, 孙祖雄2,黄所生1, 周君雷3, 黄凤宽1*,凌 炎1, 蒋显斌4, 黄 芊4, 龙丽萍4
(1. 广西农业科学院植物保护研究所, 广西作物病虫害生物学重点实验室, 南宁 530007;2. 广西防城港市植保站, 防城港 538001;3. 广西大学农学院, 南宁 530005; 4. 广西农业科学院水稻研究所, 南宁 530007)
为探明苗龄、光照强度和施氮量对水稻主要防御酶活性的影响及其与抗褐飞虱的关系,分别就苗龄、光照强度和施氮量等因子对不同抗褐飞虱水稻品种苯丙氨酸解氨酶(PAL)、多酚氧化酶(PPO)和过氧化物酶(POD)活性的影响进行研究。结果表明:IR56、Ptb33和570011对褐飞虱生物型II的抗性与PAL和PPO活性密切相关,RHT对II型的抗性与PAL、PPO和POD活性密切相关,RP1976-18-6-4-2对II型的抗性与PAL活性密切相关,570011对II型的抗性与PAL和PPO活性密切相关;IR56对褐飞虱孟加拉型的抗性与PAL、PPO和POD活性密切相关,Rathu Heenati(RHT)和RP1976-18-6-4-2对孟加拉型的抗性与与PPO密切相关,570011对孟加拉型的抗性与POD活性密切相关。苗龄、光照强度和施氮量的单独作用或交互作用对不同水稻中防御酶活性有明显影响,但品种不同,其影响程度也不同。苗龄、光照强度和施氮量单独或是交互作用显著影响IR56中PAL和PPO、RHT和RP1976-18-6-4-2中PPO、Ptb33中POD及570011中PAL和POD的活性,苗龄显著影响IR56和 RP1976-18-6-4-2中POD的活性,光照强度和施氮量单独或交互作用显著影响RHT和RP1976-18-6-4-2中PAL的活性,施氮量显著影响RHT中POD的活性,光照强度显著影响Ptb33中PAL的活性;苗龄和光照强度单独或交互作用显著影响570011中PPO的活性。
苗龄; 光照强度; 施氮量; 抗虫水稻; 防御酶; 褐飞虱
在植物与昆虫长期协同进化过程中,形成了许多适应及防御机制。过氧化物酶(POD)和过氧化氢酶(CAT)等是植物体内重要的保护性酶系,多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)等是植物体内主要防御性酶系,这些蛋白酶的活性与植物抗虫性密切相关 (Krishnan & Kodrík, 2006; Huangetal., 2007; 史瑞等, 2008; 常金华等, 2008; 严善春等, 2011; 张金等, 2014)。PAL、PPO和POD活性的激活是植物细胞应对生物或非生物胁迫的一种响应,在植物抗逆反应中起着十分重要的作用 (段灿星等, 2012; 严俊鑫等, 2013; 张凯等, 2014; 冯定邦等, 2015),是植物防御次生物质生成过程中的关键酶 (Mauch-Mani & Slusarenkl, 1996; 鲁艺芳等, 2012)。
褐飞虱是我国及亚洲国家当前水稻上主要的迁飞性害虫,该虫通过吸食水稻汁液、产卵为害及传播水稻病害3种方式为害水稻,导致稻株生长缓慢,产量损失,严重时造成“穿顶”,严重威胁我国粮食生产安全。抗虫品种的利用是控制褐飞虱最为经济有效的措施。但水稻抗虫性易受自身生育期(苗龄)土壤肥力、栽培措施、生物因素等因素影响,有研究表明,苗龄、光照强度和施氮量对水稻品种抗褐飞虱有显著影响,苗龄过小、过度遮光或偏施氮肥均导致抗虫水稻品种对褐飞虱的抗性下降,甚至丧失,加速稻株死亡 (刘春茂和吴荣宗, 1992; 吴碧球等, 2004, 2009; 黄凤宽等, 2005)。而植物中与抗虫性关系密切的防御酶PAL、PPO和保护性酶POD也易受光照、温度、水分等许多因子的影响 (杨振国和侯凤莲, 1997),研究发现光一方面可以启动和激活玉米叶片PAL活性,另一方面可以促进PAL I的合成;低光条件下可以提高满江红耐荫品系生物量与体内PPO活性,使其抵御病虫入侵能力提高,保护生理代谢功能不被破坏,从而保持了较正常的生长能力 (唐龙飞等, 1999);不同氮、磷施肥处理对红三叶PAL活性的影响极其显著 (狄浩等, 2013),施硅肥可提高棉花PPO和PAL的活性,降低POD活性 (李卫东和周泉佚, 2015);灰飞虱为害后,抗感水稻植株体内的PAL和POD活性随灰飞虱取食时间延长而显著升高,PPO活性则表现先升后降趋势,但在抗虫品种Mudgo中3种防御酶活性明显高于感虫品种 (段灿星等, 2012)。但关于气候变化、土壤肥力、栽培措施、生物因素等对抗褐飞虱水稻植株内防御酶影响的研究多集中在虫害影响方面 (陈建明等, 2003; 张金锋和薛庆中, 2004; 刘欲强等, 2005),而苗龄、光照强度和施氮量对抗褐飞虱水稻品种内PAL、PPO和POD活性的影响,尚未见报道。本文采用均匀设计法研究在不同苗龄、光照强度和施氮量条件下,水稻品种对褐飞虱的抗性表现及水稻植株内防御酶PAL、PPO和POD活性的变化,旨在明确苗龄、光照强度和施氮量对抗虫水稻品种防御酶活性的影响及其与抗褐飞虱的关系,以期为抗褐飞虱育种提供稳定性好的亲本,并为褐飞虱防控提供科学依据。
1 材料与方法
1.1 材料
供试水稻品种:感虫品种TN1,选择经苗期抗虫性鉴定的抗褐飞虱褐飞虱生物型Ⅱ和孟加拉型水稻品种为IR56、RHT、Ptb33、RP1976-18-6-4-2和570011。其中,对褐飞虱生物型Ⅱ的抗性,IR56、RHT、Ptb33、RP1976-18-6-4-2、570011分别表现为中抗、抗、抗、抗、抗、抗;对褐飞虱孟加拉型的抗性,IR56、RHT、Ptb33、RP1976-18-6-4-2、570011分别表现为中抗、中抗、中抗、抗、中抗、抗。上述供试品种中仅570011来源于广西品种资源,其余品种均来自于国际水稻研究所。
供试虫源:褐飞虱为广西南宁田间采集怀卵雌成虫进行个体培养建立种群,然后分别用含不同抗性基因的褐飞虱鉴别品种进行苗期抗性测定,根据鉴别品种的抗性表现确定褐飞虱的生物型,将分离的褐飞虱生物II和孟加拉型分别在Mudgo和IR36上饲养扩繁8代后再进行试验。
试剂:PAL、PPO、POD试剂盒,苏州科铭生物技术有限公司生产。
仪器:TES-1339照度计,台湾泰仕电子工业股份有限公司;TGL-16台式高速冷冻离心机,湘仪离心机仪器有限公司;UV-5800紫外可见分光光度计,上海元析仪器有限公司。
1.2 试验设计
本研究采用均匀设计法研究苗龄(A)、光照强度(B)和施氮量(C) 3个因子对抗褐飞虱水稻品种营养物质的影响。所选因子及水平见表1。本试验中A因子设3个水平:8 d、13 d、18 d。B因子设3个水平:0层纱网遮光(未遮光,自然光照强度)、2层纱网遮光、4层纱网遮光;后2个水平的光照强度分别为自然光强的50.33%和27.38%。C因子设3个水平:未施肥(0 Kg/hm2尿素)、225 Kg/hm2尿素、450 Kg/hm2尿素(氮肥含氮量为46.4%,广西河池化工股份有限公司生产)。试验设计采用U6(63),见表2。
