亚热带森林群落草本植物对林下光环境的响应
2016-12-19胡砚秋柯娴氡徐明锋李文斌苏志尧
胡砚秋,柯娴氡,徐明锋,李文斌,苏志尧
(华南农业大学 林学与风景园林学院,广东 广州 510642)
亚热带森林群落草本植物对林下光环境的响应
胡砚秋,柯娴氡,徐明锋,李文斌,苏志尧
(华南农业大学 林学与风景园林学院,广东 广州 510642)
确定森林群落中林下草本植物的分布及其对光照的响应,不仅能揭示草本植物对光的生态需求,而且能为其引种和利用提供关键参考。在东源康禾自然保护区的常绿阔叶林内设置2 hm2样地,样地划分为50个20m×20m样方,在样方内设置小样方进行林下草本植物(包括禾本科竹亚科植物)调查。用半球面影像技术测定林下光照指标,进而分析草本植物分布与林下光环境的关系。结果表明,林下光环境的变异为草本植物的生长提供了多样的小生境,不同光照需求的物种在样地内均广泛分布;林下散射光与林下草本分布的除趋势对应分析(DCA)排序轴的相关性高于直射光与排序轴的相关性,说明林下散射光与林下植物的分布关系更为密切;不同光照总立地因子的样方组间草本植物的多度和丰富度没有显著差异,但非参数回归分析的结果清晰地反映了不同草本植物对光照响应的差别,如山麦冬Liriope spicata喜阴且不耐强光,黑莎草Gahnia tristis、箬竹Indocalamus tessellatus在不同光照条件下生长变化不大,说明物种对光照的变化不敏感、适应性更强。在草本植物的引种和利用过程中,可以结合目标地点的光照条件选择适生的物种。
草本植物;亚热带森林群落;林下光照;物种多样性
物种多样性是群落结构和群落功能复杂性的重要指标[1],也是群落生态环境的表征。草本植物是森林群落中重要的层次和组分,是群落物种多样性的重要组成[2-5]。同时,由于其独特的生态和景观功能,草本植物作为重要的地被绿化物种,在城市绿化建设中有着不可替代的作用[6-7]。在相对脆弱的城市绿地生态系统中,外来植物的大量使用会带来一定的生态风险[8-9],因此,乡土植物的重要性日渐凸显。与应用渐趋程序化的木本植物相比,草本植物尤其是乡土种类的应用有着更大的拓展空间[10-11]。相对于乔木,草本植物对于环境的变化更为敏感,前人研究表明,在小尺度环境下,林下光照条件是限制草本植物生长的关键因子[12-13]。林下光照因子的准确测定是研究林下植物格局与林下光照关系的关键环节,在已报道的研究中,林下光照条件既可通过仪器直接测定[14-15],也可以通过郁闭度、光照指数等指标来间接评价[16]。半球面影像技术作为一种近距离遥感的方法,已成为定量测定林下直射光和散射光指标的通用手段[17-19]。本研究以康禾自然保护区亚热带天然常绿阔叶林内的林下草本植物为研究对象,分析并模拟草本植物对林下光照因子的响应,以期量化乡土草本植物生长与林下光环境的关系,为林下植物的引种及乡土草本植物的开发利用提供理论依据。
1 研究地概况
