松墨天牛包囊蛋白基因的克隆及表达特性分析
2016-12-20毛珊珊吴华俊
毛珊珊,吴华俊,林 同
(华南农业大学 林学院,广东 广州 510642)
松墨天牛包囊蛋白基因的克隆及表达特性分析
毛珊珊,吴华俊,林 同
(华南农业大学 林学院,广东 广州 510642)
松墨天牛Monochamus alternatus是危害马尾松树重大蛀干害虫,也是松材线虫的主要传播媒介昆虫,对我国松林资源构成严重威胁。用cDNA末端快速扩增(RACE)方法克隆了松墨天牛ERP基因(MalERP),GenBank登录号为KF357881.1。该基因含有一个长为864 bp的开放阅读框,编码287个氨基。同源性分析表明,MalERP与光肩星天牛Anoplophora glabripennis ERP的同源性最高,达到79%;与赤拟谷盗Tribolium castaneum的同源性为45%。实时荧光定量PCR分析显示,MaERP在松墨天牛3龄幼虫的脂肪体、马氏管、中肠、体壁、血淋巴和头部中均有表达,在脂肪体中的表达量最高;MaERP经溴氰菊酯、磷化铝、Bt、印楝素、绿僵菌胁迫后,上调表达;但经阿维菌素胁迫后,下调表达。本研究为进一步研究MalERP基因在松墨天牛防御过程中的作用奠定基础,为探索松墨天牛免疫系统与分子毒理互作关系提供参考。
松墨天牛;包囊蛋白;基因表达;农药胁迫
松墨天牛Monochamus alternates属鞘翅目天牛科,是一种危害马尾松树的蛀干害虫,又是松材线虫病的主要媒介昆虫,对我国松林资源己构成严重威胁[1]。
昆虫的天生免疫可以简单的分为体液免疫和细胞免疫[2],当入侵物较多时,细胞通过吞噬作用清除入侵物,当外源物太大而不能被血细胞吞噬时,包括寄生蜂卵和幼蜂,真菌和原生动物寄生虫,甚至非生物对象[3-4],昆虫血细胞常会在外源物外形成一层或多层的鞘状物质以包囊外源物来杀死病原物[5]。一般认为:当外源物进人寄主血腔后,寄主血细胞与外源物随机接触,若颗粒血细胞接触到外源物,在原酚氧化酶级联产物、凝集素等的协助下,它能够识别外源物为“异己”进而附于外源物上,并释放出识别因子以吸引浆血细胞。浆血细胞附着后,在颗粒血细胞与外源物的复合体上延展,并且浆血细胞间常会形成粒桥、微管,这样层层埋集,形成包囊[6-7]。包囊是无脊椎动物对抗外源物质的主要防御反应[8]。在不同种的昆虫中,参与包囊反应的细胞种类也有所差异。在鳞翅目中,参与包囊作用的主要是粒细胞和浆细胞,而在果蝇Drosophila melanogaster中是叶状细胞[9]。
