亚热带4种森林生物量估算转换参数的研究
2016-12-19侯燕南吴惠俐项文化邓湘雯
侯燕南,吴惠俐,项文化,邓湘雯
(1. 中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2. 湖南省林业厅 造林处,湖南 长沙 410004)
亚热带4种森林生物量估算转换参数的研究
侯燕南1,2,吴惠俐1,项文化1,邓湘雯1
(1. 中南林业科技大学 生命科学与技术学院,湖南 长沙 410004;2. 湖南省林业厅 造林处,湖南 长沙 410004)
对我国亚热带森林资源调查中典型的4种森林类型(杉木林、马尾松林、落叶阔叶林和常绿阔叶林)的林分生物量数据进行整合分析,计算4种森林类型从林分蓄积量估算林分生物量的主要转换参数平均值,并分析影响转换参数的林分因子。结果表明:(1)杉木林、马尾松林、落叶阔叶林和常绿阔叶林4种森林类型中优势树种的木材基本密度平均值分别为0.313 3、0.412 5、0.502 1和0.527 4,木材基本密度因树种种源、种系、立地条件、林龄、林分密度等因子的不同而不同。(2)杉木林、马尾松林、落叶阔叶林和常绿阔叶林生物量扩展因子的平均值分别为1.308 9、1.265 4、1.423 3和1.391 3,根冠比的平均值分别为0.169 4、0.177 2、0.239 1和0.263 5。(3)4种森林类型的生物量扩展因子和根冠比随林龄、平均胸径和平均树高的增加而减少,随林分密度的增加而增加。4种森林类型的生物量估算转换参数间存在明显的差异,因此,在估算森林生物量时应按具体的森林类型进行估算,同时还应考虑林龄、林分密度、平均胸径和平均树高等林分因子的影响。
杉木林;马尾松林;落叶阔叶林;常绿阔叶林;木材密度;生物量扩展因子;林分因子
在过去约160年里,全球历史累计碳排放量为345 PgC(1 PgC=10亿tC)[1],全球碳平衡遭到破坏,严重影响着全球气候的变化。森林通过光合作用,吸收大气中的CO2并将其储存于植物的各个器官(根、茎、叶、花、果实和种子)中,在生物地球化学循环中发挥着“缓冲器”和“阀”的作用。森林在区域和全球碳循环中发挥着非常关键的作用[2]。据研究统计,全球森林约保存着陆地表面以上所有碳量的80%,以及陆地表面以下所有碳量的40%[3]。在过去几十年中,森林生物量的碳储量及其变化受到了高度的关注,从而相继制定了联合国气候变化框架公约(United Nations Framework Convention on Climate Change,UNFCCC)及其京都议定书(Kyoto Protocol),其要求各国估测并报道各国CO2的释放量和森林的碳储量,准确的碳汇数据对控制和减少CO2的释放具有重要的作用。