研究样地位于广东东源康禾省级自然保护区(23 °44 ′~ 23 °53 ′ N, 115 °04 ′ ~ 115 °09 ′ E),保护区属于低山丘陵地带,山体呈东北-西南走向,区内最高点海拔839.7 m。气候属亚热带季风气候区,年均气温20.3~21.1 ℃,无霜期345~350 d,年均降水量2 142.6 mm。土壤类型主要为红壤、赤红壤,土层深厚,有机质积累丰富。保护区内有保存较完整的大片次生常绿阔叶林,林分上层主要优势种为米槠Castanopsis carlesii、木荷Schima superba、红背锥Castanopsis fargesii、杉木Cunninghamia lanceolata、鼠刺Itea chinensis等亚热带地带性树种。
2 研究方法
2.1 样地设置与植物调查
在保护区天然次生常绿阔叶林内设置面积为2 hm2的固定连续样地,并进行群落学调查,样地海拔200 ~315 m,坡度16.2°~44.5°,坡向163.9°~315.4°。按水平距离将整个样地划分为50个20m×20m的样方,在每个样方内设置5个2 m×2 m的小样方,分别位于样方中心点及对角线1/4、3/4处。记录小样方中草本植物的种名、多度(该物种在样方中的个体数)、平均高度。调查中,共有140个小样方记录到草本植物,分布在47个20m×20m的样方内。本项研究中,同时记录生长在林下的禾本科竹亚科物种,竹类虽然主干木质化,但茎的解剖结构与草本植物类同,而且一些竹类也是重要的绿化物种。野外调查在2013年1月进行。
2.2 林下光环境测定及分析
利用配置鱼眼镜头的数码相机(Nikon Coolpix 4500),在距地面1.6 m处,水平设置相机并采集林分冠层的半球面影像,以确定林下光照条件。拍照点的选取与草本植物小样方调查一致,以20m×20m的样方为调查单位,分别在样方中心及对角线的1/4、3/4处采集相片并编号。在GLA 2.0(Gap light analyzer)软件中处理冠层半球面影像图片,软件分析可直接获得林下直射光(Transmitted direct radiation, TDIR)、林下散射光(Transmitted diffuse radiation, TDIF)、林下总光照(Transmitted total radiation)以及林冠上界总光照(Above total radiation)等光照因子。某一样方的光照条件即为该样方中所采集的5张冠层影像分析所得数据的均值。
光照总立地因子(Total Site Factor, TSF)即林下总光照占林冠上界总光照的百分比。本研究按照样方的光照总立地因子将样方分为3组:第Ⅰ组,9.45%~14.63%;第Ⅱ组,15.15%~19.98%;第Ⅲ组,20.02%~29.43%。
2.3 数据统计与分析
利用STATISTICA 8.0软件进行Kruskal-Wallis检验,以检验组间林下草本植物多样性的差异。林下草本植物分布的除趋势对应分析(Detrended Correspondence Analysis, DCA)、林下光照因子与林下草本群落DCA排序轴的相关性分析在PCORD 6.0中完成。草本植物对光环境需求的模拟用非参数乘法回归法(NPMR, Non-Parametric Multiplicative Regression)在Hyperniche中完成。
3 结果与分析
3.1 林下光环境与草本植物组成