1.3 水稻品种抗褐飞虱鉴定
本试验于2014年6-9月在广西农业科学院植物保护研究所玻璃网室进行。按表2的试验方案实施。将供试水稻品种分期播于铝托(52 cm×36.5 cm×6.5 cm)内,每盆播12行,每品种播两行,每行留20苗。每试验处理播10盆。当苗龄达到试验要求时,提前1 d施肥,第二天按每苗接入褐飞虱生物型II和孟加拉型1-2龄若虫5头,然后进行遮光处理。待感虫对照TN1枯萎后7-10 d参照国际使用统一标准进行逐株定级,最后计算各品种的平均受害级别。
1.4 防御酶活性测定
1.4.1 稻样采集
将供试水稻按照1.3方法处理7 d时剪取心叶以下第一、二叶鲜样,用蒸馏水擦洗,然后置于-20℃冰箱内保存备用。
1.4.2 苯丙氨酸解氨酶PAL活性测定
称取0.1 g稻叶鲜样,加入1 mL提取液进行冰浴匀浆。10000 g 4℃离心10 min,取上清液,置冰上待测。按照苏州科铭生物技术有限公司生产的PAL试剂盒说明依次加入试剂,混匀,静置10 min后,用对照管调零(以不加样本为对照管),在290 nm处记录测定吸光值A。PAL活性计算公式如下:
表1 苗龄、光照强度和施氮量对水稻主要防御活性影响的水平设计
Table 1 Level design of effect of seedling stage, light intensity and N application rate on activity of main defense enzyme in rice planting

水平Level苗龄(A)Seedlingstage(d)光照强度(B)Lightintensity(纱网层数Screenlayers)施氮量(C)(kg/hm2)Napplicationrate180021322253184450
表2 苗龄、光照强度和施氮量对水稻主要防御活性影响的U6(63)水平试验方案
Table 2 U6(63) test program of effect of seedling stage, light intensity and N application rate on activity of main defense enzyme in rice planting
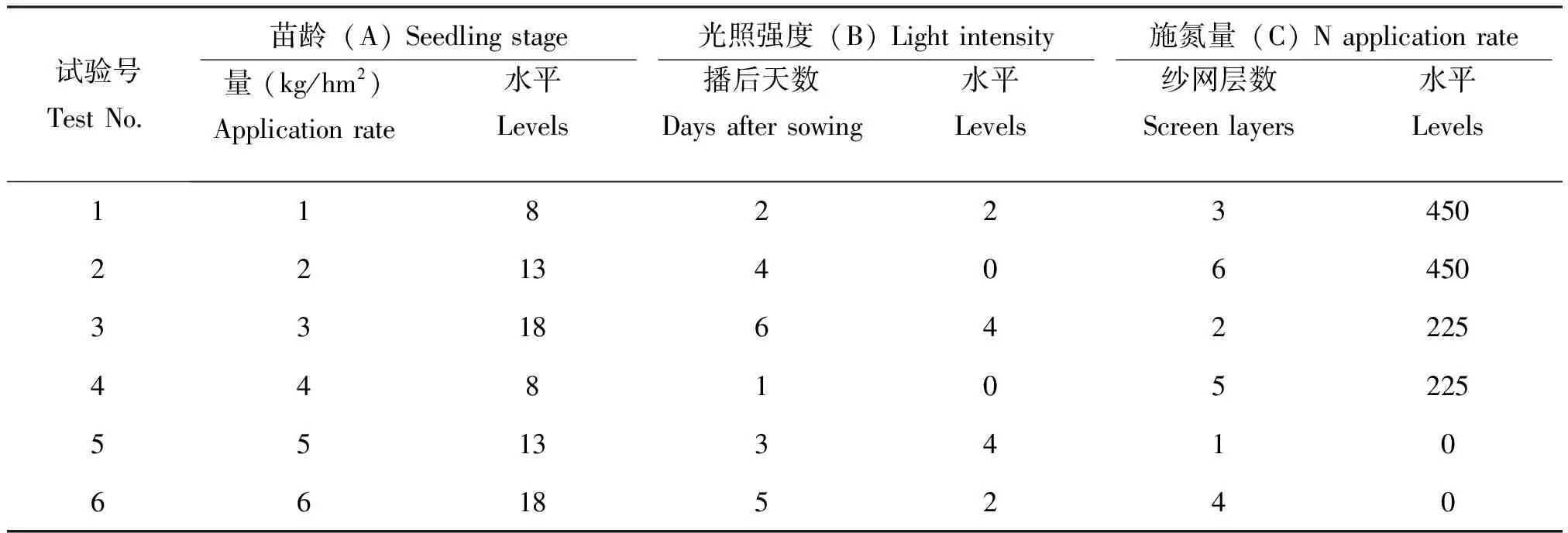
试验号TestNo 苗龄(A)Seedlingstage 光照强度(B)Lightintensity 施氮量(C)Napplicationrate 水平Levels播后天数Daysaftersowing水平Levels纱网层数Screenlayers水平Levels量(kg/hm2)Applicationrate118223450221340645033186422254481052255513341066185240
PAL(U/g 鲜重)= 反应总体积÷样本体积÷反应时间÷0.1×A÷样本鲜重(g/mL)
1.4.3 多酚氧化酶PPO活性测定
称取0.1 g稻叶鲜样,加入1 mL提取液进行冰浴匀浆。8000 g 4℃离心10 min,取上清液,置冰上待测。按照苏州科铭生物技术有限公司生产的PPO试剂盒说明依次加入试剂,充分混匀,5000 g 常温离心10 min,收集上清液,用蒸馏水调零,煮沸样本为对照管,于525 nm处检测测定管和对照管吸光度。PPO活性计算公式如下:
PPO(U/g 鲜重)=反应总体积÷样本体积÷反应时间÷0.01×Δ(A测定-A对照)÷样本鲜重(g/mL)
1.4.4 过氧化物酶POD活性测定
称取0.1 g稻叶鲜样,加入1 mL提取液进行冰浴匀浆。8000 g 4℃离心10 min,取上清液,置冰上待测。按照苏州科铭生物技术有限公司生产的POD试剂盒说明依次加入试剂,然后加入样本后立即混匀并计时,记录470 nm 处30 s时的吸光值A1 和1 min 30 s后的吸光值A2。计算ΔA=A2-A1。POD活性计算公式如下:
PPO(U/g 鲜重)=反应总体积÷样本体积÷0.01×ΔA÷样本质量(g/mL)
1.5 数据处理
采用Microsoft Excel和SPSS 15.0软件处理数据,并用Duncan新复极差法进行差异显著性比较(α=0.05)。
2 结果与分析
2.1 苗龄、光照强度和施氮量影响下水稻品种防御酶活性与其抗褐飞虱关系
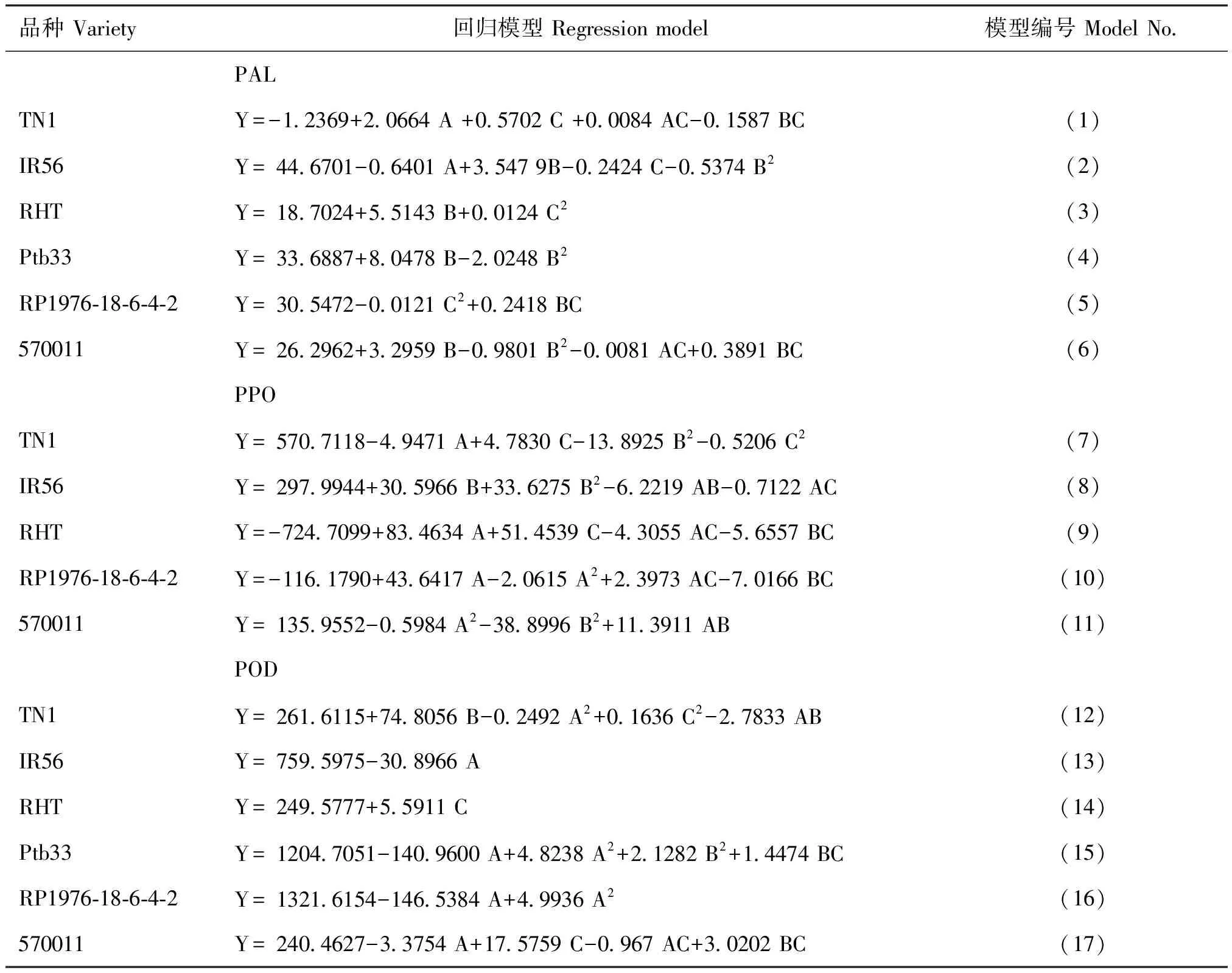
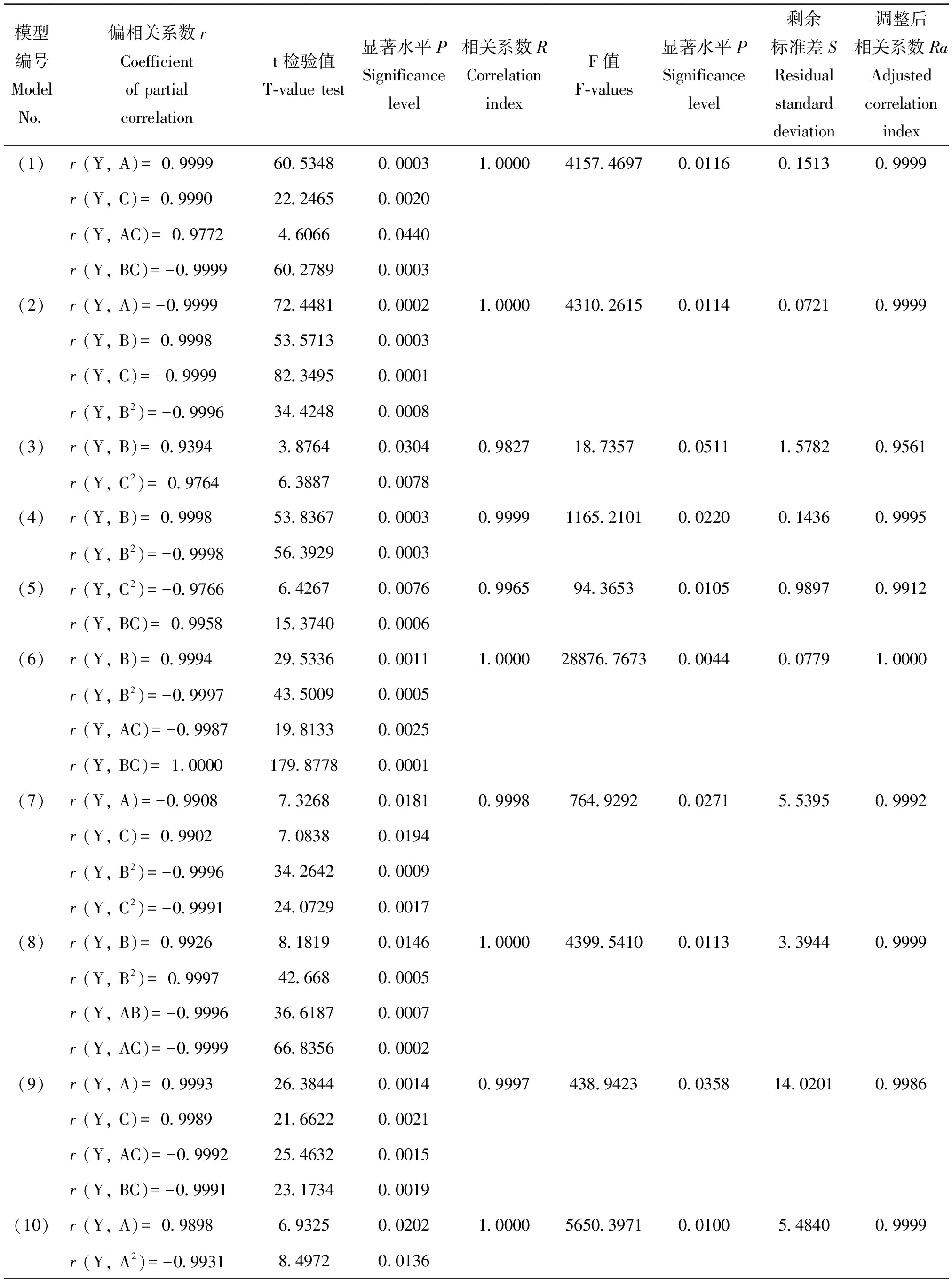
将均匀设计试验中各水稻品种防御酶活性及受害级别的试验数据进行二项式逐步回归,得到不同水稻品种受害级别(Y)与苯丙氨酸解氨酶PAL(A)、多酚氧化酶PPO(B)和过氧化物酶POD(C)的回归关系式,通过对回归模型进行检验,检验达到显著或极显著水平的模型(表3)。模型的检验结果如表4。由表4可知,模型(1)-模型(9)各因素均达显著或极显著水平。因而采用模型(1)-(9)可以很好地描述稻株中防御酶活性变化与IR56、RHT、Ptb33、RP1976-18-6-4-2和570011抗褐飞虱生物型II和孟加拉型的关系。
在苗龄、光照强度和施氮量的影响下,模型(1)可知,IR56受褐飞虱生物型Ⅱ为害的受害级别与PAL和PPO单独作用极显著负相关,与两者交互作用极显著正相关,即IR56对褐飞虱生物型Ⅱ的抗性与PAL和PPO单独作用极显著负相关,与两者交互作用极显著负相关。模型(2)可知,RHT受褐飞虱生物型Ⅱ为害的受害级别与PAL、POD和PPO平方项单独作用极显著正相关,与POD平方项作用极显著负相关,即RHT对褐飞虱生物型Ⅱ的抗性与POD、PAL和PPO平方项单独作用极显著负相关,与POD平方项作用极显著正相关。模型(3)可知,Ptb33受褐飞虱生物型Ⅱ为害的受害级别与PPO单独作用极显著负相关,与PAL和PPO交互作用极显著正相关,即Ptb33对褐飞虱生物型Ⅱ的抗性与PPO单独作用极显著正相关,与PAL和PPO交互作用极显著负相关。模型(4)可知,RP1976-18-6-4-2受褐飞虱生物型Ⅱ为害的受害级别与PAL单独作用显著正相关,即RP1976-18-6-4-2对褐飞虱生物型Ⅱ的抗性仅与PAL单独作用显著负相关。模型(5)可知,570011受褐飞虱生物型Ⅱ为害的受害级别与PAL单独作用极显著正相关,与PPO单独作用显著负相关,即570011对褐飞虱生物型Ⅱ的抗性与PAL单独作用极显著负相关,与PPO单独作用显著正相关。模型(6)可知,IR56受褐飞虱孟加拉型为害的受害级别与PAL单独作用负相关,与PPO和POD单独作用显著或极显著正相关,即IR56对褐飞虱孟加拉型的抗性与PAL单独作用显著正相关,与PPO和POD单独作用显著或极显著负相关。模型(7)可知,RHT受褐飞虱孟加拉型为害的受害级别与PPO单独作用极显著负相关,即RHT对褐飞虱孟加拉型的抗性仅与PPO单独作用极显著正相关。模型(8)可知,RP1976-18-6-4-2受褐飞虱孟加拉型为害的受害级别与PPO单独作用显著正相关,与PPO平方项作用显著负相关,即RP1976-18-6-4-2对褐飞虱孟加拉型的抗性与PPO单独作用显著负相关,与PPO平方项作用显著正相关。模型(9)可知,570011受褐飞虱孟加拉型为害的受害级别与POD单独作用极显著正相关,与POD平方项作用极显著负相关,即570011对褐飞虱孟加拉型的抗性与POD单独作用极显著负相关,与POD平方项作用极显著正相关。
