复合菌共生发酵法酶解甘薯蛋白制备生物活性肽的研究
2016-12-19李彦军胡云红
王 勇,李彦军,,王 楠,胡云红
(1.陕西农产品加工技术研究院,陕西西安 710021;2.陕西科技大学食品与生物工程学院,陕西西安 710021;3.陕西省科学院酶工程研究所,陕西临潼 710600)
复合菌共生发酵法酶解甘薯蛋白制备生物活性肽的研究
王 勇1,李彦军1,2,王 楠2,胡云红3
(1.陕西农产品加工技术研究院,陕西西安 710021;2.陕西科技大学食品与生物工程学院,陕西西安 710021;3.陕西省科学院酶工程研究所,陕西临潼 710600)
以甘薯蛋白为原料,采用复合菌共生发酵法制备生物活性肽。通过SephadexG-25型色谱柱对发酵产物进行分离,并利用高效液相色谱法做进一步的分析验证,测定了产物的DPPH自由基清除能力及还原能力。结果表明,甘薯蛋白在枯草芽孢杆菌与黑曲霉比例1.5∶1.0,以10%的接种量在5%甘薯蛋白发酵培养基中发酵48 h后,可水解为具有一定DPPH自由基清除能力以及还原能力的小分子肽类物质。
复合菌;甘薯蛋白;生物活性肽
0 引言
甘薯(Ipomoea batatas)是旋花科甘薯属的一个重要栽培品种,原产于南美洲,因具有高产、稳产、抗干旱、耐瘠薄、适应性强、营养丰富等特点,在我国广泛种植[1]。甘薯块根除含有大量的淀粉外,还有2.24%~12.21%(以干物质计)的甘薯粗蛋白。目前,对甘薯蛋白的研究主要集中在贮藏蛋白和糖蛋白[2]。由于未变型的甘薯蛋白具有胰蛋白酶抑制活性,影响其消化吸收,如使之生成小分子量的肽,会使其生物效价更高[3]。同时,研究表明甘薯蛋白多肽具有降血压、抗凝血、提高免疫力、抗肿瘤等功能,并对二苯基苦味肼基自由基(DPPH·)具有一定的清除能力[4]。
微生物发酵法是指利用微生物产生的蛋白酶来水解蛋白质,从而制备生物活性肽的一种方法[5],具有微生物来源广泛、所产蛋白酶不用分离、生长周期短、生产成本相对酶解法较低等优点。枯草芽孢杆菌所产蛋白酶主要是内切酶,能从蛋白质内部进行水解切断肽链,生成小分子的多肽和氨基酸;黑曲霉所产的酸性蛋白酶主要是端肽酶,具有很好的热稳定性和金属离子稳定性,耐酸性好[6]。从理论上讲,将枯草芽孢杆菌和黑曲霉按比例混合,用混合菌种发酵甘薯蛋白粉,所产生的酶中既有内肽酶又有端肽酶,可以同时在肽链的中间和两端进行切割,从而制备更多的甘薯多肽。目前,有关以黑曲霉和枯草芽孢杆菌为复合菌共生发酵酶解甘薯蛋白制备生物活性肽的研究还尚未见报道。试验以甘薯蛋白为底物,运用微生物共生发酵技术制备甘薯蛋白活性肽,旨在为甘薯蛋白及活性肽开发利用提供一定的研究基础。
1 材料与方法
1.1 材料与试剂
甘薯蛋白粉,实验室自制,甘薯蛋白含量85%;枯草芽孢杆菌、黑曲霉,由陕西省科学院酶工程技术研究所提供;营养琼脂培养基等试剂原料,均为分析纯。
1.2 仪器与设备
UV-1800型紫外分光光度计,北京瑞利有限公司产品;高效液相色谱仪,美国Waters产品;LSC50L型立式压力蒸汽灭菌器,江阴滨江医疗设备厂产品;HS-100型恒温恒湿试验箱,苏州晨光仪器公司产品。
1.3 复合菌共生发酵酶解甘薯蛋白
将实验室保藏的枯草芽孢杆菌和黑曲霉分别接种到液体种子培养基中,进行扩大培养,于温度30℃,转速160 r/min条件下,振荡培养36 h后用于发酵。菌种活化完成后,将菌种接种到甘薯蛋白发酵培养基中,根据枯草芽孢杆菌∶黑曲霉为2∶1 ~1∶2的比例,在接种量为8%~14%,培养箱温度为30℃条件下进行恒温培养,并每隔一段时间观察发酵液的形态,于发酵进行6,12,24,48,60,72 h后,分别采用双缩脲反应法检测发酵液中蛋白质酶解的程度。
发酵结束后,将发酵液在100℃下灭酶10 min,然后以转速4 000 r/min离心15 min,去除沉淀菌体和未发酵的甘薯蛋白残渣,取上清液,并加入适量饱和硫酸铵溶液,静置,滤液用0.45 μm微孔滤膜过滤,得甘薯蛋白发酵液,待用。
1.4 葡聚糖凝胶色谱柱分离甘薯多肽
以葡聚糖凝胶G-25为分离介质、蒸馏水为洗脱剂,于流速1.35 mL/min条件下,对甘薯蛋白发酵液进行分离纯化,分段收集洗脱液,每管5 mL,依次编号为G1~Gx,并于波长214,220,280 nm处进行紫外检测定性。
1.5 高效液相色谱法分离甘薯多肽
选择甘薯多肽富集度较高的洗脱液,利用高效液相色谱法进行进一步分析鉴定。色谱条件为色谱柱C18,检测波长214 nm,柱温30℃,上样量20 μL,流速1.0 mL/min,并按照表1中的条件梯度洗脱50 min(流动相A:0.1%三氟乙酸+100%乙腈;流动相B:0.1%三氟乙酸+100%去离子水)。
高效液相色谱法梯度洗脱条件见表1。
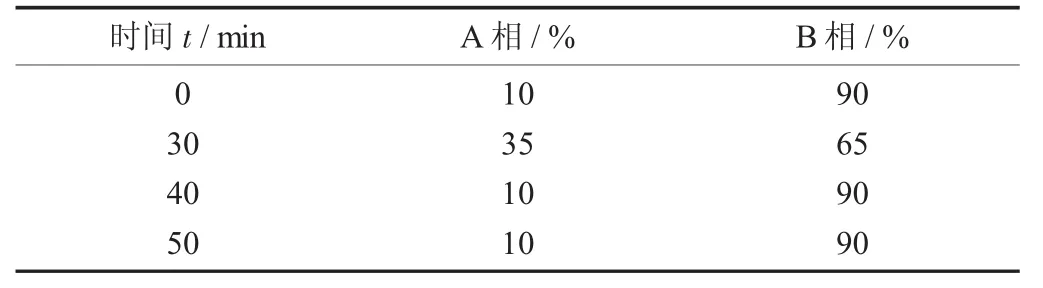
表1 高效液相色谱法梯度洗脱条件
1.6 甘薯多肽DPPH自由基清除能力及还原能力的测定
1.6.1 DPPH自由基的清除能力测定
参照Orhan I等人[7]的方法测定甘薯多肽清除DPPH自由基能力。
取试样2 mL与0.2 mmol/L的DPPH自由基无水乙醇溶液2 mL混合均匀,置于黑暗处反应30 min后取出,于波长517 nm处测定紫外吸光度;以2 mL蒸馏水与2 mL DPPH自由基无水乙醇溶液的混合物作为空白对照,用无水乙醇调零。甘薯多肽对DPPH自由基的清除率通过以下公式计算:

式中:X——甘薯多肽对DPPH自由基的清除率;
A0——2 mL蒸馏水与2 mL DPPH自由基无水乙醇溶液混合后的吸光度;
A1——2 mL甘薯多肽溶液与2 mL DPPH自由无水乙醇溶液反应后的吸光度。
1.6.2 还原能力的测定
参照Kaur R等人[8]的方法测定甘薯多肽还原能力。
将2 mL试样与5 mL磷酸盐缓冲液(0.2 mol/L,pH值6.6)混合均匀,然后加入5 mL的1%铁氰化钾溶液,混合物在50℃条件下保温20 min;再加入5 mL的10%TCA溶液,混和均匀,以转速3 000 r/min离心10 min;取2 mL上清液和1 mL蒸馏水混合,再加入0.5 mL的0.1%FeCl3溶液,最后于波长700 nm处测量吸光度。吸光度越高,说明试样的还原能力越强。
2 结果与分析
2.1.1 不同枯草芽孢杆菌和黑曲霉接种比例对发酵结果的影响
分别将枯草芽孢杆菌和黑曲霉按照2.0∶1.0,1.5∶1.0,1.0∶1.0,1.0∶1.5,1.0∶2.0的比例,以10%的接种量接种在5%甘薯蛋白发酵培养基中,于30℃条件下进行恒温培养,48 h后采用双缩脲反应法对发酵结果进行检测,绘制发酵液吸光度与复合菌接种比例的关系曲线。
不同枯草芽孢杆菌和黑曲霉接种比例对发酵结果的影响见图1。