本次调查共记录林下草本植物11种,分属于6科。草本植物易受环境影响,分布变化较大,与重要值相比较,物种的多度及出现次数能更好的反映其分布状况[20],本研究选取这两个指标作为草本植物生长状况的指示。样地内多度较高的种类为黑莎草Gahnia tristis、箬竹Indocalamus tessellatus、山麦冬Liriope spicata和宽叶割鸡芒Hypolytrum nemorum。其中,黑莎草分布范围最广,在32个样方中出现,宽叶割鸡芒、山麦冬分别在19、15个样方中出现。其他种类为少数样方中出现的偶见种,物种多度较低,如弓果黍Cyrtococcum patens、短小蛇根草Ophiorrhiza pumila仅在一个样方中出现(见表1)。林下光环境总光照为7.18 ± 1.46 mol·m-2d-1,光照总立地因子为18.75% ± 3.82%,在一些林冠郁闭的林分中,仅有10%左右的光能够透过冠层到达林下。林下直射光略强于散射光,但变异系数也较大(见表2)。
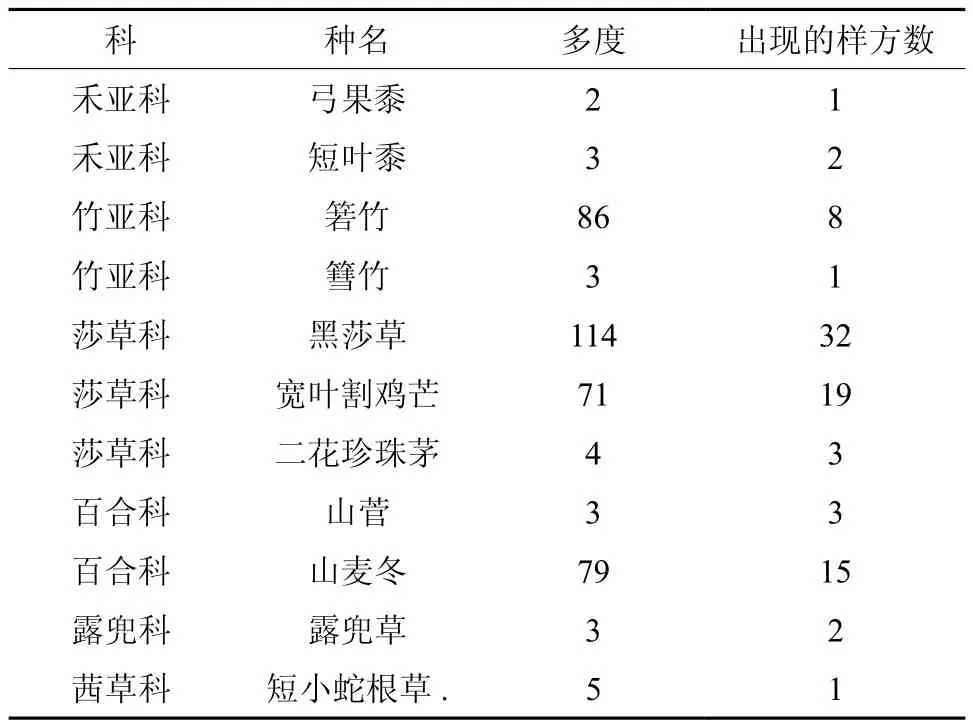
表1 林下草本植物组成Table 1 Species composition of the understory herbaceous plants
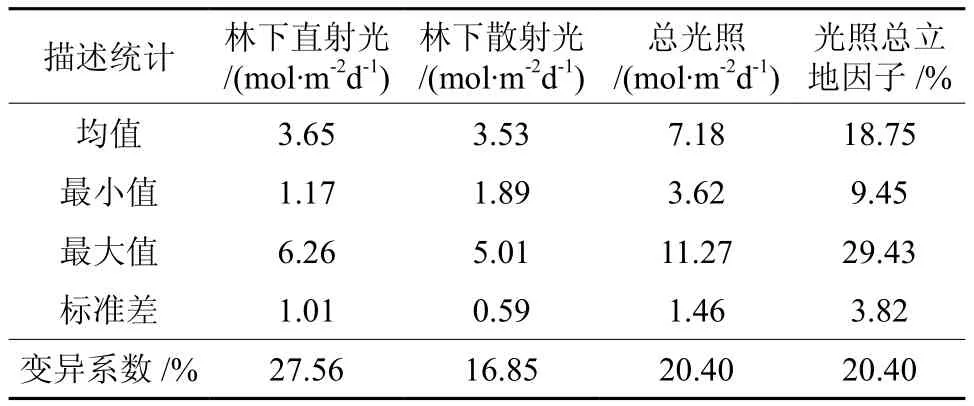
表2 林下光环境指标描述统计Table 2 Descriptive statistics of understory light environment
3.2 草本植物对林下光环境的响应
按照TSF的大小将全部样方分为3组,第Ⅲ组TSF值最大(20.02%~29.43%),包含样方数虽少于第Ⅱ组,但个体数最多,样方中位数也最大。方差分析结果表明,不同组别林下草本植物的个体数、丰富度的样方中位数之间的差异未达到显著水平(P>0.05)(见表3)。DCA排序反映了物种分布与环境因子的相关性,排序轴则代表了影响植物分布的生态因子的梯度[21-22]。分析林下光照与林下草本群落DCA排序轴的相关性,可以发现林下散射光与排序轴相关性更大,林下散射光与DCA第一轴、第二轴的Pearson’s相关系数分别为0.241、-0.210(见图1),而同等条件下直射光与两轴的相关系数仅为0.011、-0.014(见图2)。

表3 不同光照总立地因子下林下草本植物的多样性†Table 3 Herbaceous plants diversity in different TSF regimes

图1 林下散射光与DCA排序轴的相关性Fig. 1 Correlations of transmitted diffuse radiation with DCA ordination axis
3.3 草本植物光生境需求模拟
对个体数最多的4个物种的平均多度与其生长的光照条件进行非参数乘法回归分析,模拟物种生长对光生境的需求(见图3)。比较物种对光照环境的响应可以发现,山麦冬在林下总光照较小时,物种多度明显高于其他物种,耐阴性优于其他物种,当林下总光照高于10 mol·m-2d-1时,多度迅速下降,说明山麦冬是不耐强光环境的喜阴植物。山麦冬在较强光照下仍有较高丰富度则可能是因为箬竹的遮蔽,样地内箬竹与山麦冬共同出现的5个小样方中,其中4个小样方属于强光照条件(TSF第Ⅲ组)。箬竹耐阴性较强,但对强光照环境也有较好的适应性。宽叶割鸡芒在强光及弱光环境下平均多度均低,生长较差。黑莎草的多度随着总辐射的增加逐步增加,说明黑莎草较耐阴,同时在光照充足的环境下可以更好的生长。