2.2 苗龄、光照强度、施氮量对水稻品种防御酶活性的影响
苗龄、光照强度和施氮量对水稻品种中苯丙氨酸解氨酶(PAL)、多酚氧化酶(PPO)和过氧化物酶(POD)活性有影响,但影响程度因处理水平和品种而异(见表5)。由表5可知,TN1 PAL活性在试验号2(13 d苗龄、未遮光、450 kg/hm2)最高,显著高于其余试验号;在试验号4(8 d苗龄、未遮光、225 kg/hm2)最低,但与试验号5(13 d苗龄、4层纱网遮光、未施氮)差异不显著。TN1 PPO活性在试验号4最高,但与试验号6(18 d苗龄、2层纱网遮光、未施氮)差异不显著;在试验号1(8 d苗龄、2层纱网遮光、450 kg/hm2)最低,仅显著低于试验号4、5和6。TN1 POD活性在试验号1最高,显著高于试验号4和6;在试验号6最低,但仅显著低于试验号1。
IR56 PAL活性在试验号5最高,但仅显著高于试验号2;在试验号2最低,但仅显著低于试验号5。IR56 PPO活性在试验号5最高,显著高于其余试验号;在试验号2最低,显著低于其余试验号。IR56 POD活性在试验号1最高,显著高于试验号2、3、5和6;在试验号6最低,但与试验号5差异不显著。
表3 水稻品种防御酶活性与其受害级别关系
Table 3 Relationship between the defense enzyme activity of rice varieties and damaged level

品种Variety回归模型Regressionmodel模型编号ModelNo褐飞虱生物型ⅡBPHbiotypeⅡIR56Y=3348996004-116712330A-11010973B+471844AB(1)RHTY=-231386+01625C+00004A2+00000007B2-00002C2(2)Ptb33Y=94884-00812B+00019AB(3)RP1976⁃18⁃6⁃4⁃2Y=-42138+02953A(4)570011Y=-15340+02221A-00046B(5)褐飞虱孟加拉型BPHbiotypeBangladeshIR56Y=143489-02465A+00053B+00047C(6)RHTY=94060-00066B(7)RP1976⁃18⁃6⁃4⁃2Y=48264+00180B-000001B2(8)570011Y=-56499+00608C-000006C2(9)
表4 模型的检验结果
Table 4 Inspection result of model
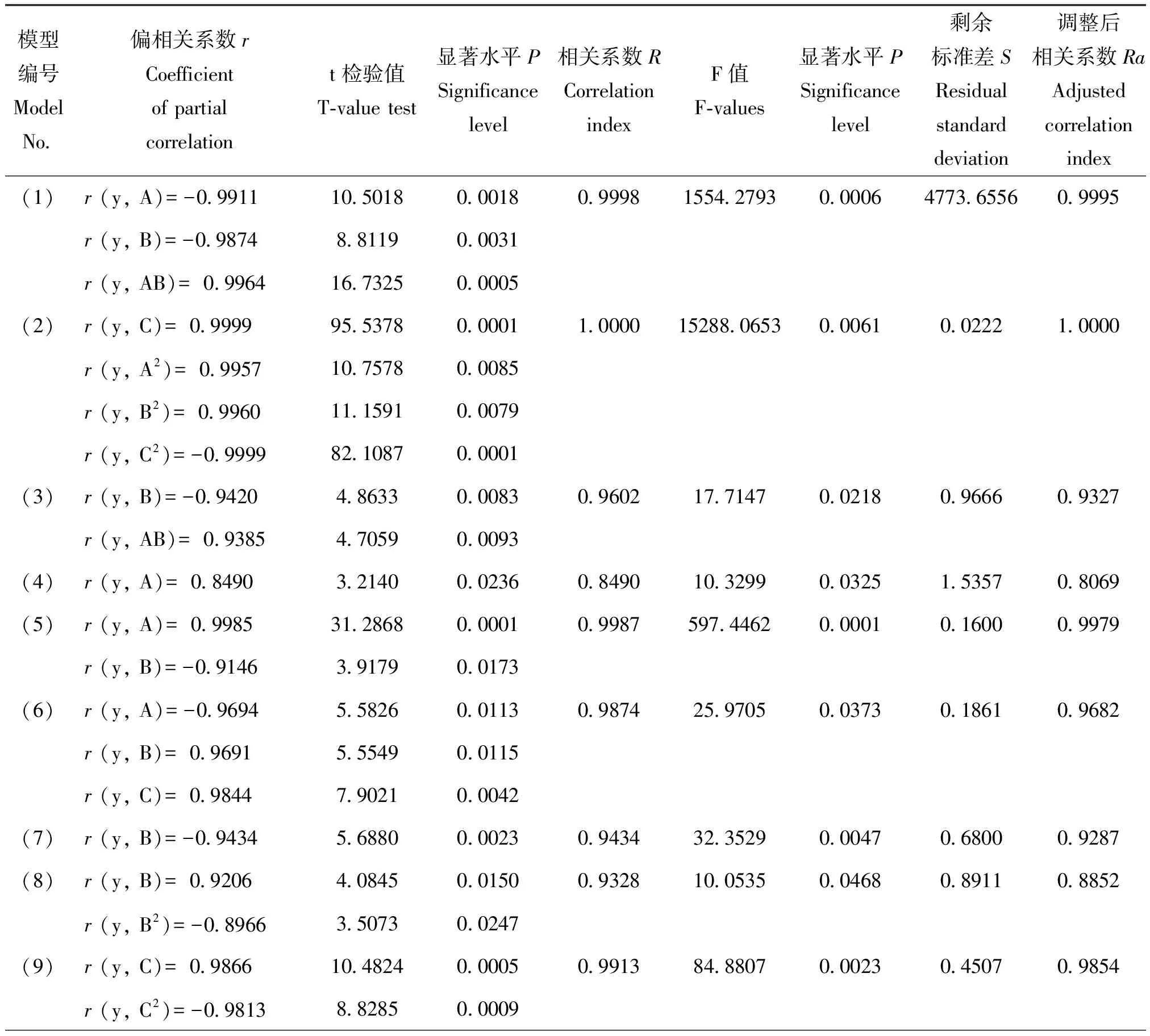
模型编号ModelNo偏相关系数rCoefficientofpartialcorrelationt检验值T⁃valuetest显著水平PSignificancelevel相关系数RCorrelationindexF值F⁃values显著水平PSignificancelevel剩余标准差SResidualstandarddeviation调整后相关系数RaAdjustedcorrelationindex(1)r(y,A)=-09911105018000180999815542793000064773655609995r(y,B)=-098748811900031r(y,AB)=0996416732500005(2)r(y,C)=099999553780000110000152880653000610022210000r(y,A2)=0995710757800085r(y,B2)=0996011159100079r(y,C2)=-0999982108700001(3)r(y,B)=-09420486330008309602177147002180966609327r(y,AB)=093854705900093(4)r(y,A)=08490321400023608490103299003251535708069(5)r(y,A)=0998531286800001099875974462000010160009979r(y,B)=-091463917900173(6)r(y,A)=-09694558260011309874259705003730186109682r(y,B)=096915554900115r(y,C)=098447902100042(7)r(y,B)=-09434568800002309434323529000470680009287(8)r(y,B)=09206408450015009328100535004680891108852r(y,B2)=-089663507300247(9)r(y,C)=098661048240000509913848807000230450709854r(y,C2)=-098138828500009
RHT PAL活性在试验号1最高,但仅显著高于活性最低的试验号4,在试验号2、3、5和6间的活性差异不显著。RHT PPO活性在试验号6最高,显著高于其余试验号;在试验号3最低,显著低于其余试验号,且各试验号间均差异显著。RHT POD活性在试验号4最高,显著高于试验号3、5和6;在试验号5最低,但与试验号6和3差异不显著。
Ptb33 PAL活性在所有试验号间差异均不显著。Ptb33 PPO活性在试验号6最高,显著高于其余试验号;在试验号2最低,显著低于试验号1、4、5和6,而与试验号3差异不显著。Ptb33 POD活性在试验号1最高,但与试验号3和4差异不显著;在试验号2最低,但仅显著低于试验号1和4。
RP1976-18-6-4-2 PAL活性在试验号3最高,显著高于其余试验号;在试验号2最低,显著低于其余试验号。RP1976-18-6-4-2 PPO活性在试验号2最高,显著高于其余试验号;在试验号6最低,显著低于试验号1、2、3、4,但与试验号5差异不显著。RP1976-18-6-4-2 POD活性在试验号4最高,显著高于试验号2、3、5和6,而与试验号1差异不显著;在试验号5最低,但与试验号2和6差异不显著。