图1 不同枯草芽孢杆菌和黑曲霉接种比例对发酵结果的影响
双缩脲反应是肽和蛋白质所特有而氨基酸没有的一个颜色反应。由于蛋白质分子中含有许多与双缩脲结构相似的肽键,能与试剂中的铜离子在碱性溶液中发生双缩脲反应,且颜色深浅与蛋白质含量的关系在一定范围内符合比尔定律,而与蛋白质的氨基酸组成及分子量无关,因此可用双缩脲法测定蛋白质的含量。即甘薯蛋白被酶解的越充分,紫外吸光度就越低。由图1可知,当枯草芽孢杆菌和黑曲霉的混合比例为1.5∶1.0时,发酵效果最佳。
2.1.2 不同接种量对发酵结果的影响
在枯草芽孢杆菌和黑曲霉复合菌的混合比例为1.5∶1.0时,分别选用8%,9%,10%,11%,12%,13%,14%的接种量将复合菌接种在5%甘薯蛋白发酵培养基中,在培养箱温度为30℃条件下进行培养,48 h后采用双缩脲反应法对发酵结果进行检测。
不同复合菌接种量对发酵结果的影响见图2。
初稿完成后,对照脚本用相关仪器再实际操作几次,记下每一步之后会出现什么样的界面和提示,再修改脚本,然后定稿。

图2 不同复合菌接种量对发酵结果的影响
发酵开始前,甘薯蛋白培养基中溶解了大量的蛋白质,表现出较高的紫外吸光度;随着发酵进行,大分子的蛋白质被分解成了小分子的多肽及氨基酸,紫外吸光度下降。当复合菌接种量较少时,由于菌种浓度过低,致使甘薯蛋白粉发酵不完全,上清液中存在比较多的大分子蛋白质,从而在540 nm处表现出较高的紫外吸光度;随着复合菌接种量的增加,甘薯蛋白被酶解的越多。由图2可知,当复合菌接种量增大到10%时,甘薯蛋白被酶解的最多;此后随着接种量的增大,紫外吸光度的变化并不显著。因此,复合菌的最佳接种量为10%。
2.1.3 不同发酵时间对发酵结果的影响
在枯草芽孢杆菌和黑曲霉的混合比例为1.5∶1.0,接种量为10%的条件下,于30℃进行恒温培养,考察发酵时间对发酵结果的影响。
不同发酵时间对发酵结果的影响见图3。
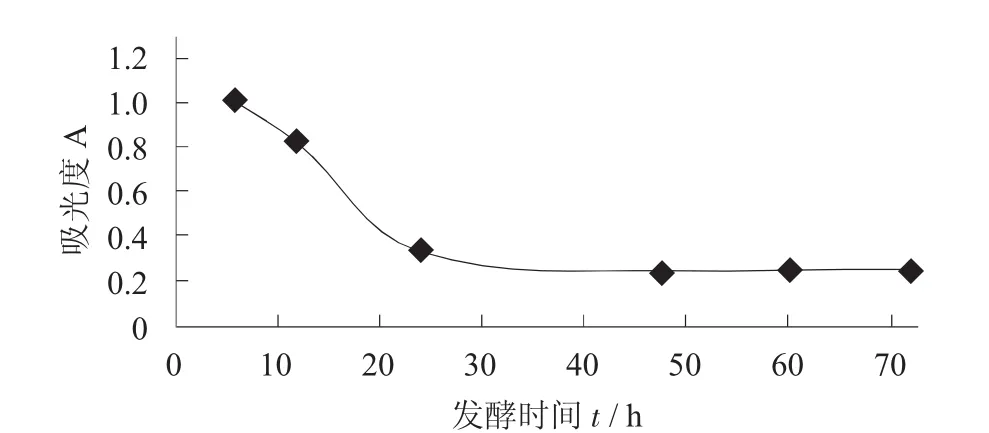
图3 不同发酵时间对发酵结果的影响
由图3可知,随着发酵时间的延长,发酵上清液的紫外吸光度逐渐减小。其原因为甘薯多肽的浓度与发酵液中蛋白酶的活性有直接关系,发酵液中蛋白酶的活性又随着微生物代谢的加快而逐渐增加,48 h之后随着微生物的逐渐衰亡产酶量又逐渐降低。当复合菌发酵48 h左右时,甘薯蛋白的酶解程度最高,之后甘薯蛋白发酵产物的紫外吸光度基本不变,这说明随着发酵的进行,甘薯蛋白的酶解程度先高后低,至48 h左右基本不变。因此,选择48 h作为发酵的最佳时间。
2.2 葡聚糖凝胶色谱柱分离甘薯多肽
在上述最优条件下复合菌共生发酵法酶解甘薯蛋白,发酵结束后按1.3的方法灭酶,经离心、除杂后取上清液,饱和硫酸铵溶液盐析静置,滤液用0.45 μm微孔滤膜过滤,得甘薯蛋白发酵液。
取样品2.5 mL在1.4中的试验条件下进行分离,分段收集洗脱液,并分别于波长214,220,280 nm处进行紫外检测,绘制洗脱曲线。
葡聚糖凝胶色谱柱分离滤过液洗脱曲线见图4。