图2 林下直射光与DCA排序轴的相关性Fig. 2 Correlations of transmitted direct radiation with DCA ordination axis

图3 物种平均多度与林下总光照的非参数回归分析Fig. 3 Non-parametric multiplicative regression for mean abundance against understory total radiation
4 结论与讨论
不同TSF条件下,林下草本植物的多度和丰富度没有显著差异,可能因为调查样地位于自然保护区内部,人为干扰少,林冠郁闭度高,林下草本植物总体多样性偏低,导致多度和丰富度整体变化幅度较小。调查中记录的草本植物中,弓果黍、短叶黍、山麦冬都是园林造景、城市绿化中常见的地被植物。喜阴的山麦冬,阳生性的箬竹,耐阴性较强的黑莎草、宽叶割鸡芒都可以在样地内广泛分布,表明森林中林下光环境变异较大,可以为不同光照需求的物种提供多样的小生境。
光照因子与林下草本群落DCA排序轴的相关性分析结果显示,DCA排序轴与散射光相关性高于与直射光的相关性,表明林下散射光与林下植物的分布关系更为密切。在太阳高度角不变的条件下,林下直射光主要取决于冠层的影响[23],林分冠层的变化导致林下直射光的变异程度高[24]。样地内林分保护良好,森林冠层郁闭度较高,直射光多被冠层拦截,因此散射光成为影响林下植物分布的重要因子。
对黑莎草等4个物种的非参数回归分析可以看出林下光照对不同草本植物生长的影响有着细微差别,回归曲线清晰的表明了物种不同的生态习性。已有研究表明林下光环境的微小变化就会导致生态位的进一步分化[25]。前人研究也提出,在研究林下植物时不应简单的将光环境分为林窗(gap)或是非林窗(non-gap)[26]。非参数回归可以很好的模拟某一范围内的光照条件变化,而不是人为划分的若干光照类型。物种的生态习性对于其绿化应用有着重要的指导作用,如山麦冬有着较好的耐阴性,在城市绿化中可以用于立交桥下、建筑中庭等光照较差的区域;在光照变化较大的构筑物周边,则可以选用对光照变化适应性强的黑莎草。
本研究采用传统群落学方法进行取样,调查数据具有代表性,能够反映群落中林下植物的整体情况,但一些个体数少的物种可能由于调查获得的数据量较小,导致物种生境需求模拟结果有一定局限性。在未来的研究中,可基于群落调查初步分析得到某些目标物种,之后针对特定物种开展调查,获取更丰富的数据对其生境需求进行模拟。在森林群落中,相较于草本植物而言,乔木对资源的利用、对生境变化的适应性都处于优势地位,正因如此,草本对于环境变化更为敏感,环境的微小差异也会影响其分布。在草本植物引种利用之前,可以结合自然生长的光照条件模拟预测其对更大范围光照的响应,对比待引入区域的环境条件,再确定是否引种或应用。
[1] 苏日古嘎, 张金屯, 王永霞. 北京松山自然保护区森林群落物种多样性及其神经网络预测[J]. 生态学报, 2013, 33(11):3394-3403.