《土地利用、土地利用变化和林业优良作法指南》(LULUCF)这个指南内容指出CO2的释放与消除同其他温室气体不一样,其只能通过森林碳储量(即森林生物量)的变化量的间接方法来估算CO2的释放量与消除量[6]。森林碳储量估算有直接和间接两种方法[7]。直接方法为收获法,该方法尽管估算准确度高,但是对林分破坏性极大且费时费力,国内外学者对林分乔木层生物量的研究主要以收获法为主,而乔木则鲜少应用此方法[8]。间接方法主要有如下3种方法:生物量方程,生物量估算转换参数以及新兴的3S技术[9]。其中生物量转换参数法在区域森林生物量估算中被广泛使用,这也是IPCC《土地利用、土地利用变化和林业优良作法指南》推荐方法之一[6]。IPCC[10]中生物量估算转换参数包括生物量转换与扩展因子(Biomass conversion and expansion factor,Bcef)、生物量扩展因子(Biomass expansion factor,Bef)、根茎比(Root:shoot ratio,R)、基本木材密度(Basic wood density,Wd)等。
我国自1973年以来,先后进行了8次全国森林资源清查(The National Forest Resource Inventory,NFRI),森林资源清查结果提供了大量的森林蓄积量数据,却未对森林生物量的数据做详细报道[11]。目前,我国对亚热带地区森林生物量进行了大量的研究,也有大量的森林生物量研究方法[12-14],但是针对大区域的森林生物量研究方法甚少。将综合全国森林资源清查数据和IPCC中生物量估算参数,并利用已发表的亚热带森林生物量数据,试图解决如下两方面的问题:(1)确定亚热带地区4种森林类型(杉木林、马尾松林、落叶阔叶林和常绿阔叶林)的Wd、Bef和R及其平均值,并探讨不同森林类型Wd、Bef和R的差异,为大区域森林生物量估算奠定基础;(2)明确4种森林类型的生物量估算转换参数与林龄(Stand age,A)、林分密度(Stand density,D)、平均胸径(Diameter at breast height,Dbh)、平均树高(Height,H)林分因子的关系。
1 研究方法
1.1 数据来源
收集整理已公开发表或出版的4种亚热带森林类型的数据,主要数据包括:研究地概况(地点、经纬度、海拔、年均温、年均降水量)、树种、林分起源、林龄(A)、林分密度(D)、平均胸径(Dbh)、平均树高(H)、蓄积量(VTotal)、木材基本密度(Wd)、乔木树干(带皮)/树枝/树叶/根生物量(BS/BB/BL/BR)。其中,杉木林的生物量数据主要来自于16篇文献[14-29],马尾松林的数据主要来自于9篇文献[13,14,21,30-35],落叶阔叶林数据主要来源于9篇文献[12,29,36-42],常绿阔叶林数据主要来源于15篇文献[28,43-56],数据概况请见表1。4种森林类型的Wd数据在后文2.1中详述。
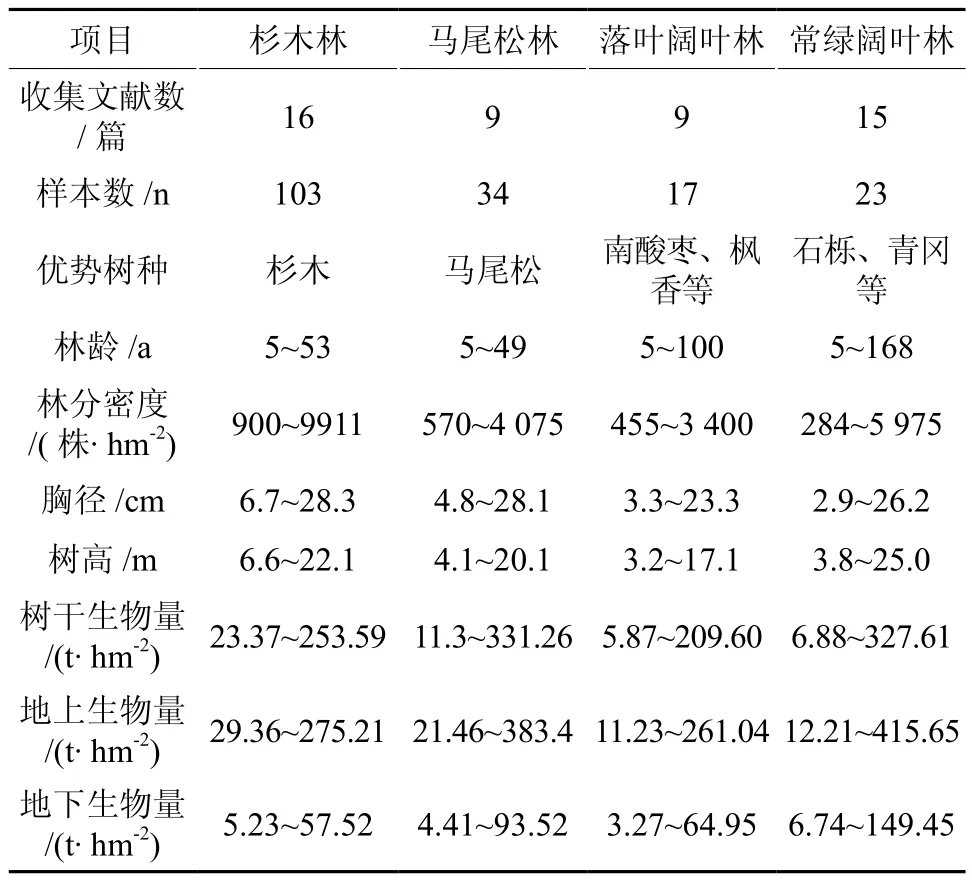
表1 亚热带4森林生物量相关数据的基本情况Table 1 Overview of stand biomass data collected from published literature for four subtropical forests