包囊蛋白(encapsulation-relating protein,ERP)是昆虫产生包囊反应时最先在外来物周围积聚的蛋白[8]。目前国内外对ERP的研究较少,最早测得的鞘翅目ERP基因来自黄粉虫Tenebrio molitor。本研究用cDNA末端快速扩增(Rapid ampli fi cation of cDNA ends,RACE)方法克隆了松墨天牛ERP基因,用实时荧光定量PCR (Realtime quantitative PCR,RT-qPCR) 分析该基因在不同组织中的表达谱,为进一步研究该基因在松墨天牛防御反应中的作用奠定基础。通过探讨不同农药处理后ERP基因表达情况,为探索松墨天牛免疫系统与分子毒理互作关系提供参考。
1 材料与方法
1.1 松墨天牛
松墨天牛幼虫采自广州市萝岗区马尾松次生林,配置人工饲料[10],置于智能人工气候箱内饲养,饲养条件为黑暗、温度(25士1.0)℃、相对湿度70%~75%。
1.2 组织分离
选取松墨天牛3龄幼虫,用酒精消毒虫体并对其麻醉。用解剖针将幼虫固定在解剖板上,背部向上,用剪刀剖开,移液器收集血淋巴;解剖镜下分别分离脂肪体、马氏管、中肠、头部和体壁,置于离心管中立即用液氮处理,-80 ℃保存备用。
1.3 主要试剂
pMD20-T Vector试 剂 盒、E.coliDH5α Competent Cells、Taq DNA聚合酶、dNTP 购自宝生物公司(TaKaRa);UNIQ-10柱式Trizol总RNA抽提试剂盒、DNA cloning Vector 试剂盒、T4 DNA连 接 酶、EZ Spin column total RNA Isolation Kit、3'Full RACE Core Set Ver.2.0 kit、LA Taq DNA聚合酶、dNTP、MgCl2、IPTG、X-Gal购自上海生工生物技术有限公司; PrimeScript® RT reagent Kit With gDNA Eraser、 pMD20-T Vector试 剂 盒、E.coliDH5α Competent Cells、SYBR® Premix Ex TaqTM购自TaKaRa公司;Universal通用型DNA纯化回收试剂盒、DNA Marker (D2000)、质粒小提试剂盒、氨苄青霉素钠盐等购自天根生化科技有限公司。
1.4 松墨天牛总RNA的提取
从-80℃冰箱中取出松墨天牛幼虫样品,参照UNIQ-10柱式Trizol总RNA抽提试剂盒说明书提取松墨天牛总RNA。纯化后的总RNA经琼脂糖凝胶电泳检测其质量,并用微量紫外分光光度仪 (Nanodrop 2000) 检测其纯度并定量后,置于-80℃保存备用。
1.5 RACE方法扩增ERP基因
