青海湖畔3种高原盐生植物叶绿体淀粉粒的变化特征
2016-12-22周勇辉刘玉萍张晓宇
周勇辉,刘玉萍,张晓宇,吕 婷,胥 芮,苏 旭
(青海师范大学 生命与地理科学学院 青藏高原药用动植物资源重点实验室 青海省自然地理与环境过程重点实验室 青藏高原环境与资源教育部重点实验室,西宁 810008)
青海湖畔3种高原盐生植物叶绿体淀粉粒的变化特征
周勇辉,刘玉萍,张晓宇,吕 婷,胥 芮,苏 旭
(青海师范大学 生命与地理科学学院 青藏高原药用动植物资源重点实验室 青海省自然地理与环境过程重点实验室 青藏高原环境与资源教育部重点实验室,西宁 810008)
以同种非盐生植物作为对照,利用透射电子显微镜对盐胁迫下3种高原盐生植物——鹅绒委陵菜、西伯利亚蓼和灰绿藜叶绿体淀粉粒的变化特征进行了研究。结果表明,在非盐胁迫下3种高原盐生植物其超微结构无明显差异;而在低温及盐胁迫等多种逆境生态条件下,3种高原盐生植物叶绿体中淀粉粒呈现大量积累且增大的趋势,同时它们多呈圆形沿叶绿体长轴,电子密度中间高、四周低的不均匀分布。此外,还发现3种高原盐生植物产生的潴泡与其叶绿体类囊体程度不同的膨大密切相关。据此,我们认为3种高原盐生植物中淀粉粒增大、增多及潴泡的产生,应是青海湖畔盐渍、低温、低气压、强辐射等多种逆境生态条件胁迫下的反应。
青海湖;盐生植物;亚显微结构;淀粉粒
高原盐生植物(Alpine halophyte)即高海拔盐生植物,是指生长在高原树线以上至雪线[1],属天然植物体区系,盐渍土壤渗透压不低于3.35 × 105Pa,即70 mmol·L-1单价盐[2]。在中国,高原盐生植物主要分布于青藏高原盐渍地区。绝大多数高等植物体系中,淀粉是植物光合作用下主要的光合产物,大多以颗粒状态形式存在,也是碳水化合物在细胞质中最普遍的储藏形式。通常,淀粉是植物光合作用下产生的,由葡萄糖分子聚合而成的长链化合物,以便暂时贮藏在植物茎叶中。淀粉粒水解成葡萄糖,转运至细胞质中储藏并重新聚合成淀粉粒。淀粉粒在形态及结构上因不同的植物种类,与所处的环境不同而产生较大的区别。
迄今,国内外对青藏高原地区高山植物的研究,主要集中于形态、显微结构[3-4]、超微结构[5-8]和生理功能[9-12]等学科领域,但对高山盐生植物的研究报道较少[13-18],尤其是对叶绿体中淀粉粒的研究仍属空白。本文以青海湖畔自然生长的3种高原盐生植物——西伯利亚蓼(Polygonumsibiricum)、鹅绒委陵菜(Potentillaanserina)和灰绿藜(Chenopodiumglaucum)为实验材料,对其淀粉粒在叶绿体中的变化特征进行较为系统地探讨,同时结合生态环境与淀粉粒变化关系进行分析,旨在揭示高原盐生植物与生态环境之间的结构性关系,为高原盐生植物在植物生理学方面的研究提供理论依据。
1 材料与方法
1.1 研究区概况