1.2 生物量估算转换参数的计算
依据IPCC[6,10],森林生物量(BTotal)计算公式如下:BTotal= VTotal×WD×BEF×(1+R)。式中:VTotal为林分蓄积量(m3),即指一定森林面积上存在着的林木树干部分的总体积;Wd为基本木材密度(mg/m3),Wd的意义即利用林分蓄积量推算林分树干生物量;Bef为林分地上生物量与树干生物量的比(无量纲),Bef的意义即利用林分树干生物量推算林分地上生物量; R为林分地下生物量与地上生物量的比(无量纲),R的意义即利用林分地上生物量推算林分地下生物量。
在我国森林资源清查报告中,尽管提供了不同森林类型不同林龄的森林面积及其林分蓄积量,但是对森林生物量的信息并未做详细的报道[11]。因此通过林分立木蓄积量计算林分生物量,必需先利用木材基本密度(Wd)来计算林分树干生物量,然后再利用生物量扩展因子(Bef)计算地上生物量,最后通过根冠比(R)获得林分总生物量。鉴于生物量估算转换参数Wd、Bef和R在区域森林生物量估算中的重要作用,因此,本研究利用收集到的 Wd、A、D、Dbh、H、BS/BB/BL/BR和 VTotal数据,根据上述参数的定义计算4种森林类型的Wd、Bef和R及其平均值。
1.3 数据分析
应用Excel计算各生物量估算转换参数的值。应用Origin 8.0软件作各森林生物量估算转换参数的相对频率分布图,拟合各参数与A、D、Dbh、H的回归曲线,并分析其相关性。
2 生物量估算转换参数分析
2.1 木材基本密度
根据收集到的基本木材密度(Wd)的数据,计算4种森林类型优势树种Wd的平均值,结果如下:杉木林优势树种为杉木,其Wd平均值为0.313 3;马尾松林优势树种为马尾松,其Wd平均值为0.412 5;落叶阔叶林优势树种有枫香、拟赤杨等落叶阔叶树,其Wd平均值为0.502 1;常绿阔叶林优势树种有木荷、木荚红豆等常绿阔叶树,其Wd平均值为0.527 4。4种森林类型的R平均值大小顺序为:常绿阔叶林>落叶阔叶林>马尾松林>杉木林。各森林类型优势树种的木材基本密度不仅因优势树种的不同而不同,而且受树种的种源、家系、立地条件、林龄、林分密度[57-60]等因子的影响。如杉木不同种源间木材的基本密度有极显著的遗传差异,杉木8年生幼苗在山坡、山谷和山洼不同立地条件下木材密度也不同[57]。不同林龄龄组下4种森林类型各优势树种Wd的平均值请见表2,结果分析发现所有优势树种的Wd随着年龄的增加而增加,增加到最大值后,随着年龄的增加又减少。
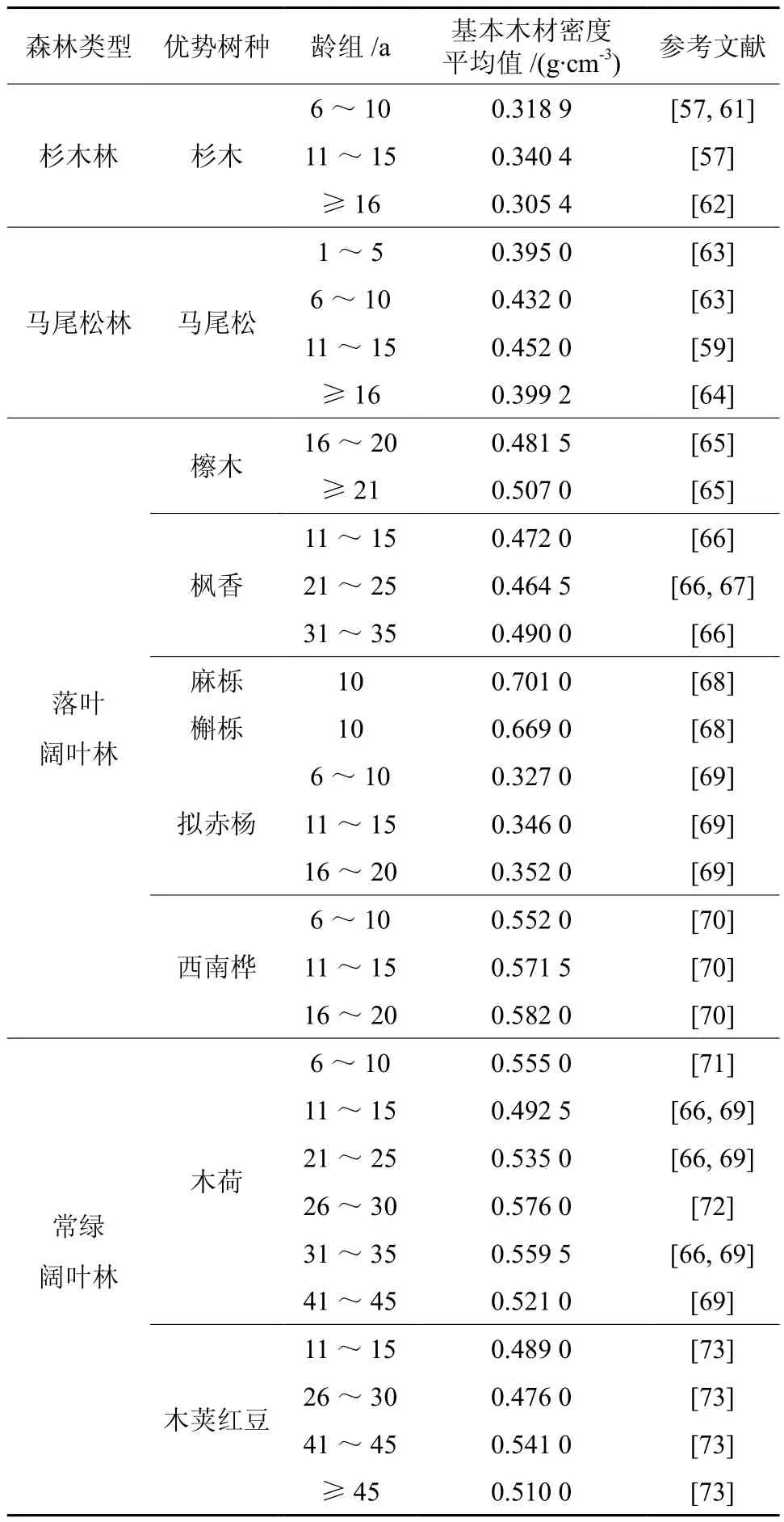
表2 部分亚热带森林优势树种基本木材密度情况Table 2 The wood basic density information of dominant tree species in subtropical forests