图4 葡聚糖凝胶色谱柱分离滤过液洗脱曲线
据蛋白质与肽类物质的紫外吸收特性分析,蛋白质的紫外特征吸收峰为280 nm,多肽的紫外特征吸收峰多为214,220 nm。试验选择上述3个特征波段为检测点,绘制甘薯蛋白水解液洗脱曲线,综合分析蛋白的水解情况以及多肽溶液在葡聚糖凝胶色谱柱中的洗脱情况。由图4可知,洗脱液在280 nm处吸光度随着洗脱液的流出(管号的增加)变化不大且吸光度较小,这说明洗脱液中不含甘薯蛋白或甘薯蛋白较少,表明甘薯蛋白已基本被水解;随着洗脱液的流出,从第5管开始,洗脱液在214 nm及220 nm处的紫外吸光度开始明显增加,并于第8、第9管达到峰值,表明甘薯蛋白已在复合菌的作用下水解为肽类物质,此后随着管号的增加吸光度逐渐减少。结合葡聚糖凝胶色谱柱的分离原理可知,随着洗脱液的流出,未水解的残余蛋白及分子量大的多肽首先流出,紧接着是分子量相对较小的短肽、二肽以及氨基酸等小分子物质。
2.3 高效液相色谱法分离甘薯多肽结果分析
将2.2中的5~15管洗脱液收集编号,标记为G5~G10,并进行紫外扫描及薄层色谱分析后,选择甘薯多肽富集度较高的G8,G9以及制备的5.52 mg/mL丝氨酸标准品,5.58 mg/mL甘氨酸标准品按照1.5中所述的方法进行高效液相色谱分析。
丝氨酸标准品液相见图5,甘氨酸标准品液相见图6,G8洗脱液液相见图7,G9洗脱液液相见图8。

图5 丝氨酸标准品液相

图6 甘氨酸标准品液相

图7 G8洗脱液液相
由图7和图8可知,G8,G9的高效液相色谱图相似度较高,但有一定的区别,峰形较杂乱,分离度不高,可能原因为所制备的样品组分复杂,且分子量相近,这也从侧面反映了微生物发酵的不确定性;另一方面也可能为在试验条件下采用Sephadex G-25型色谱柱以及高效液相色谱法对试验所制备的肽类物质分离效果不佳,今后可从此处入手做进一步优化。据报道,生物活性肽的分子量范围一般为50~1 000 Da,结合图5和图6丝氨酸以及甘氨酸的液相色谱图,可以推断G8,G9分子量较小及其分布较广,分子量在100~1 000 Da范围内,这与活性肽的分子量范围一致。经综合分析可知,在甘薯蛋白的微生物发酵液中存在小分子肽类物质,且分子量较小、范围广。

图8 G9洗脱液液相
2.4 甘薯多肽DPPH自由基清除能力、还原能力的测定结果
2.4.1 甘薯多肽对DPPH自由基的清除能力结果分析
根据2.2的试验结果,选取G5~G12号洗脱液,参照1.6中的方法测试洗脱液对DPPH自由基的清除能力。
甘薯多肽对DPPH自由基的清除能力见图9。

图9 甘薯多肽对DPPH自由基的清除能力
由图9可知,从G5开始,随着洗脱液的流出,甘薯蛋白水解物对DPPH自由基的清除能力逐渐增强,其中在G9时出现了极大值,DPPH自由基的清除能力达到了68.19%;此后随着洗脱液的流出,DPPH自由基的清除能力逐渐减弱。这说明甘薯蛋白水解物具有一定的DPPH自由基清除能力,这与文献报道一致,进一步验证了甘薯蛋白已在复合菌的作用下水解为活性较高的小分子肽结论,且高活性肽主要集中于G8和G9中。
2.4.2 甘薯多肽还原能力的测定结果
将2.2中的5~12号洗脱液参照1.6中的方法进行还原能力的测定。
甘薯多肽的还原能力测定见图10。