青海湖地处北纬35°32′~37°15′,东经99°36′~100°47′,是我国面积最大的内陆湖泊,属于新构造断陷而形成的咸水湖,东西长约106 km,南北宽约63 km,平均海拔3196 m,湖水容量743亿m3,最大水深27 m,平均水深16 m;矿化度12.32 g/L,含盐量14.13 g/L,透明度在3m以下。由于入湖水量入不敷出,导致湖面水位下降幅度增大,湖中含氧量极低,浮游生物十分稀少;强辐射、低温、盐渍、低气压等为青海湖畔生态环境条件的显著特征。年均辐射为6344.30 kJ·m-2,年降水量为289.5~579.5 mm,年蒸发量大,年日照时数2699~3089.9 h,1月份平均气温-24.0℃~-6.7℃,7月份平均气温6.7℃~18.7℃,极端最低最高气温分别是-35.8℃、25.6℃。
1.2 研究方法
位于青海湖畔东北部距水陆交界5 m处,于7月—8月份植物生长旺季,土壤盐浓度为1000 ds·cm-1;Alt.3212 m的区域,采取全受光生长健壮的植物叶片中部为实验材料,利用单面刀片切割,同时避免大叶脉将其切割成1 mm2的小块,并迅速投入0.2 mol/L(pH 7.2)磷酸缓冲液配制的4 %戊二醛溶液中固定12 h,然后用其冲洗4次,再用0.2 mol·L-1(pH 7.2)磷酸缓冲液配制的2 %饿酸溶液在4℃环境下固定4 h,利用不同溶度乙醇进行梯度脱水,丙酮溶液过渡,KD-BM生物包埋机包埋,CUT 4062型超薄切片机切片,切片厚度为60 nm,用柠檬酸铅-醋酸双氧铀溶液对切片进行双染色,在日立H-800透射电子显微镜下观察并拍照,同一切片上进行5个不同视野的照相保存,取视野中25个叶绿体的测量数据作为平均数。
2 观察结果
2.1 非胁迫下3种高原盐生植物叶肉细胞的超微结构
电子显微镜观察表明,在非盐胁迫条件下3种高原盐生植物的成熟叶片结构基本相同,没有显著差异存在,即它们均具有明显的细胞核、叶绿体以及线粒体等细胞器,并且叶绿体中含有较少的淀粉粒;细胞内细胞器膜结构明显,清晰可见;叶绿体数量较少,呈椭圆形,被膜清晰完整,基粒结构清晰(图1-I~III)。另外,在非盐胁迫下,3种高原盐生植物叶绿体的类囊体均没有出现膨大和破裂的现象(图1-I~III)。
2.2 盐胁迫下3种高原盐生植物叶肉细胞的超微结构
透射电子显微镜观察表明,所研究的3种高原盐生植物的叶绿体中淀粉粒数量均较多(图1-A~F),尤其是在鹅绒委陵菜和西伯利亚蓼的叶绿体中还有巨大淀粉粒存在的现象(图1-C~F);同时,在3种高原盐生植物的叶绿体中,个体较小的淀粉粒通常呈圆形,而个体较大的淀粉粒则往往呈长椭圆形,所有叶绿体内的淀粉粒均沿长轴分布,并且电子密度通常较低(图1-A~F)。此外,所研究的3种高原盐生植物的叶绿体类囊体均出现膨大,薄膜破裂、部分瓦解,甚至空泡化等(图1-A、C、E),特别是在灰绿藜叶绿体中,本研究发现其类囊体破裂最终形成了潴泡(图1-B)。
3 讨论