2.2 生物量扩展因子
本研究的4种亚热带森林类型的Bef平均值分别为:杉木林为1.308 9(n=103,SD=0.105 8),马尾松林为1.265 4(n=34,SD=0.104 1),落叶阔叶林为1.423 3(n=17,SD=0.234 0),常绿阔叶林为 1.391 3(n=23,SD=0.168 4)。4种森林类型的Bef平均值大小顺序为:落叶阔叶林>常绿阔叶林>杉木林>马尾松林。4种森林类型Bef值分布情况如图1所示,其中杉木林Bef值集中分布于1.25~1.35,马尾松林Bef值主要分布于1.1~1.4,落叶阔叶林Bef值集中分布于1.3~1.4,常绿阔叶林Bef值主要分布于1.2~1.5。
4种森林类型的Bef与林龄(A)、平均胸径(Dbh)、平均树高(H)呈负相关性,与林分密度(D)呈正相关性(图2)。通过利用常见的对数函数、二项式函数、幂函数和指数函数进行回归分析,发现4种森林类型的Bef与各林分因子的拟合函数形式不一,结果请见表3。4种森林类型的Bef与Dbh和H两个林分因子的拟合效果较好,Bef与A和D的拟合效果较差。
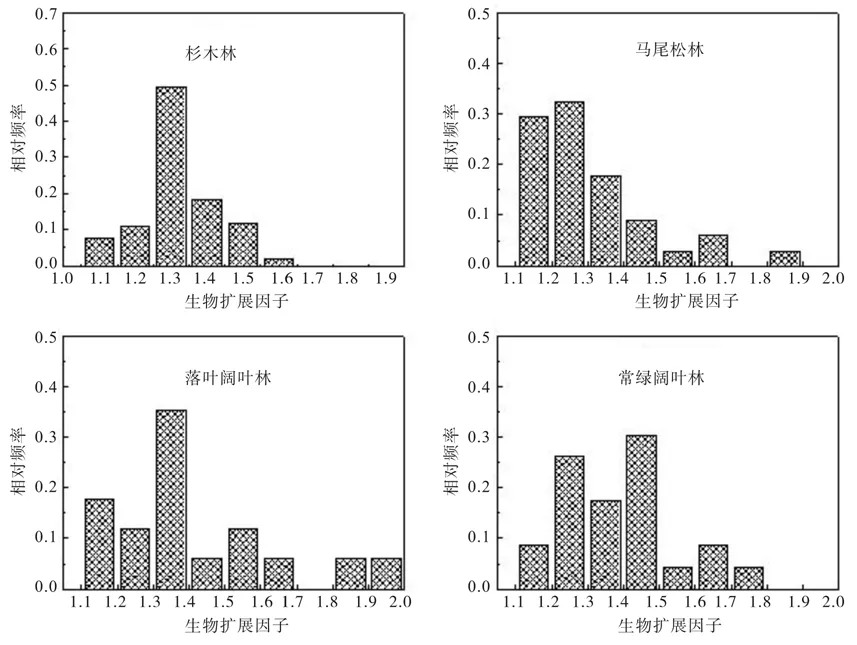
图1 4种森林类型Bef值相对频率分布Fig. 1 The relative frequency distribution of Bef value in four forest
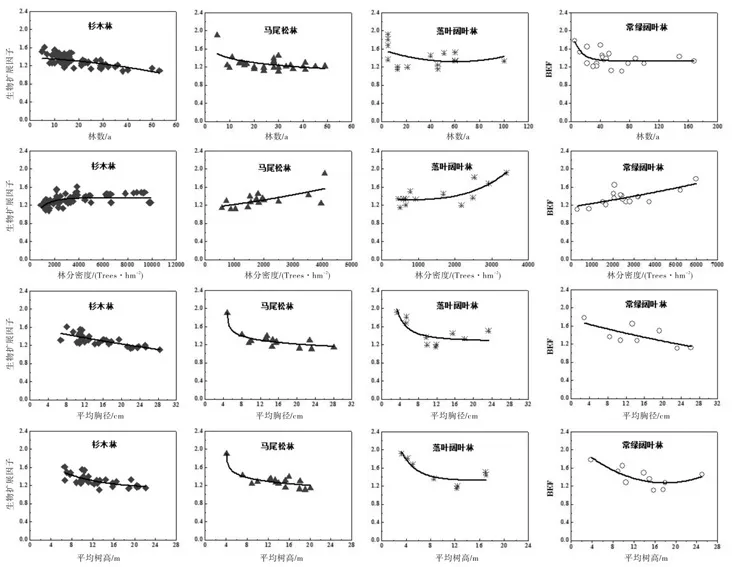
图2 生物量扩展因子(Bef)与林龄(A)、林分密度(D)、平均胸径(Dbh)、平均树高(H)的关系Fig. 2 Biomass expansion factor in relation to stand Age (A), stand density (D), diameter at breast height (Dbh) and height (H).
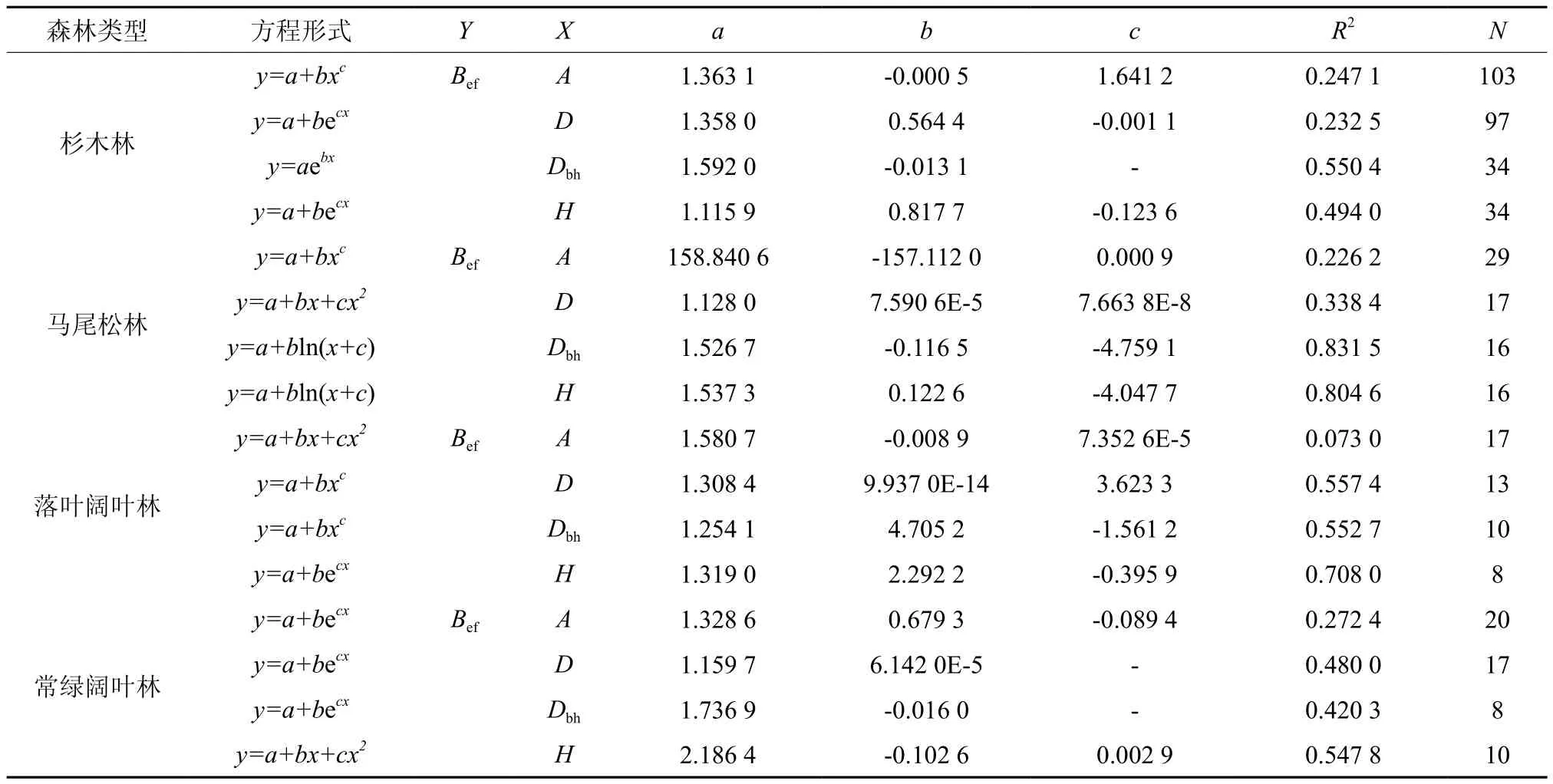
表3 Bef与A、D、Dbh和H的回归方程Table 3 Equations for Bef in relations to A, D, Dbh and H
2.3 根茎比
本研究的4种亚热带森林类型的R平均值分别为:杉木林为0.169 4(n=103,SD=0.067 1),马尾松林为0.177 2(n=29,SD=0.089 5),落叶阔叶林为0.239 1(n=11,SD=0.081 3),常绿阔叶林为0.263 5(n=19,SD=0.111 5)。4种森林类型的R平均值大小顺序为:常绿阔叶林>落叶阔叶林>马尾松林>杉木林。4种森林类型的R值分布情况如图3所示,其中杉木林R值集中分布于0.10~0.15,马尾松林R值集中分布于0.10~0.20,落叶阔叶林R值主要分布于0.15~0.30,常绿阔叶林R值主要分布于0.1~0.4。