570011 PAL活性在试验号1最高,显著高于试验号2、4、5和6,而与试验号3差异不显著;在试验号2最低,但仅显著低于试验号1和3。570011 PPO活性在试验号6最高,显著高于其余试验号;在试验号5最低,显著低于试验号1、3、4和6,而与试验号2差异不显著。570011 POD活性在试验号1最高,显著高于其余试验号,在试验号6最低,但与试验号5差异不显著。
表5 苗龄、光照强度和施氮量对水稻防御酶活性的影响 (U/g)
Table 5 Effect of seedling stage, light intensity and N application rate on defense enzyme activity of rice
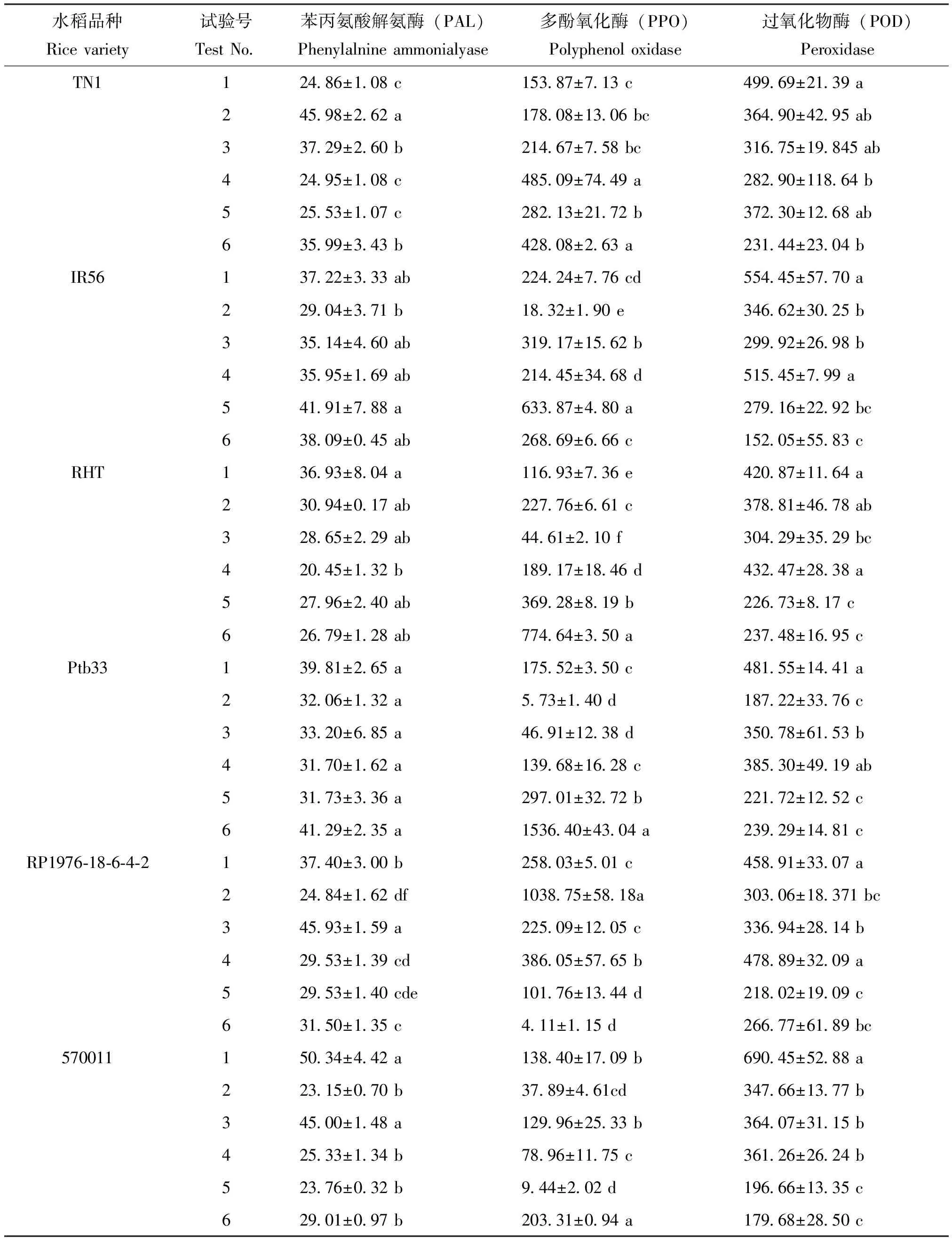
水稻品种Ricevariety试验号TestNo苯丙氨酸解氨酶(PAL)Phenylalnineammonialyase多酚氧化酶(PPO)Polyphenoloxidase过氧化物酶(POD)PeroxidaseTN112486±108c15387±713c49969±2139a24598±262a17808±1306bc36490±4295ab33729±260b21467±758bc31675±19845ab42495±108c48509±7449a28290±11864b52553±107c28213±2172b37230±1268ab63599±343b42808±263a23144±2304bIR5613722±333ab22424±776cd55445±5770a22904±371b1832±190e34662±3025b33514±460ab31917±1562b29992±2698b43595±169ab21445±3468d51545±799a54191±788a63387±480a27916±2292bc63809±045ab26869±666c15205±5583cRHT13693±804a11693±736e42087±1164a23094±017ab22776±661c37881±4678ab32865±229ab4461±210f30429±3529bc42045±132b18917±1846d43247±2838a52796±240ab36928±819b22673±817c62679±128ab77464±350a23748±1695cPtb3313981±265a17552±350c48155±1441a23206±132a573±140d18722±3376c33320±685a4691±1238d35078±6153b43170±162a13968±1628c38530±4919ab53173±336a29701±3272b22172±1252c64129±235a153640±4304a23929±1481cRP1976⁃18⁃6⁃4⁃213740±300b25803±501c45891±3307a22484±162df103875±5818a30306±18371bc34593±159a22509±1205c33694±2814b42953±139cd38605±5765b47889±3209a52953±140cde10176±1344d21802±1909c63150±135c411±115d26677±6189bc57001115034±442a13840±1709b69045±5288a22315±070b3789±461cd34766±1377b34500±148a12996±2533b36407±3115b42533±134b7896±1175c36126±2624b52376±032b944±202d19666±1335c62901±097b20331±094a17968±2850c
注:同列平均数后字母相同者表示未达0.05显著水平。Note: Same-column means followed by the same letter are not significantly different at the 0.05 levels.
2.3 苗龄、光照强度、施氮量对水稻品种防御酶活性的影响及其相关性分析
对表5不同水稻品种中防御性酶活性的试验数据进行二项式逐步回归,得到不同水稻品种PAL、PPO和POD活性(Y)与苗龄(A)、光照强度(B)、施氮量(C)的回归关系式,通过对回归模型进行检验,检验达到显著或极显著水平的模型(表6)。模型的检验结果如表7。由表7可知,模型(1)—模型(17)各因素均达显著或极显著水平。因而采用模型(1)—模型(17)可以很好地描述苗龄、光照强度和施氮量对TN1、IR56、RHT、Ptb33、RP1976-18-6-4-2和570011中PAL、PPO和POD活性的影响。
表6 苗龄、光照强度、施氮量与不同水稻品种防御酶活性的关系
Table 6 Relationship among defense enzyme activity of rice and seedling stage, light intensity and N application rate
品种Variety回归模型Regressionmodel模型编号ModelNoPALTN1Y=-12369+20664A+05702C+00084AC-01587BC(1)IR56Y=446701-06401A+35479B-02424C-05374B2(2)RHTY=187024+55143B+00124C2(3)Ptb33Y=336887+80478B-20248B2(4)RP1976⁃18⁃6⁃4⁃2Y=305472-00121C2+02418BC(5)570011Y=262962+32959B-09801B2-00081AC+03891BC(6)PPOTN1Y=5707118-49471A+47830C-138925B2-05206C2(7)IR56Y=2979944+305966B+336275B2-62219AB-07122AC(8)RHTY=-7247099+834634A+514539C-43055AC-56557BC(9)RP1976⁃18⁃6⁃4⁃2Y=-1161790+436417A-20615A2+23973AC-70166BC(10)570011Y=1359552-05984A2-388996B2+113911AB(11)PODTN1Y=2616115+748056B-02492A2+01636C2-27833AB(12)IR56Y=7595975-308966A(13)RHTY=2495777+55911C(14)Ptb33Y=12047051-1409600A+48238A2+21282B2+14474BC(15)RP1976⁃18⁃6⁃4⁃2Y=13216154-1465384A+49936A2(16)570011Y=2404627-33754A+175759C-0967AC+30202BC(17)