本研究在以往获得的松墨天牛ERP基因5'端序列(GenBank:JZ143844)[11]的基础上进行3'RACE扩增,以获得松墨天牛ERP基因全序列。用DNAStar中的Primer select软件设计EPR基因特异性引物(ERPouter,ERPinner,见表1),结合试剂盒提供的两个下游引物 3'RACE outer Primer 和3'RACE inner Primer 进行巢式 PCR[12]。首先用通用引物3'RACE Outer Primer和特异性引物ERPouter进行第一次 Outer PCR扩增, PCR反应条件为:94℃预变性 3 min,94℃ 30 s,55℃ 30 s,72℃1 min,20个循环,然后72℃延伸 10 min。反应完毕后取1 µL反应产物,使用通用引物3'RACE Inner PCR 和特异性引物ERPinner进行巢式PCR的Inner PCR扩增。反应进行30个循环,其它反应条件同第一次 Outer PCR扩增。PCR产物经由2%的琼脂糖凝胶电泳检测,用Universal通用型DNA纯化回收试剂盒回收目的DNA片段,与pMD20-T载体连接,将连接产物转化DH5α感受态细胞,进行蓝白斑筛选,挑阳性菌落提取质粒。经PCR鉴定后送上海生工生物技术有限公司测序。
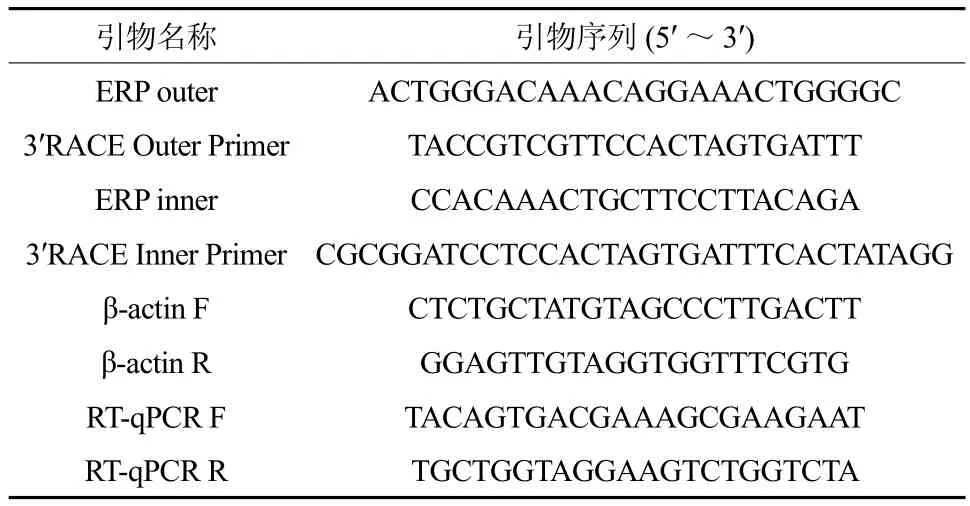
表1 本研究所使用的引物Table 1 Primers used in the study
1.6 序列分析
使用DNAMAN软件将测得的ERP基因序列进行拼接和开放阅读框(ORF)预测,所得序列在NCBI上进行同源性比较分析 (http://www.Ncbi.Nlm.gov/blast/); 使 用 BioEdit、ProtScale等 软 件对推测的ERP进行氨基酸组成、疏水性分析;用TMpred (http://ch.embnet.org/software/TMPRED_form.html) 进行跨膜区分析,预测跨膜蛋白的跨膜片段;COILS Server(http://www.ch.embnet.org/software/COILS_form.html) 预测卷曲螺旋;NetPhos(http://www.cbs.dtu.dk/services/NetPhos/) 预 测 磷酸化修饰位点;丹麦科技大学(DTU) 的CBS服务器上的SignalP 4.0 Server程序进行蛋白质序列的信号肽 (signal peptide) 预测 (http://www.cbs.dtu.dk/services/SignalP/);用NCBI (http: //www. Ncbi. nlm.Nih. gov/) 中的blastx程序进行同源性比对[13]。