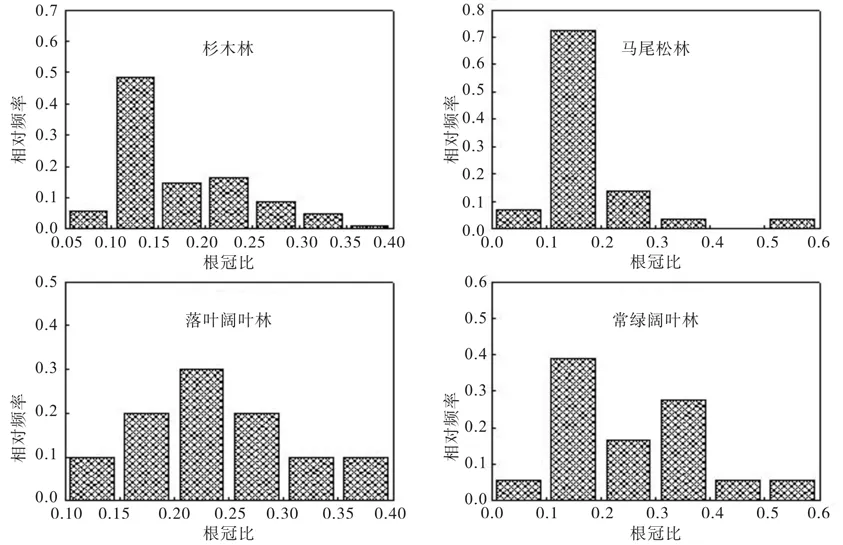
图3 4种森林类型根冠比(R)值相对频率分布Fig. 3 The relative frequency distribution of root:shoot ratio (R) value in four forest
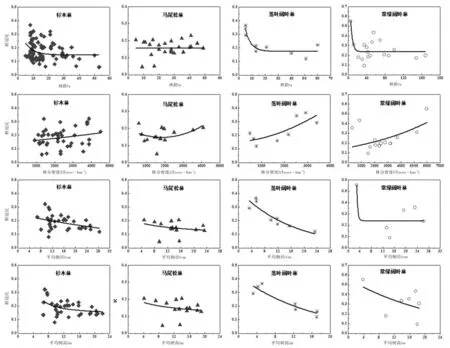
图4 根冠比(R)与林龄(A)、林分密度(D)、平均胸径(Dbh)、平均树高(H)的关系Fig. 4 Root:shoot ratio (R) in relation to stand age (A), stand density (D), diameter at breast height (Dbh) and height (H).
4种森林类型的R随着A、Dbh和H的增加而减少(马尾松林的R随A的变化而无明显的变化趋势),随D的增加而增加(图4)。通过利用上述的对数函数、二项式函数、幂函数和指数函数进行回归分析,发现4种森林类型的R与A、D、Dbh和H这4个林分因子的拟合函数形式不一(表4)。尽管4种森林类型的R与上述4个林分因子具有一定的相关性,但除了落叶阔叶林与之相关性较强外,其他3种森林类型与之相关性均较弱。
3 结论与讨论
(1)本研究计算了亚热带地区4种森林类型优势树种基本木材密度的平均值,分别为:杉木林0.313 3,马尾松林0.412 5,落叶阔叶林0.502 1,常绿阔叶林0.527 4。除树种间的密度具有差异外,优势树种的种源、种系、立地条件、林分密度、林龄等因子也密切影响着基本木材密度。IPCC[10]中列出了不同气候带不同树种基本木材密度的缺省值,而IPCC中给定的基本木材密度缺省值为固定的数值,如果利用IPCC缺省值来估算我国亚热带森林生物量,其结果可能大大偏离实际生物量。因此,在利用基本木材密度估算森林树干生物量时,应同时考虑不同森林类型影响因子的影响,根据具体的森林类型选择合适的基本木材密度,以减少估算误差。
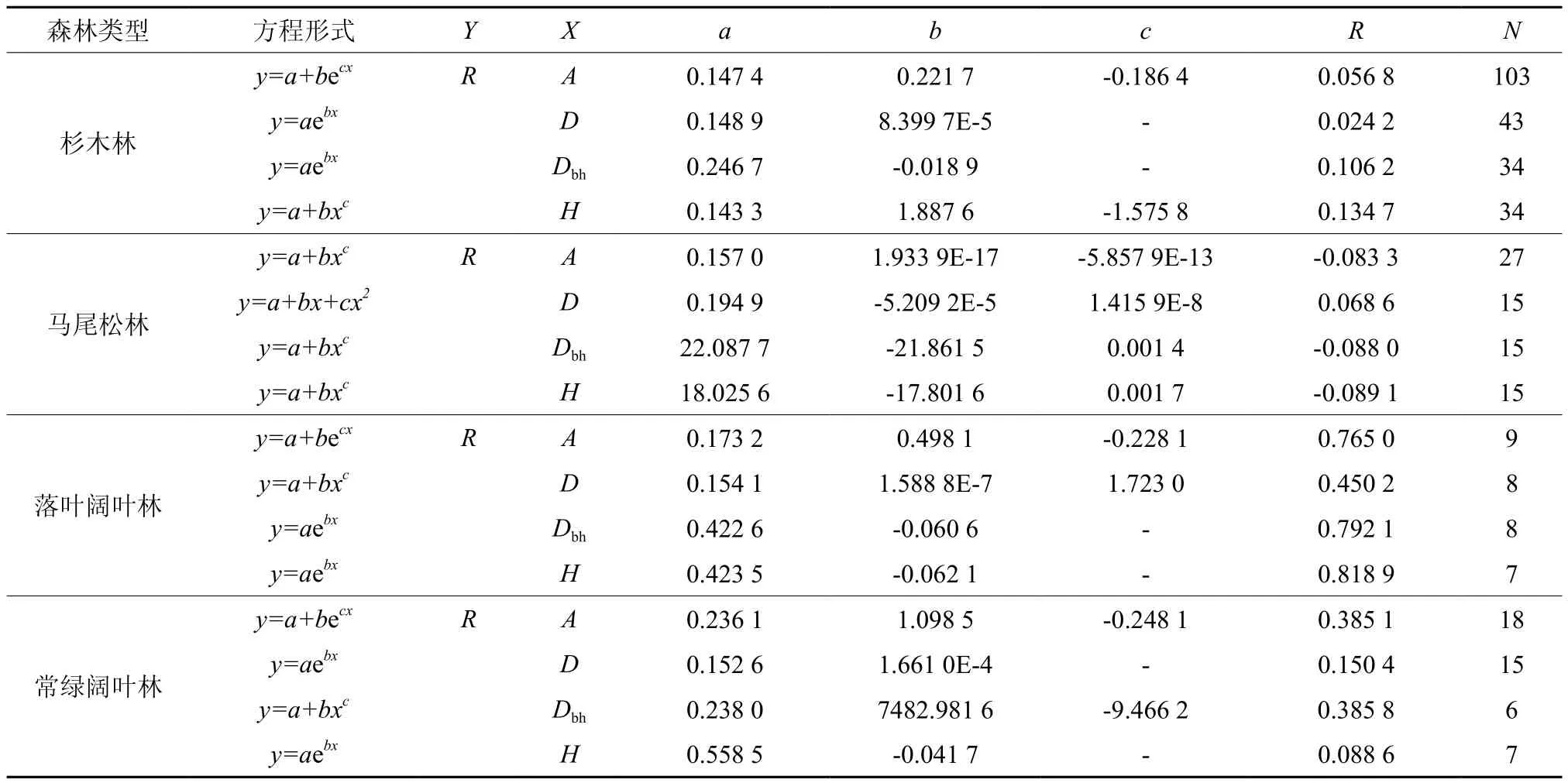
表4 R与A、D、Dbh和H的回归方程Table 4 Equations for R in relations to A, D, Dbh and H.
(2)IPCC[10]给出了不同气候带不同森林类型生物量扩展因子生物量扩展因子的缺省值,针叶林的生物量扩展因子平均值为1.300 0,阔叶林的生物量扩展因子平均值为1.400 0。本研究计算的杉木林和马尾松林的生物量扩展因子均值分别为1.308 9和1.265 4,比IPCC缺省值分别大0.008 9和小0.034 6;落叶阔叶林和常绿阔叶林的生物量扩展因子平均值分别为1.423 3和1.391 3,与IPCC阔叶林缺省值相比,分别大0.023 3和小0.008 7。IPCC也给出了根冠比的缺省值,根据不同植被类型的地上部生物量选取相应的值,亚热带旱林的根冠比平均值为0.270 0。