模型(1)可见,TN1 PAL活性与苗龄和施氮量单独作用及两者间交互作用显著或极显著正相关,与光照强度和施氮量交互作用极显著负相关。模型(2)可见,IR56 PAL活性与苗龄和施氮量单独作用及光照强度平方项作用极显著负相关,与光照强度单独作用极显著正相关。模型(3)可见,RHT PAL活性与光照强度单独作用及施氮量平方项作用显著或极显著正相关。模型(4)可见,Ptb33 PAL活性与光照强度单独作用极显著正相关,与光照强度平方项作用极显著负相关。模型(5)可见,RP1976-18-6-4-2 PAL活性与施氮量平方项作用极显著负相关,与光照强度和施氮量交互作用极显著正相关。模型(6)可见,570011 PAL活性与光照强度单独作用及光照强度和施氮量交互作用极显著正相关,与光照强度平方项作用及苗龄和施氮量交互作用极显著负相关。
表7 模型的检验结果
Table 7 Inspection result of model
模型编号ModelNo偏相关系数rCoefficientofpartialcorrelationt检验值T⁃valuetest显著水平PSignificancelevel相关系数RCorrelationindexF值F⁃values显著水平PSignificancelevel剩余标准差SResidualstandarddeviation调整后相关系数RaAdjustedcorrelationindex(1)r(Y,A)=09999605348000031000041574697001160151309999r(Y,C)=0999022246500020r(Y,AC)=097724606600440r(Y,BC)=-0999960278900003(2)r(Y,A)=-09999724481000021000043102615001140072109999r(Y,B)=0999853571300003r(Y,C)=-0999982349500001r(Y,B2)=-0999634424800008(3)r(Y,B)=09394387640030409827187357005111578209561r(Y,C2)=097646388700078(4)r(Y,B)=09998538367000030999911652101002200143609995r(Y,B2)=-0999856392900003(5)r(Y,C2)=-09766642670007609965943653001050989709912r(Y,BC)=0995815374000006(6)r(Y,B)=099942953360001110000288767673000440077910000r(Y,B2)=-0999743500900005r(Y,AC)=-0998719813300025r(Y,BC)=10000179877800001(7)r(Y,A)=-099087326800181099987649292002715539509992r(Y,C)=099027083800194r(Y,B2)=-0999634264200009r(Y,C2)=-0999124072900017(8)r(Y,B)=0992681819001461000043995410001133394409999r(Y,B2)=099974266800005r(Y,AB)=-0999636618700007r(Y,AC)=-0999966835600002(9)r(Y,A)=09993263844000140999743894230035814020109986r(Y,C)=0998921662200021r(Y,AC)=-0999225463200015r(Y,BC)=-0999123173400019(10)r(Y,A)=0989869325002021000056503971001005484009999r(Y,A2)=-099318497200136
续上表

模型编号ModelNo偏相关系数rCoefficientofpartialcorrelationt检验值T⁃valuetest显著水平PSignificancelevel相关系数RCorrelationindexF值F⁃values显著水平PSignificancelevel剩余标准差SResidualstandarddeviation调整后相关系数RaAdjustedcorrelationindexr(Y,AC)=10000124931100001r(Y,BC)=-0999963412800002(11)r(Y,A2)=-092543453000408098582290890042119004609640r(Y,B2)=-098117168600056r(Y,AB)=097756557800072(12)r(Y,B)=0998417687400032099999913343002383278009994r(Y,A2)=-099358751800128r(Y,C2)=0999636690300007r(Y,AB)=-09932854400134(13)r(Y,A)=-090914363200073090911903780012070811208849(14)r(Y,C)=08288296280031408288877840041456612107802(15)r(Y,A)=-1000012431530000110000180417638000560946310000r(Y,A2)=10000113984200001r(Y,B2)=0999326987200014r(Y,BC)=0999976358500002(16)r(Y,A)=-089803535200241094121163400038645741608998r(Y,A2)=084643151500076(17)r(Y,A)=-10000128349000001100001249998125000210116610000r(Y,C)=10000890052500001r(Y,AC)=-10000686242200001r(Y,BC)=100001488509400001