1.7 农药处理
用6种农药对松墨天牛3龄整只幼虫喷雾或熏蒸,每种农药处理设3个重复,每个重复10头幼虫。磷化铝熏蒸持续进行1.5 d,其他农药喷雾处理后,将试虫静置6 h;之后将每种农药处理的试虫分别提取RNA,等量混合,作为RT-qPCR样品。使用的药剂名称和使用方法等详见表2。
1.8 RT-qPCR
获得ERP的ORF后,以此设计引物(RT-qPCRF, RT- qPCR,见表1)。RNA提取方法同1.4,按照 PrimeScript® RT reagent Kit With gDNA Eraser说明合成cDNA第1链。以β-actin (见表1)为内参基因,以无菌超纯水为阴性对照。每个反应设置3个平行实验[14]。两步法PCR反应程序为:95℃ 5 min ,1个循环(预变性);95 ℃ 10 s,60℃ 20 s,40个循环(PCR反应);40℃ 30 s (冷却)。反应结束后采集目标基因的 Ct 值和两个内参基因的 Ct 平均值,利用 2-ΔΔCT相对定量法计算ERP的相对表达量。 RT-qPCR在LightCycler Real Time PCR中进行。
2 结果与分析
2.1 松墨天牛ERP基因的克隆与序列分析
用DNAMAN软件将测序所得的序列和原序列拼接结果如图1所示,该序列全长为1 571 bp,3′端包含12 bp的PolyA尾巴和一个特征性序列AATAAA。经过DNAMAN预测,该序列ORF为864 bp,编码287个氨基酸,推测分子量为31.772 kDa, 将松墨天牛ERP基因命名为MaERP,GenBank登录号为KF357881.1。
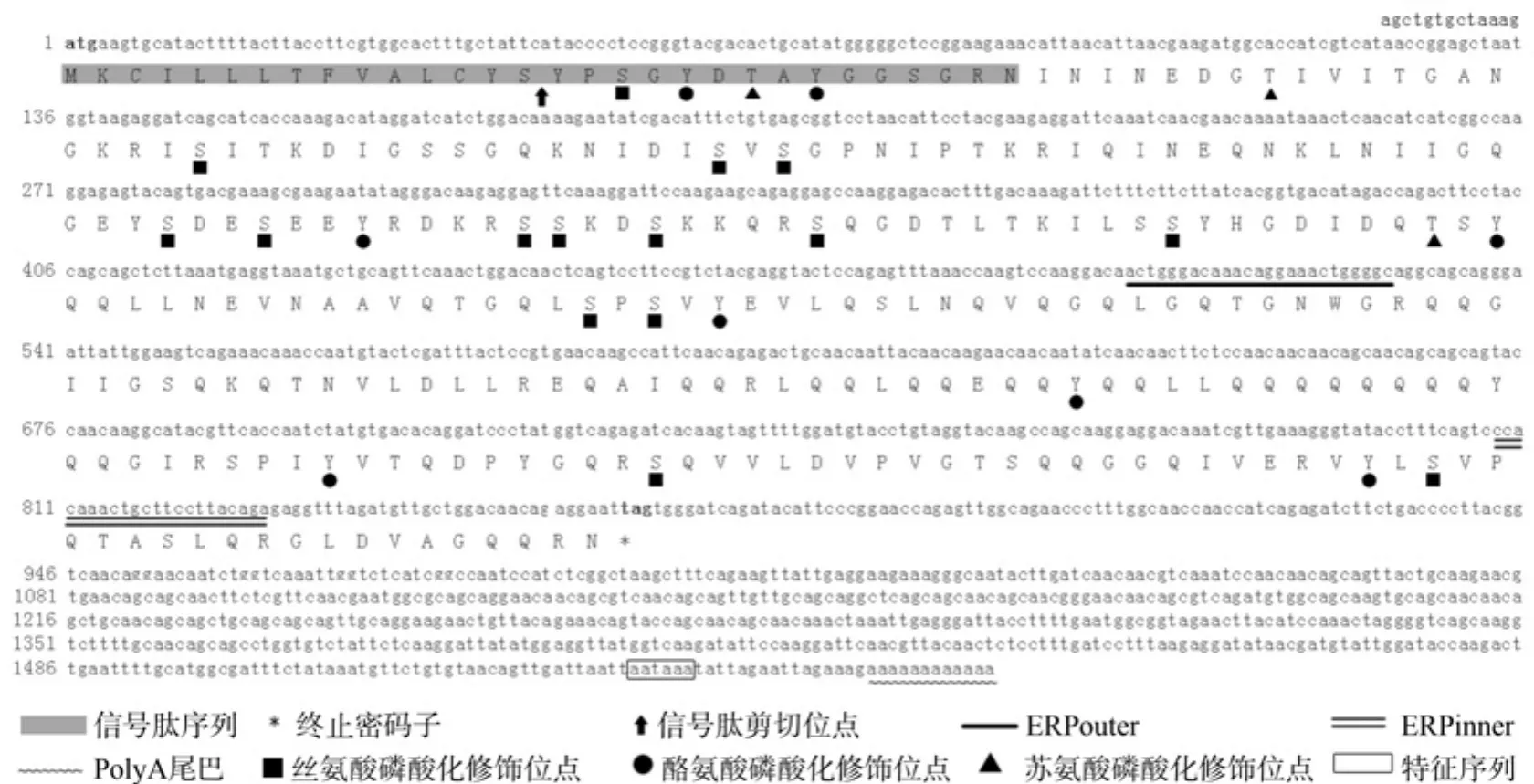
图1 MaERP基因全长及其推导的氨基酸序列Fig.1 Nucleotide and deduced amino acid sequences of MaERP
由MaERP推测的蛋白Gln (谷氨酰胺)含量最多,摩尔百分比接近18%; Gly(甘氨酸)含量次之,摩尔百分比达10%。此结果与黄粉虫幼虫56 kDaERP的氨基酸组成特点一致,黄粉虫的ERP组成特点是富含Gln(谷氨酰胺),Gly含量也居第二位。在8~13氨基酸位含有一个跨膜片段,方向为膜内向膜外,且为疏水性区域;含有6个卷曲螺旋,有丝氨酸磷酸化位点15个,酪氨酸的磷酸化位点8个,苏氨酸的磷酸化位点3个。蛋白质磷酸化是将外界刺激转换为胞内信号的一个主要机制,是生物界最普遍也是最重要的一种蛋白质翻译后修饰(Post-Translational Modi fi cations,PTMs)。由此可知此类蛋白参与免疫过程的胞内信号传导。信号肽序列为MKCILLLTFVALCYSYPSGYDTAYGGSGRN,信号肽的剪切位点位于第15和16位氨基酸处。