生态环境是植物生存和发展的条件[19],是植物适应环境与发展的前提,而对环境的适应,在生理功能、结构和形态发生显著变化,逐渐发育了一套适应青海湖畔低温,高盐渍生态环境的生理机制和特殊结构,故揭示植物形态结构与盐生环境的关系,始终是一项重要的课题。青海湖畔具有不利于植物生长的逆境条件,如高盐浓度、低温、强辐射等,尤其是高盐条件下,容易造成土壤在生理上的干旱,因而长期生长于这一区域的植物,在生理功能、结构上都发生改变和产生相应的生理机制。正是这种生理机制的产生,而使植物能在青藏高原强逆境的生态环境下得以正常生长。
3.1 淀粉粒的积累
叶绿体中的淀粉粒是多数高等植物的主要光合产物,其氧化后为植物生长提供大量能量,是细胞碳水化合物主要存在的形式之一,还可为植物叶绿体细胞蛋白质的合成提供碳骨架[20]。研究结果表明[8,21],低海拔盐生植物和高山植物叶绿体中均含有许多较大的淀粉粒,在叶肉细胞中因高山低温和高盐渍的土壤环境造成光合产物运输受阻。所研究的3种高原高盐植物叶肉细胞的亚显微结构时,我们发现它们通常含有2~10个较大的淀粉粒(图1-A~F),这与先前学者对低海拔盐生植物以及高山植物的研究结果都是一致的[8]。通常,高山植物在白天光合作用下进行淀粉积累,夜间进行有氧呼吸而水解,并转运出叶绿体[8],自然高原盐生植物也应符合这种植物生理机制。青海湖地区隶属于高原高寒大陆性气候,晴天多降水少,太阳辐射强烈,大气透明度高,地面对太阳入射和反射能力强,致使青海湖地区昼夜温差大,最高气温7月—8月份,平均气温仍比较低且温差大(6.7℃~18.7℃)。青海湖地区因夜间大气逆辐射弱气温低,植物细胞中的酶活性降低,呼吸代谢能力下降,ATP减少,从而导致植物叶绿体内淀粉粒的水解及光合产物转运受到抑制[8],最终在植物叶绿体中积累了较多较大的淀粉粒。我们推测,3种高原盐生植物叶肉细胞中淀粉粒的大量积累(图1-A)是适应青海湖地区低温和盐胁迫等多种胁迫的主要方式,叶肉细胞内淀粉粒因夜间低温而解体,解体的淀粉粒不仅可以提高细胞质的渗透压,而且还可以提高细胞质的吸水性及细胞质浓度,从而增强了高原盐生植物的抗逆性。另外,糖类氧化分解也是能量释放的一个重要过程,它可以提高细胞的活性,进而提高植物抵抗低温的能力;在高盐渍胁迫下,高山植物叶绿体中淀粉粒的数量通常是非盐胁迫下植物叶绿体内淀粉粒数量的4~5倍。盐生植物叶绿体内较大淀粉粒的积累不但可以缓冲细胞因能量短缺后仍进行正常生命活动,还可以提高细胞对水分的吸收和保持的能力[22],尤其是在高海拔盐胁迫下,叶肉细胞的生理活性和代谢能力均降低较多,同化物的转运系统严重受到抑制,3种高原盐生植物细胞中淀粉粒的大量积累,是抵抗青海湖地区低温、高盐渍等逆境生态环境的重要方式。
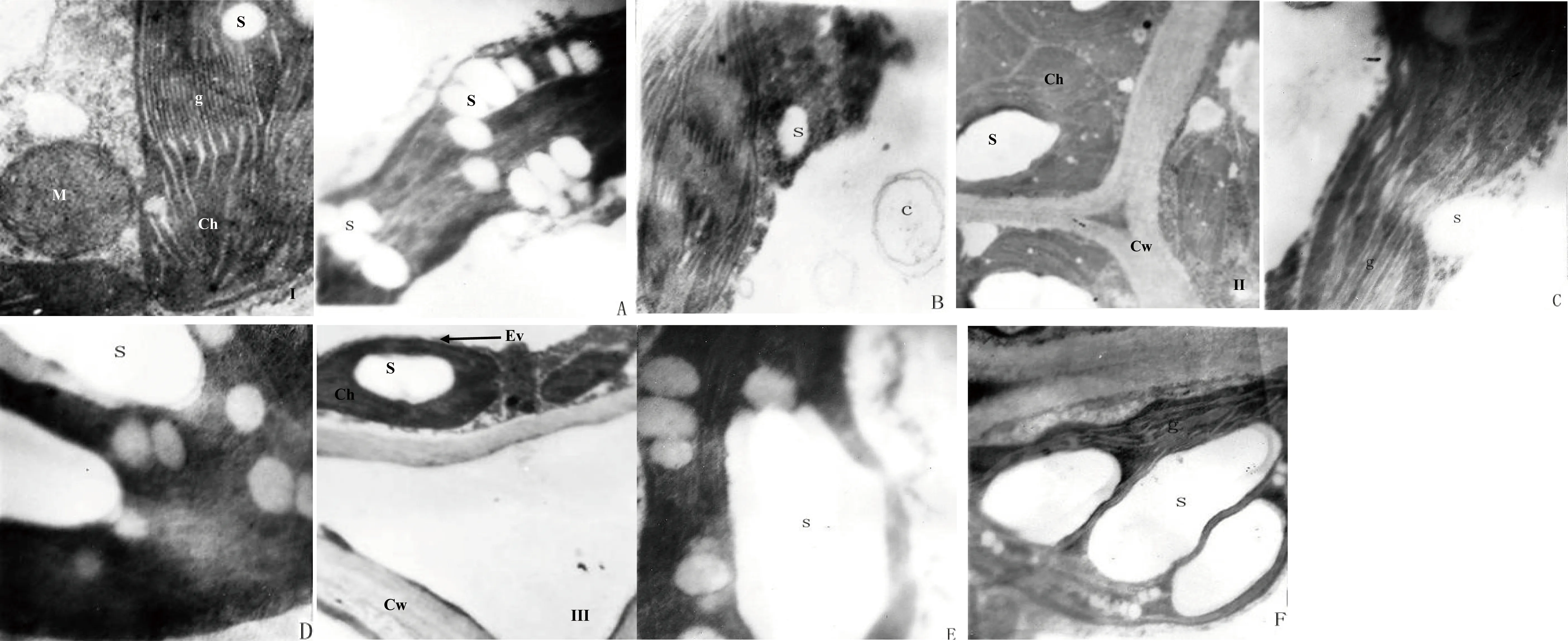
图1 3种高原盐生植物叶绿体淀粉粒的变化特征
Fig 1 Characteristics of starch grains in chloroplast of three alpine halophytes