由模型(7)可见,TN1 PPO活性与苗龄单独作用及光照强度和施氮量平方项作用显著或极显著负相关,与施氮量单独作用显著正相关。模型(8)可见,IR56 PPO活性与光照强度单独及平方项作用显著或极显著正相关,与苗龄和光照强度及苗龄和施氮量的交互作用极显著负相关。模型(9)可见,RHT PPO活性与苗龄和施氮量的单独作用极显著正相关,与苗龄和施氮量及光照强度和施氮量的交互作用极显著负相关。模型(10)可见,RP1976-18-6-4-2 PPO活性与苗龄单独作用及苗龄和施氮量交互作用显著或极显著正相关,与苗龄平方项作用及光照强度和施氮量交互作用显著或极显著负相关。模型(11)可见,570011 PPO活性与苗龄和光照强度平方项作用显著或极显著负相关,与苗龄和光照强度交互作用极显著正相关。模型(12)可见,TN1 POD活性与光照强度单独作用及施氮量平方项作用极显著正相关,与苗龄平方项作用及苗龄和光照强度交互作用显著负相关。模型(13)可见,IR56 POD活性仅与苗龄单独作用极显著负相关。模型(14)可见,RHT POD活性仅与施氮量单独作用显著正相关。模型(15)可见,Ptb33 POD活性仅与苗龄单独作用显著负相关。模型(16)可见,RP1976-18-6-4-2 POD活性与苗龄单独作用极显著负相关,与苗龄平方项作用极显著正相关。模型(17)可见,570011 POD活性与苗龄单独作用及苗龄和施氮量交互作用极显著负相关,与施氮量单独作用及光照强度和施氮量交互作用极显著正相关。
3 结论与讨论
PAL 、PPO和POD是植物重要的防卫性酶,与植物抗虫性关系密切 (庄炳昌等, 1992; 李润植等, 1998),可作为植物抗虫育种的一个指标 (代莉等, 2003)。PAL是植物形成木质素、酚类等次生代谢物质的关键酶 (李润植等, 1998; 曾永三和王振, 1999);PPO能够催化酚类化合物转化为更高活性的醌类物质,将昆虫食物蛋白中必须氨基酸烷基化,阻碍昆虫营养的利用 (Tscharntkeetal., 2001),或与昆虫肠胃里的消化酶结合使酶失活,减缓昆虫的生长发育并促使其死亡 (王曼玲等, 2005);POD在由酚类物质聚合成木质素中起着重要作用,高活性的POD有助于植物细胞壁的强化,含丰富木质素的细胞壁对刺吸式害虫可产生一定的取食障碍 (贾贞等, 2004)。本研究发现IR56对褐飞虱生物型Ⅱ的抗性与PAL和PPO活性密切相关,对孟加拉型的抗性与PAL、PPO和POD活性密切相关;RHT对褐飞虱生物型Ⅱ的抗性与PAL、PPO和POD活性密切相关,对孟加拉型的抗性与PPO密切相关;Ptb33对褐飞虱生物型Ⅱ的抗性与PAL和PPO活性密切相关;RP1976-18-6-4-2对褐飞虱生物型Ⅱ的抗性与PAL活性密切相关,对孟加拉型的抗性与PPO活性密切相关;570011对褐飞虱生物型Ⅱ的抗性与PAL和PPO活性密切相关,对孟加拉型的抗性与POD活性密切相关。说明水稻品种对褐飞虱的抗性与PAL、PPO和POD紧密相关,但品种不同,生物型不同,其相关性也有差异。植物在遭受机械损伤、病虫害等生物因子及干旱、盐渗透、高低温等非生物因子胁迫时,会诱导一系列与抗逆性相关的形态学、生物化学、分子生物学变化 (Cheong & Yang, 2003; Frostetal., 2008; Huetal., 2011; Consalesetal., 2012) 来防御或是抗衡这种压力,从而提高植物的抗性。植物的这种抗性,其实是激发各种酶,如蛋白酶抑制剂、苯丙氨酸解氨酶(PAL)、多酚类氧化酶(PPO)、过氧化物酶(POD)和次生代谢产物,如萜类化合物(terpenoids)、酚醛类化合物(phenolics)、丹宁酸(tannins)、生物碱(alkaloids)等参与的防御过程 (Cheong & Yang, 2003; 刘欲强等, 2005; Gulsenetal., 2010; 段灿星等, 2012)。因此,PAL、PPO和POD活性易受环境因素的影响。据报道,烤烟叶片中PAL和PPO活性随施氮量的增加而升高 (王爱华等, 2005);黑暗有利于PAL合成,而光照则是启动和激活PAL所必须 (杨振国和侯凤莲, 1997),不同光照强度对针叶内主要防御蛋白活力有显著影响,50%和25%光照强度下,POD、PAL、PPO等防御蛋白活性显著高于自然光照 (鲁艺芳等, 2012);植物不同生长期防御蛋白活性也不同 (谷岱霏等, 2014)。本研究结果表明,苗龄、光照强度和施氮量对抗虫水稻品种PAL、PPO和POD活性的影响显著,IR56中PAL和PPO活性受苗龄、光照强度和施氮量的影响显著,POD活性受苗龄影响显著;RHT中PAL活性受光照强度和施氮量影响显著,PPO活性受苗龄、光照强度和施氮量的影响显著,POD活性受施氮量的影响显著;Ptb33中PAL活性受光照强度的影响显著,POD活性受苗龄、光照强度和施氮量的影响显著;RP1976-18-6-4-2中PAL活性受光照强度和施氮量影响显著,PPO活性受苗龄、光照强度和施氮量的影响显著,POD活性受苗龄的影响显著;570011中PAL和POD活性受苗龄、光照强度和施氮量的影响显著,PPO活性受苗龄和光照强度的影响显著。苗龄、光照强度和施氮量对抗虫水稻品种PAL、PPO和POD活性的影响是一个复杂的过程,存在显著的交互作用,在不同水稻品种间的变化表现有差异。PAL、PPO和POD是植物形成木质素、酚类等次生代谢物质的关键酶,环境因子对这些防御酶活性有影响,那对次生化合物质的影响如何?有待于进一步研究。
References)
Change JH, Zhang L, Shen SX,etal. Correlation analysis of physical and chemical characteristics with resistance to sorghum aphid melanaphis sacchari in different sorghum genotypes [J].JournalofPlantGeneticResources, 2008, 9(1): 55-61. [常金华, 张丽, 申书兴, 等. 不同基因型高粱的理化特性与抗蚜性的相关性研究[J]. 植物遗传资源学报, 2008, 9(1): 55-61]
Chen JM, Yu XP, Cheng JA,etal. The changes of physiological indexes of different rice varieties after infestation by brown planthopperNilaparvatalugens(Stål) [J].ActaPhytophylacicaSinica, 2003, 30(3): 225-231. [陈建明, 俞晓平, 程家安, 等. 不同水稻品种受褐飞虱危害后体内生理指标的变化[J]. 植物保护学报, 2003, 30(3): 225-231]
Cheong JJ, Yang DD. Methyl jasmonate as vital substance in plants [J].TrendsinGenetics, 2003, 19(7): 409-413.