2.2 同源性比对
经由blastx做出的MaERP与其它4种昆虫ERP的氨基酸序列同源性比对的结果(见图2)可知,MaERP与光肩星天牛Anoplophora glabripennis ERP同源性最高,高达79%;与赤拟谷盗Tribolium castaneum和黄粉虫的同源性分别为45%和43%;与玉米根虫Diabrotica virgifera的同源性最低,为26%。
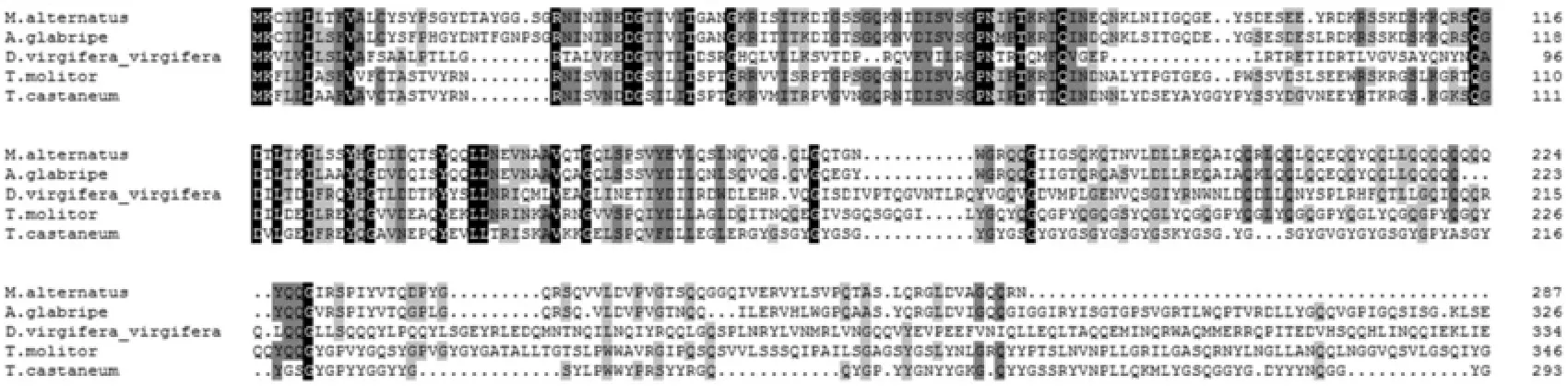
图2 MaERP与其它4种昆虫 ERP氨基酸序列比对Fig.2 Amino acid sequence alignment of MaERPfrom M. alternatusand ERPs from other 4 insects
2.3 MaERP表达分析
如图3所示,MaERP在脂肪体中的相对表达量最高,是体壁的13.2倍;在马氏管、中肠和血淋巴中的表达量分别为在体壁中的3.1倍、1.7倍和1.4倍;在头部的相对表达量最低,仅为在体壁中的10%。
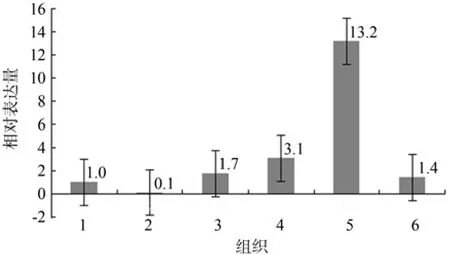
图3 松墨天牛幼虫不同组织中MaERP表达谱Fig.3 Expression pro fi les of MaERP in different tissues of 98 larvae detected by RT-qPCR
由图4可知,经阿维菌素胁迫后,MaERP的相对表达量最低,只是对照的0.05倍,为下调表达,说明阿维菌素明显抑制了MaERP的表达;溴氰菊酯、磷化铝、Bt.、印楝素、绿僵菌胁迫后,MaERP的表达量分别是对照的134.5倍、111.1倍、82.3倍、56.5倍和31.4倍,为上调表达,表明这5种农药对MaERP的表达起增强作用。
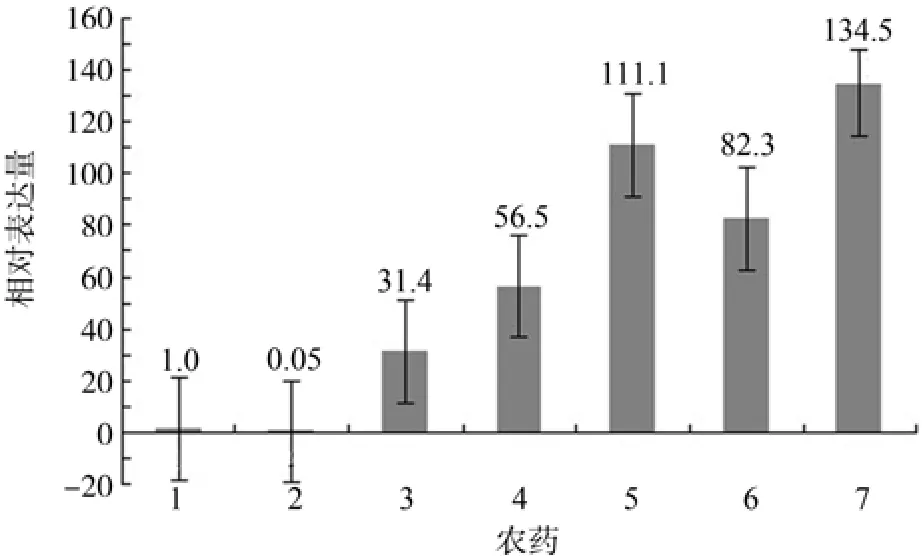
图4 农药胁迫对MaERP表达量的影响Fig.4 Expression levels of MaERPafter pesticide stress
3 结论与讨论
本研究用RACE方法成功克隆了MaERP基因,该基因的ORF为864 bp,编码287个氨基酸;MaERP与光肩星天牛ERP氨基酸同源性为79%;MaERP在松墨天牛3龄幼虫的脂肪体、马氏管、中肠、体壁、血淋巴和头部中均有表达,但在头部的表达量最低;MaERP在溴氰菊酯、磷化铝、Bt、印楝素、绿僵菌等5种农药胁迫下上调表达,在阿维菌素胁迫下下调表达。