本研究计算的杉木林、马尾松林、落叶阔叶林和常绿阔叶林的根冠比平均值分别为0.169 4、0.177 2、0.239 1和0.263 5,比IPCC缺省值分别小0.100 6、0.092 8、0.030 9和0.006 5。通过上述比较分析可知,如果只选择IPCC缺省值来计算我国亚热带森林的生物量,其结果可能会高估或低估了各森林生物量,因此,应根据不同气候带不同森林类型的具体情况,制定具体的准确的生物量估算转换参数。
(3)不同森林类型的生物量估算转换参数之间存在明显的差异,造成这种差异的原因可能有:①不同森林类型组成树种不同,其立木蓄积量和森林生物量差异显著。浙江龙泉针叶林立木蓄积年增长量是阔叶林的2.7倍,森林活生物量是阔叶林的3.19倍[74],这将造成生物量估算转换参数的差异。②不同森林类型的立地条件不同,如土壤、光照等。本研究中杉木林和马尾松林多为人工林,多种植于土壤肥沃、地力条件佳的土地,其生长所需的养分充足而扎根较浅,而阔叶林多为天然次生林,土地相对贫瘠,其因生长所需的养分较缺乏而扎根较深,所以杉木林和马尾松林的根生物量相对较低,地上部分生物量较高,又由于其林木生长所需营养充足,生长速度较快,从而导致基本木材密度、生物量扩展因子、根冠比的平均值较低。③不同森林类型受林龄、林分密度、胸径、树高等林分因子不同程度的影响。本研究中4种森林类型的生物量扩展因子和根冠比随林龄、胸径和树高的增加而减少,随林分密度的增加而增加,这一结果与Lehtonen等人研究结果一致[75]。其中生物量扩展因子与胸径和树高的拟合效果优于与林龄和林分密度的拟合效果,说明4种森林类型的生物量扩展因子与胸径和树高的相关性较强,受其影响较大。4种森林类型的根冠比与林龄、林分密度、胸径、树高的拟合效果除落叶阔叶林外均较差,表明根冠比受这4个林分因子的影响较小。本研究表2分析结果发现,4种森林类型优势树种的基本木材密度随着年龄的增加呈先增加后降低的趋势。
森林生物量估算转换参数在区域森林生物量估算过程中发挥着重要的作用,然而关于我国亚热带气候区森林生物量转换参数的研究甚少。目前,亚热带不同区域森林生物量的研究情况不一,一些区域研究较全面,而另一些区域研究较薄弱,且不同研究者采用的生物量估算方法不一致而导致估算结果不一致,从而不可避免的导致本研究的生物量估算参数值与实际值之间存在一定的误差。因此,应加强对我国亚热带气候区森林生物量及生物量估算参数的进一步研究,主要包括如下几点:①亚热带气候区典型森林类型优势树种的基本木材密度数据总体来说较缺乏,应加强对其研究;②应对亚热带气候区不同地区的森林进行全面的研究,以获得更准确的生物量估算参数;③不同研究者应加强森林生物量研究的交流,尽可能的减少因利用的估算方法和参数定义的不一致而造成估算结果的差异。
[1] 朱江玲,岳 超,王少鹏,等. 1850—2008年中国及世界主要国家的碳排放—碳排放与社会发展Ⅰ[J]. 北京大学学报,2010, 46(4): 497-504.