Consales F, Schweizer F, Erb M,etal. Insect oral secretions suppress wound-induced responses inArabidopsis[J].JournalofExperimentalBotany, 2012, 63(2): 727-737.
Dai L, Li SL, Sun HZ. Preliminary study on the enzyme activities in the barks ofPopulustomentosazoophobous and insect-attacked clones [J].GuizhouForestryScienceandTechnology, 2003, 31(2): 1-5. [代莉, 李淑玲, 孙红召. 毛白杨抗虫、感虫无性系树皮内酶活性的初步研究[J]. 贵州林业科技, 2003, 31(2): 1-5]
Di H, Zhao YY, Chu GX,etal. Silicon-mediated defense enzyme activities in cotton attacked byTetranychusturkestaniand its insect resistance [J].JournalofShiheziUniversity(Natural Science), 2013, 31(6): 661-668. [狄浩, 赵伊英, 褚贵新, 等. 硅对叶螨危害后棉花防御酶活性的调控作用及其与抗虫性的关系[J]. 石河子大学学报(自然科学版), 2013, 31(6): 661-668]
Duan CX, Yu JJ, Li WC,etal. Effect of small brown planthopper stress on defense enzyme activities and cell ultrastructure in rice leaf [J].PlantPhysiologyJournal, 2012, 48(10): 971-978. [段灿星, 余娇娇, 李万昌, 等. 灰飞虱胁迫对水稻叶片中防御酶活性及细胞超微结构的影响[J]. 植物生理学报, 2012, 48(10): 971-978]
Feng DB, Huang JC, Peng ZP,etal. Effects of different organic carbon (nitrogen) nutritions application on main stress-resistant physiological indexes of waterlogged rice [J].JournalofSouthernAgriculture, 2015, 46(12): 2016-2111. [冯定邦, 黄继川, 彭智平, 等. 淹涝胁迫后施用不同有机碳(氮)营养对水稻主要抗逆生理指标的影响[J]. 南方农业学报, 2015, 46(11): 2106-2111]
Frost C, Mescher MC, Carlson JE,etal. Plant defense priming against herbivores: Getting ready for a different battle [J].PlantPhysiology, 2008, 146(3): 818-824.
Gu DF, Hu X, Liu T,etal. Seasonal variations of defense protein activities inTaxuscuspidateleaves [J].JournalofNortheastForestryUniversity, 2014, 42(7): 48-50. [谷岱霏, 胡晓, 刘彤, 等. 不同生长时期东北红豆杉针叶内防御蛋白的变化[J]. 东北林业大学学报, 2014, 42(7): 48-50]
Gulsen O, Eickhoff T, Heng-Moss T,etal. Characterization of peroxidase changes in resistant and susceptible warm-season turfgrasses challenged byBlissusocciduus[J].Arthopod-PlantIneteractions, 2010, 4: 45-55.
Huang FK, Wu BQ, Wei SM,etal. Influence of light intensity and seedling stage on the resistance of rice varieties to rice brown planthopper biotypes [J].JournalofSouthwestAgriculturalUniversity(Natural Science), 2005, 27(2): 143-147, 162. [黄凤宽, 吴碧球, 韦素美, 等. 光照和苗龄对水稻品种抗褐飞虱生物型的影响[J]. 西南农业大学学报, 2005, 27(2): 143-147, 162]
Huang W, Jia ZK, Han QF. Effects of herbivore stress byAphismedicaginisKoch on the malondialdehyde contents and the activities of protective enzymes in different alfalfa varieties [J].ActaEcologicaSinca, 2007, 27(6): 2177-2183.
Hu J, Zhou JB, Peng XX,etal. TheBphi008agene ineracts with the ethylene pathway and transcriptionally regulatesMAPKgenes in the response of rice to brown planthopper feeding [J].PlantPhysiology, 2011, 156 (2): 856-872.
Jia Z, Song ZW, Jin ZY,etal. Studies on the POD activities in pearleaf crabapple (Maluszumi) leaves damstaged by plum spider mite (Tetranychusviennensis) [J].ActaBotanicaBoreal-OccidentaliaSinica, 2004, 24(11): 2136-2139. [贾贞, 宋占午, 金祖荫, 等. 山楂叶螨危害对海棠叶片POD的影响[J]. 西北植物学报, 2004, 24(11): 2136-2139]
Krishnan N, Kodrík D. Antioxidant enzymes inSpodopteralittoralis(Boisduval): Are they enhanced to protect gut tissues during oxidative stress [J].JournalofInsectPhysiology, 2006, 52(1): 11-20.
Li RZ, Mao X, Li CX,etal. The relationship between activities of secondary metabolism-related enzymes and induced resistance to aphids in cotton [J].JournalofShanxiAgriculturalUniversity, 1998, 18(2): 165-168. [李润植, 毛雪, 李彩霞, 等. 棉花诱导抗蚜性与次生代谢相关酶活性的关系[J]. 山西农业大学学报, 1998, 18(2): 165-168]
Liu CM, Wu RZ. Influence of light intensity and nitrogen fertilizer on resistance to the brown planthopper in rice [J].JournalofSouthChinaAgriculturalUniversity, 1992, 13(2): 27-33. [刘春茂, 吴荣宗.光照强度和氮肥对水稻品种抗褐稻虱的影响[J]. 华南农业大学学报, 1992, 13(2): 27-33]
Li WD, Zhou QY. Effects of nitrogen, phosphate fertilizer on phenylalanine ammonia lyase activity in red clover [J].ContemporaryAnimalHusbandry, 2015, 18: 39-42. [李卫东, 周泉佚. 氮肥磷肥对红三叶中苯丙氨酸解氨酶活性的影响[J]. 当代畜牧, 2015, 18: 39-42]
Liu YQ, Jiang L, Sun LH,etal. Changes in some defensive enzyme activity induced by the piercing-sucking of brown planthopper in rice [J].JournalofPlantPhysiologyandMolecularBiology, 2005, 31(6): 643-650. [刘欲强, 江玲, 孙立宏, 等. 褐飞虱刺吸诱导的水稻一些防御酶活性的变化[J]. 植物生理与分子生物学学报, 2005, 31(6): 643-650]
Lu YF, Shi L, Yan SC. Effects of different light intensities on activities of the primary defense proteins in needles ofLarixgmelinii[J].ActaEcologicaSinica, 2012, 32(11): 3621-3627. [鲁艺芳, 石蕾, 严善春. 不同光照强度对兴安落叶松几种主要防御蛋白活力的影响[J]. 生态学报, 2012, 32(11): 3621-3627]
Mauch-Mani B, Slusarenkl AJ. Production of salicylic acid precursors is a major function of phenylalanine ammonia-lyase in the resistance ofArobidopsistoPeronosporaparasitica[J].PlantCell, 1996, 8: 203-212.
Shi R, Chi DF, Zhang SM. Relationship between enzyme activity of ten poplar species and their resistance ability [J].JournalofNortheastForestryUniversity, 2008, 36(9): 74-75. [史瑞, 迟德富, 张晟铭. 10种杨树酶活性与抗性的关系[J]. 东北林业大学学报, 2008, 36(9): 74-75]
Tang LF, Zhong HM, Chen J. Variation ininvivopolyphenol oxidase activity of fourAzollastrains under different illumination conditions [J].ActaPhytophysiologicaSinica, 1999, 25(1): 98-102. [唐龙飞, 钟红梅, 陈坚. 不同光照条件下4种满江红Azolla品系体内多酚氧化酶活性的变化[J]. 植物生理学报, 1999, 25(1): 98-102]
Tscharntke T, Thiessen S, Dolch R,etal. Herbivory, induced resistance and interplant signal transfer inAlnusglutinosa[J].BiochemicalSystematicsandEcology, 2001, 29(10): 1025-1047.