通过对MaERP基因编码蛋白结构的分析,推测MaERP可能是一种产生于细胞膜内而分泌于细胞膜外、直接对细胞壁起作用的蛋白。由于细胞各部分都有特定的蛋白质组分,因此,合成的蛋白质必须准确无误地定向运送才能保证生命活动的进行。一般认为,蛋白质定位的信息存在于该蛋白质自身结构中,并且通过与膜上特殊受体的互相作用得以表达。在起始密码子后,有一段编码疏水性氨基酸序列RNA片段,这个氨基酸序列就称为信号序列。本研究预测的MaERP信号肽位置与长度和Cho等人通过分子克隆和化学分析法得到的黄粉虫56 kDa ERP所预测到的信号肽的位置和长度相同,都处于N末端;而且剪切位点的位置也都在15位和16位氨基酸之间[8]。此结果表明得到了完整的MaERP开放阅读框。下一步可以表达出MaERP,对其进行亚细胞定位,以期深入研究MaERP的功能。
昆虫的脂肪体是其体内的一种重要结构,它既是糖、脂和蛋白质代谢的中心,又是主要激素(如神经激素、蜕皮激素、保幼激素)作用的靶组织,还具有贮存排泄和解毒等生理功能。Cho等通过蛋白印迹法发现,黄粉虫幼虫中的86 kDa ERP在血淋巴和脂肪体中的含量明显较多,表明ERP在脂肪体中合成并分泌到血淋巴中[8]。根据本研究中RT-qPCR分析的结果,在松墨天牛3龄幼虫各组织中,脂肪体中MaERP相对表达量最大,为在体壁中表达量的13.2倍,与Cho等的结果[8]一致。推测ERP在昆虫的脂肪体中的含量是最高的。
昆虫的表皮层作为首要物理及化学屏障能够有效阻止病原物的附着与入侵。昆虫消化道作为病原物入侵的另一个主要途径,能够有效阻滞病原物入侵,抑制病原物活性,消解病原物残体。昆虫最初的防御是以体壁作为物理屏障,其次的防御是中肠及血淋巴中的凝集反应,还有细胞毒素分子在受伤部位形成的多种产物[7]。MaERP在马氏管、中肠和血淋巴中的表达量为在体壁中的1~3倍,表明ERP在体壁、马氏管、中肠和血淋巴中均可能参与细胞免疫包囊反应。相对于体壁,MaERP在松墨天牛头部的表达量最少,仅为在体壁中的0.1倍,预示在头部的包囊反应较为薄弱。
阿维菌素对虫体的作用机制是使神经膜处于抑制状态,阻断神经冲动传导而使昆虫丧生[15]。经阿维菌素处理后,MaERP的表达量仅为对照的0.05倍,说明阿维菌素对MaERP的表达有极强的抑制作用。推测阿维菌素也影响到了松墨天牛的免疫系统。