[2] 项文化. 杉木林采伐迹地人工造林与自然恢复的生态功能过程研究[D]. 长沙: 中南林业科技大学, 2003.
[3] 续珊珊,姚顺波. 基于生物量转换因子法的我国森林碳储量区域差异分析[J]. 北京林业大学学报:社会科学版, 2009,8(3): 109-4.
[4] Cannell M G R. World forest biomass and primary production data[M]. London: Academic press, 1982.
[5] Reichle D E. Dynamic properties of forest ecosystems[M].London: Cambridge University Press, 1982.
[6] IPCC. Good Practice Guidance for land use, Land-use Change and Forestry[R]. Japan: Institute of Global Environmental Strategies, 2003.
[7] West W P. Tree and Forest Measurement[M]. Berlin: Springer-Verlag, 2004.
[8] 巨文珍,农胜奇. 森林生物量研究进展[J]. 西南林业大学学报, 2011, 31(2): 78-83.
[9] Somogyi Z, Cienciala E, Mäkipää R, et al. Indirect methods of large-scale forest biomass estimation[J]. European Journal of Forest Research, 2007, 126(2): 197-207.
[10] IPCC. IPCC Guidelines for National Greenhouse Gas Inventories: Agriculture, Forestry and other Land use[R]. Japan:Institute of Global Environment Strategies, 2006.
[11] Fang J Y, Chen A P, Peng C H, et al. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science,2001, 292(5525): 2320-2322.
[12] 沈 燕. 亚热带天然次生混交林生物量及养分生物循环研究[D]. 长沙: 中南林业科技大学, 2011.
[13] 杨清培, 李鸣光, 王伯荪, 等. 粤西南亚热带森林演替过程中的生物量与净第一性生产力动态[J]. 应用生态学报, 2003,14(12): 2136-2140.
[14] 杜 虎, 曾馥平, 王克林, 等. 中国南方3种主要人工林生物量和生产力的动态变化[J]. 生态学报, 2014, 34(10): 2712-2724.
[15] 周晓峰. 中国森林生态系统定位研究[D]. 田大伦, 潘维俦,雷志星,等. 哈尔滨: 东北林业大学出版社, 1994.
[16] 田大伦, 盘宏华, 康文星, 等. 第二代杉木林生物量的研究[J].中南林学院学报, 1998, 18(3):11-16.
[17] 赵 坤, 田大伦. 会同杉木林成熟阶段生物量的研究[J]. 中南林学院学报, 2000, 20(4):7-13.
[18] 温远光, 梁宏温, 蒋海平. 广西杉木林生物量及分配规律的研究[J]. 广西农业大学学报, 1995, 14(1): 55-64.
[19] 闫文德, 田大伦, 何功秀. 湖南会同第2代杉木林乔木层生物量的分布格局[J]. 林业资源管理, 2003, 2(2): 5-12.
[20] 黄贤松, 吴承祯, 洪 伟, 等. 两种杉木林密度与立木生物量的研究[J]. 福建林学院学报 2012, 31(2): 102-105.
[21] 张 林, 黄 永, 罗天祥, 等. 林分各器官生物量随林龄的变化规律-以杉木、马尾松人工林为例[J]. 中国科学院研究生学报, 2005, 22(2): 170-178.
[22] 段爱国, 张建国, 何彩云, 等. 杉木林生物量变化规律的研究[J]. 林业科学研究, 2005, 18(2): 125-132.
[23] 俞月凤, 宋同清, 曾馥平, 等. 杉木林生物量及其分配的动态变化[J]. 生态学杂志, 2013, 32(7): 1660-1666.
[24] 叶镜中, 姜志林. 苏南丘陵杉木林的生物量结构[J]. 生态学报, 1983, 3(1): 7-14.
[25] 应金花, 何宗明, 范少辉, 等. 一代杉木林( 29年生 )林分生物量结构[J]. 福建林学院学报, 2001, 21(4): 339-342.
[26] 李淑花. 第二代杉木林生物量的估测与分布特征研究[D]. 长沙: 中南林业科技大学, 2006.
[27] 吕德文. 不同经营模式杉木林的生物量研究[J]. 山东林业科技, 2011(5): 36-9.
[28] 刘爱琴, 刘春华, 马庆祥. 福建青冈人工林的生物生产力研究[J]. 福建林学院学报, 2004, 24(4): 294-297.
[29 何贵平, 陈益泰, 余元华, 等. 南酸枣人工林早期生长特性及其与杉木混交效应研究[J]. 林业科学研究,2004,17(2):206-212.
[30] 郭久江. 福州北郊木荷林与马尾松林生物量和能量的研究[J].林业科技开发, 2013(17):51-4.
[31] 吴守蓉, 杨惠强, 洪 蓉, 等. 马尾松林生物量及其结构的研究[J]. 福建林业科技, 1999, 26(1): 18-21.
[32] 许 雯, 胡海波, 周长海. 皖东地区马尾松林生物量结构及其分布特征[J]. 中南林业科技大学学报, 2011, 36(1): 112-5.
[33] 樊后保, 李燕燕, 黄玉梓. 马尾松-细叶青冈混交林的生物量及其生产力结构[J]. 中南林学院学报, 2005, 25(6): 38-41.
[34] 岳军伟, 龙 蔚, 刘光正, 等. 红壤低丘马尾松低效林生物量特征[J]. 林业科技开发, 2010, 24(2): 19-22.
[35] 徐世红. 马尾松阔叶树混交林生物量和矿质养分特征[D]. 合肥: 安徽农业大学, 2010.
[36] 沈 燕, 田大伦, 项文化, 等. 天然次生檫木枫香混交林生物量及生产力研究[J]. 中南林业科技大学学报, 2011, 31(5): 26-30.
[37] 易利萍, 文仕知, 王珍珍, 等. 枫香人工林的生物产量及生产力[J]. 中南林业科技大学学报, 2008, 28(2): 50-53.