Ⅰ:正常生长的灰绿藜叶肉细胞的超微结构(×35 660);A:灰绿藜的叶绿体,示膨大的类囊体和淀粉粒(× 4000);B:灰绿藜的叶绿体,示淀粉粒和潴泡(× 5000);Ⅱ:正常生长的鹅绒委陵菜叶肉细胞的超微结构(×13 286);C:鹅绒委陵菜的叶绿体,示膨大的类囊体和淀粉粒(× 5000);D:鹅绒委陵菜的叶绿体,示淀粉粒(× 5000);Ⅲ:正常生长的西伯利亚蓼叶肉细胞的超微结构(×13 478);E:西伯利亚蓼的叶绿体,示膨大的类囊体和淀粉粒(× 5000);F:西伯利亚蓼的叶绿体,示淀粉粒(× 3000)。g:基粒;s:淀粉粒;c:潴泡;Ch:叶绿体;Ev:叶绿体被膜;Cw:细胞壁
3.2 潴泡的产生

3.3 淀粉粒积累与光合速率的关系
先前学者在研究植物光合产物过程中,发现光合速率与光合产物的积累或转运存在必然的联系。已有研究[28-29]表明,低温逆境生态因子的胁迫,可使植物体内淀粉粒在形态、结构或数量等方面发生变化,如淀粉粒形状变大,数量增多,同时植物体内生理代谢失调,光合速率下降。目前,就光合速率与光合产物的积累是否存有关联?目前有两种截然不同的观点:一种观点[30]认为光合速率下降导致叶绿体内大颗粒淀粉的积累是受低温影响,同时光合速率与光合产物之间会出现负反馈抑制作用;另一种观点[31]则认为,在一定区域内,叶片中产物储存的多寡和光合能力的大小将决定植物叶片光合产物的转运能力。当植物叶片转运光合产物的数量和速度提高时,说明叶片的光合速率得到提高,产物形成与积累不断增加;反之,当植物叶片对光合产物的转运能力下降时,表明植物叶片光合作用能力下降。光合速率与光合作用之间不存在反馈抑制作用,至此植物叶片光合转运能力的大小随着光合作用能力的变化而发生改变。大量研究[8-9,11-12]表明,高山植物的光合速率通常较低。潘瑞炽和董愚得[32]认为,夜晚黑暗环境或光合速率下降,磷酸丙糖在细胞质中产生的量减少,而Pi含量则相对较高,结果导致磷酸丙糖/Pi的比值降低,低的磷酸丙糖/Pi比将会刺激果糖-6-磷酸-2-激酶活性,进而形成较多的果糖-2,6-二磷酸酶,结果果糖-1,6-二磷酸酶的水解遭到抑制,蔗糖的合成速度受到抑制,磷酸丙糖在植物叶绿体向淀粉合成方向流动,最终在植物叶绿体内合成2~10个淀粉粒或更多数量的淀粉。何涛等[8]通过几种高山植物叶绿体淀粉粒变化特征的研究,认为高山植物淀粉粒水解能力下降,淀粉粒在植物叶绿体中大量积累,这是由于高山地区昼夜温差大夜间温度低所导致的,因此高山植物光合速率的下降与淀粉粒在叶绿体内积累密切相关。然而,关于高原盐生植物淀粉粒与光合速率关系的研究目前尚属空白。本研究表明,青海湖畔3种高原盐生植物的叶绿体中,数目较多且体积较大的淀粉粒,这与先前关于高山植物叶绿体中淀粉粒的变化原因是一致的。青海湖畔盐生植物不但受到盐胁迫的影响,将受到低温、低气压、强辐射等多种逆境生态因子的胁迫,这些逆境生态因子可能共同作用于高原盐生植物,从而导致它们对淀粉水解和光合产物的运输能力急剧下降,从而导致其叶绿体内大量的淀粉粒得以积累。据此,我们认为高原盐生植物淀粉粒的积累很可能就是光合速率下降的结果,这与高山植物叶绿体中淀粉粒与光合速率的关系类似。
[1]王勋陵,王 静.植物形态结构与环境[M].兰州:兰州大学出版社,1989.