在溴氰菊酯等其它5种农药作用下,MaERP上调表达,表明这些农药可能刺激了松墨天牛产生包囊反应,因而致使ERP的表达量增高。昆虫具备相当完善的先天免疫系统,它可通过特定的蛋白分子快速有效地介导整个免疫过程。昆虫居住环境的类型较为复杂,为了应对外部的选择压力,昆虫已进化出不同类型且高效的防御策略,以更好地保护自己,抵御外来物,包括农药的入侵。研究因分子介导所产生的对包囊反应的抑制或诱发,有助于了解无脊椎动物细胞免疫反应的基本过程[16]。本研究中使用的农药分属于植物性农药、微生物农药、拟除虫菊酯类农药、内酯类抗生素等,杀虫机理不同,可为探索不同类型农药的分子毒理和包括包囊作用在内的昆虫防御反应之间的互作提供参考。
[1]周华康. 2%噻虫啉微胶囊粉剂对松墨天牛的防治效果[J]. 林业科技,2012,37(4): 21-22.
[2]STRAND M R. The insect cellular immune response[J]. Insect Science,2008,15(1): 1-14.
[3]CARTON Y,NAPPI A J. Drosophila cellular immunity against parasitoids[J]. Parasitol Today,1997,13(6): 218 - 227.
[4]Marmaras V J,Lampropoulou M. Regulators and signalling in insect haemocyte immunity[J]. Cellular Signalling,2009,21(2):186-195.
[5]Schmidt O,Theopold U,Strand M. Innate immunity and its evasion and suppression by hymenopteran endoparasitoids[J].BioEssays,2001,23 (4): 344-351.
[6]Lavine M D,Strand M R. Surface characteristics of foreign targets that elicit an encapsulation response by the moth Pseudoplusia includes[J]. Journal of Insect Physiology,2001,47(9): 965-974.
[7]Lavine M D,Strand M R. Insect hemocytes and their role in immunity[J]. Insect Biochemistry and Molecular Biology,2002,32(10): 1295-1309.