[2] 李俊涛, 谭晓京, 蔡体久, 等. 凉水国家级自然保护区森林群落林下物种多样性[J]. 北京林业大学学报, 2007, 29(S2):266-271.
[3] 王 欢, 牛树奎, 绍 潇, 等. 森林生态系统内灌草植被生物量估测方法的研究[J]. 草业学报, 2014, 23(3): 20-29.
[4] 范文娟, 程积民, 张继敏, 等. 子午岭森林群落中草本植物的结构特征研究[J]. 水土保持通报, 2010, 30(2): 108-113.
[5] 崔 飞, 邓湘雯, 邓东华, 等. 炼山造林后不同年龄杉木人工林林下植物多样性动态[J]. 中南林业科技大学学报, 2015,35(6): 63-69.
[6] 詹姆斯·希契莫夫, 刘 波, 杭 烨. 城市绿色基础设施中大规模草本植物群落种植设计与管理的生态途径[J]. 中国园林,2013(3): 16-26.
[7] 张佳平. 云台山野生草本植物资源的园林开发利用评价[J].南京林业大学学报(自然科学版), 2013, 37(1): 37-43.
[8] 彭 程, 宿 敏, 周伟磊, 等. 北京地区外来植物组成特征及入侵植物分布[J]. 北京林业大学学报, 2010, 32(S1): 29-35.
[9] 陈 波, 包志毅. 建设城市森林需警惕生物入侵[J]. 城市问题,2004(5): 12-16.
[10] 张佳平, 丁彦芬. 连云港云台山野生草本植物资源调查、应用及保护研究[J]. 草业学报, 2012, 21(4): 215-223.
[11] 张 超, 徐 希, 李雪珂, 等. 北京奥林匹克森林公园乡土草本地被植物调查及分析[J]. 草业科学, 2012, 29(8): 1193-1198.
[12] 段文标, 王丽霞, 陈立新, 等. 红松阔叶混交林林隙大小及光照对草本植物的影响[J]. 应用生态学报, 2013, 24(3): 614-620.
[13] Bartemucci P, Messier C, Canham C D. Overstory influences on light attenuation patterns and understory plant community diversity and composition in southern boreal forests of Quebec[J]. Can J Forest Res, 2006, 36(9): 2065-2079.
[14] Posada J M, Lechowicz M J, Kitajima K. Optimal photosynthetic use of light by tropical tree crowns achieved by adjustment of individual leaf angles and nitrogen content [J]. Ann Bot-London,2009, 103(5): 795-805.
[15] Brantley S T, Young D R. Contribution of sun flecks is minimal in expanding shrub thickets compared to temperate forest [J].Ecology, 2009, 90(4): 1021-1029.
[16] Svenning J C. Crown illumination limits the population growth rate of a neotropical understorey palm (Geonoma macrostachys,Arecaceae) [J]. Plant Ecol, 2002, 159(2): 185-199.
[17] Jonckheere I, Fleck S, Nackaerts K, et al. Review of methods for in situ leaf area index determination-Part I. Theories, sensors and hemispherical photography [J]. Agr Forest Meteorol, 2004,121(1-2): 19-35.