[2]GREENWAY H,MUNNS R.Mechanisms of salt tolerance in nonhalophytes[J].Annals Review of Plant Physiology,1980,31: 149-190.
[3]周广泰,刘风琴,郭书贤,等.青海高山植物解剖特点的研究[J].青海师范大学学报(自然科学版),1992(4): 45-60.
[4]吴学明.五种高山藏医药用植物茎叶的结构特征研究[J].西北植物学报,1996,16(1): 56-60.
[5]祁如虎,李有忠,范建平.16种高山植物叶片内的异细胞及其生态学意义[J].西北植物学报,1998,18(2): 270-276.
[6]吴学明.高山植物盘花垂头菊和唐古特乌头光合膜系统的超微结构研究[J].西北植物学报,1997,17(5): 98-102.
[7]杜军华,吴学明,刁治民,等.达坂山蚤缀和裸茎金腰叶绿体超微结构的研究[J].西北植物学报,2001,21(5): 1031-1034.
[8]何 涛,吴学明,张文静,等.高山植物叶绿体与线粒体位置相关性研究[J].西北植物学报,2004,24(4): 728-731.
[9]何 涛,吴学明,张改娜,等.几种高山植物叶绿体淀粉粒的变化特征[J].武汉植物学研究,2005,23(6): 545-548.
[10]卢存福,贲桂英,韩 发,等.矮嵩草光合作用与环境因素关系的比较研究[J].植物生态学报,l995,19(1): 72-78.
[11]师生波,贲桂英,赵新全,等.增强UV-B辐射对高山植物麻花艽净光合速率的影响[J].植物生态学报,2001,25(5): 520-524.
[12]魏 捷,贲桂英,余 辉,等.青海高原不同海拔地带生长的珠芽蓼光合特性的比较[J].生物物理学报,1998,14(3): 530-536.
[13]卢存福,简令成,贲桂英.高山植物短管兔儿草光合作用特性及其对冰冻胁迫的反应[J].植物学通报,2000,17(6): 559-564.
[14]黄志伟,彭 敏,陈桂琛,等.青海湖盐碱湿地灰绿藜叶的形态解剖学研究[J].西北植物学报,2001,21(6): 1199-1203.
[15]刘吉祥,吴学明,何 涛,等.盐胁迫下芦苇叶肉细胞超微结构的研究[J].西北植物学报,2004,24(6): 1035-1040.
[16]苏 旭,吴学明,祁生贵,等.青海湖畔两种盐生植物叶片的超微结构研究[J].青海草业,2004,13(3): 14-16.
[17]刘玉萍,吴学明,苏 旭.青海湖畔3种典型盐生植物叶片超微结构研究[J].青海草业,2005,14(4): 16-23.
[18]刘玉萍,苏 旭,李建民,等.青海湖畔3种典型盐生植物叶片超微结构研究[J].华中师范大学学报(自然科学版),2014,48(1): 91-96.
[19]苏 旭,刘玉萍,左晓丽.青海湖畔3种典型盐生植物叶片结构研究[J].草地学报,2014,22(1): 194-198.
[20]卢存福,简令成,匡廷云.高山植物的抗寒抗冻特性[J].植物学通报,1998,15(3): 17-22.