[8]Cho M Y,Lee H S,Lee K M,et al. Molecular cloning and functional properties of two early-stage encapsulation-relating proteins from the coleopteran insect,Tenebrio molitor larvae[J].European Journal of Biochemistry,1999,262(3): 737-744.
[9]Vass E,Nappi A J. Fruit Fly Immunity[J]. BioScience,2001,51(7): 529-535.
[10]徐金华,黄秀凤,徐华潮,等.松墨天牛室内人工饲养及其生物学特性观察[J]. 浙江林业科技,2009,29(4): 86-88.
[11]Wei Chunmei,Luo Linlin,Wu Huajun,et al. Oligonucleotide icroarray-based gene expression analysis of pine sawyer(Monochamus alternatus) after treatment with a sublethal dose of di fl ubenzuron[J]. Journal of Asia-Paci fi c Entomology,2013,16:489-495.
[12]王建勇,谭晓风,龙洪旭,等. 油茶脂氢过氧化物裂解酶基因的克隆与序列分析[J].中南林业科技大学学报,2014,34(6):49-54.
[13]王 淋,乌云塔娜,刘慧敏,等.杜仲MVA 途径相关基因全长cDNA 序列特征研究[J]. 中南林业科技大学学报,2014,34(1): 94-101.
[14]刘 凯,梁国校,杨 麒,等. 油桐DGAT1 基因在不同品种及种子发育阶段的表达模式[J].中南林业科技大学学报,2014,34(10): 61-64.
[15]王广成,张忠明,高立明,等.阿维菌素的作用机理及其应用现状[J]. 植物医生,2009,19(1): 4-5.
[16]Choi J Y,Whitten M M A,Cho M Y,et al. Calreticulin enriched as an early-stage encapsulation protein in wax moth Galleria mellonella larvae[J]. Developmental and Comparative Immunology,2002,26 (4): 335-343.
Cloning and expression of encapsulation-relating protein gene of Monochamus alternatus
MAO Shan-shan,WU Hua-jun,LIN Tong
(College of Forestry,South China Agricultural University,Guangzhou 510642,Guangdong,China)
Pine sawyer,Monochamus alternatus Hope (Coleoptera: Cerambycidae),a serious trunk borer in pines stands,also is a major media insects of pine wood nematode,posed a serious threat to the forest resources in China. It is the key vector of the exotic pine wood nematode Bursaphelenchus xylophilus throughout Asia. The ERP gene from M. alternates,called MaERP (GenBank accession number is KF357881.1),was cloned and sequenced by rapid ampli fi cation of cDNA ends. Sequence analysis revealed that the open reading frame of MaERP had 864 bp in full length encoding a 287 predicted amino acids residues. Blastx showed that MaERP and Anoplophora glabripennis had EPR homology,up to 79%,and had EPR homology to Tribolium castaneum,being 45%. The Real-time PCR quantitative analysis showed that MaERP was expressed in body wall,head,midgut,Malpighian tubule and hemolymph from the 3rdinstar larvae of M. alternatus,and expressed in fat body with the highest expression. The RT-qPCR also indicated that MaERP was upregulated when exposed to Metarrhizium anisopliae,azadirachtin,aluminum phosphide,Bt and deltamethrin,and downregulated when exposed to avermectin.The findings of the study lay a foundation for further research on MalERP’s defensive reaction to M.alternatus,provide a reference for studying the interaction between immune system and molecular toxicology of M. alternates.
Monochamus alternatus; encapsulation-relating protein; gene expression; pesticide stress
S718.7
A
1673-923X(2016)01-0107-05
10.14067/j.cnki.1673-923x.2016.01.018
2014-05-10
国家自然科学基金项目(31170612)
毛珊珊,硕士研究生 通讯作者:林 同,副教授,博士;E-mail:lintong@scau.edu.cn
毛珊珊,吴华俊,林 同. 松墨天牛包囊蛋白基因的克隆及表达特性分析[J].中南林业科技大学学报,2016,36(1): 107-111,133.
[本文编校:谢荣秀]