[18] White M A, Asner G P, Nemani R R, et al. Measuring fractional cover and leaf area index in arid ecosystems: Digital camera,radiation transmittance, and laser altimetry methods [J]. Remote Sens Environ, 2000, 74(1): 45-57.
[19] Koch G W, Sillett S C, Jennings G M, et al. The limits to tree height [J]. Nature, 2004, 428(6985): 851-854.
[20] 范玉龙, 胡 楠, 丁圣彦, 等. 伏牛山自然保护区森林生态系统草本植物功能群的分类[J]. 生态学报, 2008, 28(7): 3092-3101.
[21] Ejrnæs R. Can we trust gradients extracted by Detrended Correspondence Analysis?[J]. J. Veg. Sci., 2000, 11(4): 565-572.
[22] Hill M O, Gauch Jr H G. Detrended correspondence analysis: an improved ordination technique [J]. Vegetatio, 1980, 42(1-3): 47-58.
[23] 区余端, 苏志尧. 粤北山地常绿阔叶林自然干扰后冠层结构与林下光照动态[J]. 生态学报, 2012, 32(18): 5637-5645.
[24] Gendron F, Messier C, Comeau P G. Comparison of various methods for estimating the mean growing season percent photosynthetic photon flux density in forests [J]. Agr Forest Meteorol, 1998, 92(1): 55-70.
[25] Svenning J. Small canopy gaps influence plant distributions in the rain forest understory [J]. Biotropica, 2000, 32(2): 252-261.
[26] Scanga S E. Population dynamics in canopy gaps: nonlinear response to variable light regimes by an understory plant [J].Plant Ecol., 2014, 215(8): 927-935.
Herbaceous plant in response to understory light regimes in a subtropical forest community
HU Yan-qiu, KE Xian-dong, XU Ming-feng, LI Wen-bin, SU Zhi-yao
(College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642, Guangdong, China)
Determining the distribution and ecological response of herbaceous plants for light has implications for revealing the demands of plants for radiation, as well as for species introduction and utilization. In this study, a 2 hm2plot was set up, which was further divided into fifty 20 m× 20 m subplots, for sampling herbaceous plant (including graminoid bamboo plants) abundance and distribution and measuring understory light regimes using hemispherical photography. Understory herbaceous plant patterns in relation to the light factors were analyzed based on the field collected data. The results showed that herbaceous species with different light demands distributed widely in the sample plots, suggested that the variation of understory light environment led to habitat heterogeneity. The correlation of transmitted diffuse radiation (TDIF) to detrended correspondence analysis (DCA) axis were much higher than that of transmitted direct radiation (TDIR), indicating that TDIF had greater influence in shaping the distribution of understory herbaceous plants. Herbaceous plant abundance and richness had no significant difference among the total site factors (TSF) groups, but non-parametric multiplicative regression (NPMR) analysis clearly reflected the different responses of herbaceous species to varied understory light conditions. For example, Liriope spicata preferred shady environment and was intolerant to strong radiation, while Gahnia tristis and Indocalamus tessellatus were more adaptable to the variation of light environment, the abundance of these two species had no pronounced changes under different light conditions. These results suggested that light conditions of habitat and species light adaptability should be taken into consideration in herbaceous plants introduction or utilization.
herbaceous plants; subtropical forest community; understory light; species diversity
S781.5
A
1673-923X(2016)08-0072-05
10.14067/j.cnki.1673-923x.2016.08.014
2015-12-28
广东省科技计划项目(2013B020305008);广东省林业科技创新专项资金项目(2011KJCX031-02)
胡砚秋,博士研究生
苏志尧,教授,博士,博士生导师;E-mail:zysu@scau.edu.cn
胡砚秋,柯娴氡,徐明锋,等. 亚热带森林群落草本植物对林下光环境的响应 [J].中南林业科技大学学报,2016, 36(8):72-76.
[本文编校:文凤鸣]