[21]SHINOZAKI K,YAMAGUCHI-SHINOZAKI K.Gene expression and signal transduction in water-stress response[J].Journal of Plant Physiology,1997,115(2): 327-334.
[22]胡永红,张启翔,王奎玲,等.低温对切花菊叶片细胞器超微结构的影响[J].莱阳农学院学报,2000,17(1): 38-43.
[23]BROWN R H,RIGSBY L L,AKIN D E.Enclosure of mitochondria by chloroplasts[J].Plant Physiology,1983,71(2): 437-439.
[24]SALIN M L.Toxic oxygen species and protective systems of the chloroplast[J].Physiologia Plantarum,1988,72(3): 681-689.
[25]蔡志全,曹坤芳,冯玉龙,等.夜间低温对两种热带树苗活性氧代谢的影响[J].西北植物学报,2002,22(4): 859-864.
[26]SAILEROVA E,ZWIAZEK J J.Effects of triadimefon and osmotic stress on plasma membrane composition and ATPase activity in white spruce (Piceaglauca) needles[J].Physiology Plantarum,1993,87: 475-472.
[27]WANG H C.Plant stress physiology[J].Plant Physiology Common,1981,6: 72-81.
[28]陈健辉,李荣华,郭培国,等.干旱胁迫对不同耐旱性大麦品种 叶片超微结构的影响[J].植物学报,2011,46(1): 28-36.
[29]SATO K,PARK K B.The physiological changes of high plant at chilling temperature[J].Japanese Journal of Crop Science,1982,5: 402-406.
[30]PARK I K,TSUNODA S.Effect of low temperature on chloroplast structure in cultivar of rice[J].Plant and Cell Physiology,1979,20(7): 1449-1453.
[31]CHATERTON N J,CARLSON G E,HUNGERFORD W E,et al.Effect of tillering and cool nights on photosynthesis and chloroplast starch in pangola[J].Crop Science,1972,12(2): 206-208.
[32]潘瑞炽,董愚得.植物生理学[M].3版.北京:高等教育出版社,2000: 97-98.
Characteristics of starch grains in chloroplast ofthree alpine halophytes near the Qinghai Lake
ZHOU Yong-hui,LIU Yu-ping,ZHANG Xiao-yu,LYU Ting,XU Rui,SU Xu
(Key Laboratory of Medicinal Plant and Animal Resources the Qinghai-Tibetan Plateau,Key Laboratory ofPhysical Geography and Environmental Process in Qinghai Province,Key Laboratory of Education Ministry of Environments and Resources in the Qinghai-Tibetan Plateau,College of Geography and Life Science,Qinghai Normal University,Xining 810008,China)
The ultrastructure of chloroplast in the mesophyll cells of three typical alpine halophytes includingPotentillaanserina,PolygonumsibiricumandChenopodiumglaucumgrown at low temperature and in salt marsh near the Qinghai Lake was studied by using the transmission electron microscope.The result showed that the subcellular structure did not differ between the three halophytes under non-stress condition,but there were a lot of starch grains in the chloroplast of three alpine halophytes,which were usually round or oval in shape arranged along the longitudinal axis of chloroplasts,and the whole starch grains were low in electron-density.Meanwhile,the thylakoid of chloroplasts swelled differently,and some of them even became into cisterna.It suggested that the variations of starch grains in the chloroplast of three alpine halophytes stated above were the result of the long-time stress under the adverse ecological factors such as low temperature,low air pressure,strong radiation,salt marsh,etc.,and were an adaptation to the environment.
Qinghai Lake; halophyte; ultrastructure; starch grain
2016-01-12;
2016-04-08
收稿日期:国家自然科学基金项目(31260052);青海省自然科学基金项目(2014-ZJ-947Q);青藏高原药用动植物资源重点实验室(2015-Z-Y06);青海师范大学2015年度本科生科技创新项目(2015-12,2015-13)资助
周勇辉,硕士研究生,主要从事植物形态解剖学研究
苏旭,博士后,教授,硕士生导师,主要从事植物形态解剖学研究,E-mail: xusu8527972@126.com
10.3969/j.issn.2095-1736.2016.06.073
Q942.4
A
2095-1736(2016)06-0073-05