Wang AH, Wang SF, Gong CR. The dynamic effects of nitrogen rate on polyphenols of flue-cured tobacco upper leaf [J].JournalofNorthwestSci-TechUniversityofAgricultureandForestry(Natural Science Edition), 2005, 33(3): 58-60. [王爱华, 王松峰, 宫长荣. 氮素用量对烤烟上部叶片多酚类物质动态的影响[J]. 西北农林科技大学学报(自然科学版), 2005, 33(3): 58-60]
Wang ML, Hu ZL, Zhou MQ,etal. Advances in research of polyphenol oxidase in plants [J].ChineseBulletinofbotany, 2005, 22(2): 215-222. [王曼玲, 胡中立, 周明全, 等. 植物多酚氧化酶的研究进展[J]. 植物学通报, 2005, 22(2): 215-222]
Wu BQ, Huang FK, Huang SS,etal. Inflence of seedling ststage and nitrogen fertilizer on resistance of rice varieties to different rice brown planthopper biotypes [J].SouthwestChinaJournalofAgriculturalSciences, 2009, 22(2): 315-318. [吴碧球, 黄凤宽, 黄所生, 等. 苗龄和施氮量对水稻品种抗褐飞虱不同生物型的影响[J]. 西南农业学报, 2009, 22(2): 315-318]
Wu BQ, Huang FK, Wei SM,etal. Influence of light intensity and nitrogen fertilizer on resistance of rice varieties to different rice brown planthopper biotypes [J].SouthwestChinaJournalofAgriculturalSciences, 2004, 17(6): 733-737. [吴碧球, 黄凤宽, 韦素美, 等. 光照强度和施氮量对水稻品种抗褐飞虱不同生物型的影响[J]. 西南农业学报, 2004, 17(6): 733-737]
Yan JX, Guan HN, Chi DF,etal. Induction of defensive enzymes in leaves ofRosarugosa‘plena’ by mechanical wounding [J].BulletinofBotanicalResearch, 2013, 33(4): 499-503. [严俊鑫, 观桦楠, 迟德富, 等. 机械损伤对重瓣玫瑰防御酶的诱导[J]. 植物研究, 2013, 33(4): 499-503]
Yan SC, Men LN, Shi L,etal. Comparative analysis of antioxidase activities in needles of different larch hybrids [J].ScientiaSilvaeSinicae, 2011, 47(10): 111-115. [严善春, 门丽娜, 石蕾, 等. 杂种落叶松不同子代针叶内抗氧化酶活性比较[J]. 林业科学, 2011, 47(10): 111-115]
Yang ZG, Hou FL. On effect illumination upon maize phenylalanine ammonia lyase activity[J].JournalofJilinForestryUniversity, 1997, 13(3): 151-153. [杨振国, 侯凤莲. 光照对玉米苯丙氨酸解氨酶活性的影响[J]. 吉林林学院学报, 1997, 13(3): 151-153]
Zeng YS, Wang Z. Role of phenylalanine ammonia-lyase in plant disease resistance [J].JournalofZhongkaiAgrotechnicalCollege, 1999, 12(3): 56-65. [曾永三, 王振. 苯丙氨酸解氨酶在植物抗病反应中的作用[J]. 仲恺农业技术学院学报, 1999, 12(3): 56-65]
Zhang JF, Xue QZ. The activity dynamics of main protective enzymes in rice plants under feeding stresses ofSogatellafurciferaandNilaparvatalugens[J].ScientiaAgriculturaSinica, 2004, 37(10): 1487-1491. [张金锋, 薛庆中. 稻飞虱为害胁迫对水稻植株内主要保护酶活性的影响[J]. 中国农业科学, 2004, 37(10): 1487-1491]
Zhang J, Sun XJ, Shi Y,etal. Response of defensive enzyme activity and some primary metabolites inLonicerajaponicaThunb. leaves to aphid damstage [J].ShandongAgriculturalSciences, 2014, 46(8): 57-60. [张金, 孙秀娟, 石岩, 等. 蚜虫为害对忍冬叶片防御酶活性及初生代谢的影响[J]. 山东农业科学, 2014, 46(8): 57-60]
Zhang K, Xu B, Meng ZJ,etal. Effect of copper and cadmium on defensive protein activity in poplar leavea [J].JournalofNortheastForestryUniversity, 2014, 42(11): 43-46. [张凯, 徐波, 孟昭军, 等. 铜、镉胁迫对杨树叶片中防御蛋白活性的影响[J]. 东北林业大学学报, 2014, 42(11): 43-46]
Zhuang BC, Yue DR, Wang YM,etal. The correlation of contents of flavonoid, total phenolate and enzymes in soybean with resistance level toL.Glycinivorella[J].OilCropsofChina, 1992, 3: 18-20. [庄炳昌, 岳德荣, 王玉民, 等. 大豆不同品种次生代谢产物及相关酶类含量与抗食心虫的关系[J]. 中国油料, 1992, 3: 18-20]
Influence of seedling stage, light intensity and nitrogen (N) application rate on the main defensive enzymes activity in rice varieties resistant to brown planthopper (BPH),Nilaparvatalugens(Stål)
WU Bi-Qiu1, LI Cheng1, SUN Zu-Xiong2, HANG Suo-Sheng1, ZHOU Jun-Lei3, HUANG Feng-Kuan1,*, LING Yan1, JIANG Xian-Bin4, HUANG Qian4, LONG Li-Ping4
(1. Plant Protection Research Institute, Guangxi Academy of Agricultural Sciences/Guangxi Key laboratory for Biology of Crop Diseases and Insect Pests, Nanning 530007, China; 2. Plant Protection Station of Fangchenggang City, Fangchengguang 538001, Guangxi Province, China; 3. Agricultural College, Guangxi University, Nanning 530005, China; 4. Rice Research institute, Guangxi Academy of Agricultural Sciences, Nanning 530007, China)
In order to clarify the effect of seedling stage, light intensity and N application rate on the activities of defensive enzymes of rice and relationship with the resistance to brown planthopper (Nilaparvatalugens), the activities of phenylalnine ammonialyase (PAL), polyphenol oxidase (PPO) and peroxidase (POD) in different rice varieties were studied after treated with different seedling stage, light intensity and nitrogen (N) application levels. The results showed that the resistance of IR56, Ptb33 and 570011 to BPH biotype II was closely related with the activities of PAL and PPO, RHT to BPH biotype II was related with the activities of PAL, PPO and POD, RP1976-18-6-4-2 to BPH biotype II was related with the activities of PAL, and 570011 to BPH biotype II was related with the activities of PAL and PPO. The resistance of IR56 to BPH biotype Bangladesh was closely related with the activities of PAL, PPO and POD, resistance of RHT and RP1976-18-6-4-2 to BPH biotype Bangladesh was closely related with the activity of PPO and resistance of 570011 to BPH biotype Bangladesh was closely related with the activities of POD. The separate action or interaction effect of seedling stage, light intensity and N application rate on activities of defensive enzymes in rice was obviously, and effect degree different in different varieties. The separate action or interaction influence of seedling stage, light intensity and N application rate on PAL and PPO activity of IR56, PPO activity of RHT and RP1976-18-6-4-2, POD activity of Ptb33 and PAL and POD activity of 570011 was significantly. POD activity of IR56 and RP1976-18-6-4-2 only significantly affected by seedling stage. The separate action or interaction effect of light intensity and N application rate on PAL activity of RHT and RP1976-18-6-4-2 was obviously. POD activity of RHT affected by N application rate and PAL activity of Ptb33 affected by light intensity was significantly. PPO activity of 570011 affected by seedling stage and light intensity separate action or interaction was obviously.
Seedling stage; light intensity; N application rate; insect-resistant rice; defensive enzyme; brown planthopper
国家自然科学基金项目(31160369,31560510);“十二五”国家科技支撑计划项目(2012BAD19B03);广西科学研究与技术开发计划项目(桂科合14125007-2-4);广西自然科学基金项目(2015GXNSFAA139054);广西作物病虫害生物学重点实验室基金项目(14-045-50-ST-06);广西农业科学院科技发展基金项目(2015JZ46)
吴碧球,女,1978年生,广西贺州人,博士,副研究员,研究方向为水稻害虫综合防控、水稻抗虫性及机制,E-mail: wubiq@126.com
*通讯作者 Author for correspondence, E-mail: huangfengkuan@gxaas.net.
2016-11-01;接受日期:2016-11-14
S433.1;S435.112+.3
A
1674-0858(2016)06-1121-13