图10 甘薯多肽的还原能力测定
由图10可知,随着洗脱液管号的增大,甘薯蛋白水解物的还原能力先增强后减弱。G9具有最强的还原能力,这与甘薯活性肽具有较强还原能力的特征一致。结合2.1,2.2,2.3试验结果,进一步说明甘薯蛋白已在复合菌的作用下生成了肽类物质,且产物的还原能力较好。
3 结论
以甘薯蛋白为原料,采用复合菌共生发酵法制备甘薯活性肽。枯草芽孢杆菌和黑曲霉在甘薯蛋白发酵培养基中可以较好地生长,产生的蛋白酶对甘薯蛋白进行酶解,得到了小分子多肽和氨基酸。通过单因素试验,得出复合菌共生发酵的最佳条件为枯草芽孢杆菌和黑曲霉按照1.5∶1.0的比例以10%的接种量接种在5%甘薯蛋白发酵培养基中,发酵48 h可使甘薯蛋白得到充分酶解。利用葡聚糖凝胶色谱柱的同时依据分子筛原理分离甘薯蛋白发酵液,并通过高效液相色谱法进行进一步分析检测,水解液基本得到了分离,收集了富集度较高的甘薯活性肽样品,但分离效果不佳,有待进一步研究改进。经检测,样品对DPPH自由基具有一定的清除能力以及还原能力。
[1]马代夫.世界甘薯生产的发展与预测 [J].世界农业,2001(1):17-19.
[2]Maeshima M,Sasaki T,Asahi T.Characterization of major proteins in sweet potato tuberous roots[J].Phytochemistry,1985,24:1 899-1 902.
[3]吴广辉,木泰华,高愿军,等.Alcalase碱性蛋白酶水解甘薯蛋白的研究 [J].食品科技,2008(7):22-25.
[4]石雨,王道园,鲁卫卫,等.甘薯蛋白多肽的制备及其对DPPH自由基清除作用 [J].食品科技,2010,35(5):168-172.
[5]何倩.绿豆多肽的制备工艺及其抗氧化性和促发酵作用 [D].广州:暨南大学,2011.
[6]王莉娟,陶文沂.大豆肽体外抗氧化活性研究 [J].生物加工过程,2008,6(4):69-73.
[7]Orhan I,Kartal M,Abu-Asaker M,et al.Free radical scavenging properties and phenolic characterization of some edible plants[J].Food Chemistry,2009,114(1):276-281.
[8]Kaur R,Arora S,Singh B.Antioxidant activity of the phenol rich fractions of leaves of Chukrasia tabularis A.Juss[J].Bioresource Technology,2008,99:7 692-7 698.
The Study for Complex Symbiotic Fermentation of Sweet Potato Protein to Prepare Bioactive Peptides
WANG Yong1,LI Yanjun1,2,WANG Nan2,HU Yunhong3
(1.Shaanxi Institute of Agricultural Products Processing Technology,Xi'an,Shaanxi 710021,China;2.College of Food and Biological Engineering,Shaanxi University of Science and Technology,Xi'an,Shaanxi 710021,China;3.Institute of Enzyme Engineering,Shaannxi Academy of Sciences,Lintong,Shaanxi 710600,China)
Using sweet potato protein as raw material,the biological activity peptide is prepared by the method of compound bacteria symbiotic fermentation.The fermentation products are separated by SephadexG-25 column,and further analysis is made by using high performance liquid chromatography,the free radical scavenging ability and reducing ability of DPPH·are determined.The results show that Bacillus subtilis and Aspergillus niger,according to the proportion of 1.5∶1.0,with 10% of the inoculum amount,in the fermentation medium of 5%sweet potato protein,after 48 h fermentation,sweet potato protein can be hydrolyzed to small molecular peptides with DPPH·radical scavenging ability and reducing power.
complex bacteria;sweet potato proteins;bioactive peptides
TS239
A
10.16693/j.cnki.1671-9646(X).2016.11.031
1671-9646(2016)11b-0008-05
2016-09-21
西安市科技计划项目(NC1207(2));西安市未央区科技计划项目(201307)。
王 勇(1977—),男,博士,工程师,研究方向为农产品加工技术。