[38] 黄勇来. 枫香与不同树种混交林的生长及生物生产力研究[J].西南林学院学报, 2006, 26(2): 15-8.
[39] 王向雨, 胡 东, 贺金生. 神农架地区米心水青冈林和锐齿槲栎林生物量的研究[J]. 首都师范大学学报, 2007, 28(2): 62-67.
[40] 王卫斌. 南桦人工群落特征研究[D]. 昆明: 云南大学, 2011.
[41] 郝焰平. 青阳县栎类阔叶林生物量与优势树种生长规律研究[D]. 合肥: 安徽农业大学, 2009.
[42] 黄庆丰. 皖南低山天然次生阔叶林林分结构与优势树种地上生物量[D]. 合肥: 安徽农业大学, 2011.
[43] 陈洁如, 江泽平, 梅建波. 长沙县红壤丘陵石栎次生林的调查研究[J]. 中南林学院学报, 1987, 2(12): 23-26.
[44] 李铭红. 40龄青冈林林木层的生物量和净生产量研究[J]. 浙江师大学报. 2000, 23(2): 186-189.
[45] 黄典忠. 闽江下游福建青冈次生林群落的生物量特征[J]. 防护林科技, 2006 (1): 16-18.
[46] 杨同辉, 宋 坤, 达良俊, 等. 中国东部木荷-米槠林的生物量和地上净初级生产力[J]. 中国科学, 2010, 40(7): 610-619.
[47] 杨同辉, 达良俊, 宋永昌, 等. 浙江天童国家森林公园常绿阔叶林生物量研究(Ⅰ)群落结构及主要组成树种生物量特征[J].浙江林学院学报, 2005, 22(4): 363-369.
[48] 杨同辉, 达良俊, 李修鹏. 浙江天童国家森林公园常绿阔叶林生物量研究(Ⅱ)群落生物量及其分配规律[J]. 浙江林学院学报, 2007, 24(4): 389-395.
[49] 李 根. 南岭小坑流域三种次生常绿阔叶林地上部分生物量研究[D]. 北京: 中国林业科学研究院, 2011.
[50] 张 林, 罗天祥, 邓坤枚, 等. 广西黄冕林场次生常绿阔叶林生物量及净第一性生产力[J]. 应用生态学报, 2004, 15(11):2029-2033.
[51] 郭久江. 福州北郊木荷林与马尾松林生物量和能量的研究 [J].林业科技开发, 2013(17): 51-54.
[52] 谢寿昌, 刘文耀, 李寿昌, 等. 云南哀牢山中山湿性常绿阔叶林生物量的初步研究[J]. 植物生态学报, 1996, 20(2): 167-176.
[53] 党承林, 吴兆录. 季风常绿阔叶林短刺栲群落的生物量研究[J].云南大学学报, 1992, 14(2): 95-107.
[54] 党承林, 吴兆录, 张 泽. 黄毛青冈群落的生物量研究[J]. 云南大学学报, 1994, 16(3): 205-9.
[55] 李 冬. 西双版纳季风常绿阔叶林的碳储量及其分配特征研究[D]. 北京: 中国科学院研究生院, 2006.
[56] 陈章和, 张宏达, 王伯荪, 等. 广东黑石项常绿阔叶林生物最及其分配的研究[J]. 植物生态学报与地植物学学报, 1993,17(4): 289-298.
[57] 李晓储, 黄立斌, 王 伟, 等. 杉木木材基本密度变异的研究[J].林业科学研究, 1999, 12(2): 179-84.
[58] 苏兵强. 杉木优良家系木材密度及生长遗传稳定性分析[J].福建林业科技, 2001, 28: 1-4.
[59] 连辉明, 曾令海, 何波祥, 等. 马尾松林木材基本密度、管胞长度遗传变异规律及其与生长量、产脂力的相关性研究[J].广东林业科技, 2006, 22(1): 5-12.
[60] 洪昌瑞, 沈辛作, 陈天霞, 等. 杉木种源木材密度的遗传变异与选择[J]. 浙江林学院学报, 1992, 9(3): 246-52.
[61] 胡德活, 阮梓材, 钱志能, 等. 杉木无性系木材密度遗传变异及其与生长性状的相关性[J]. 中南林学院学报, 2004, 24(5): 24-7.
[62] 林金国, 黄清峰, 林志高. 坡相对杉木林和湿地松人工林木材密度的影响[J]. 四川农业大学学报, 1998, 16(1):118-21.
[63] 毛 桃. 马尾松优树子代测定林生长和材质的遗传分析及联合选择[D]. 南京: 南京林业大学, 2007.
[64] 储德裕, 刘青华, 金国庆, 等. 马尾松优树无性系生长、干形和木材密度评价[J]. 浙江林业科技, 2010, 30(5): 46-52.
[65] 鄢洪星, 王洪友, 李党法, 等. 豫南引种檫木木材物理力学性质的试验研究[J]. 河南林业科技, 2007, 27(2): 1-5.
[66] 周侃侃. 不同年龄枫香、木荷木材性质及加工利用适应性研究[D]. 杭州: 浙江林学院, 2009.
[67] 吴远彬. 枫香人工林和天然林木材物理力学性质比较[J]. 附件林业科技, 2002, 29(3): 37-9.
[68] 刘志龙, 虞木奎, 方升佐, 等. 壳斗科 4个树种木材基本密度及燃烧值的变异分析[J]. 江西农业大学学报, 2009, 31(4):674-8.
[69] 林金国. 福建中亚热带人工阔叶林材质与林学因子关系研究[D]. 南京: 南京林业大学, 2005.
[70] 孙庆丰, 陈太安, 王昌命. 不同树龄西南桦材性差异研究[J].西南林学院学报, 2006, 26(6): 79-81.
[71] 王秀花, 陈柳英, 马丽珍, 等. 7年生木荷生长和木材基本密度地理遗传变异及种源选择[J]. 林业科学研究, 2011, 24(3):307-313.
[72] 王秀花, 马雪红, 金国庆, 等. 木荷天然林分个体类型及材性性状变异[J]. 林业科学, 2011, 47(3): 134-139.
[73] 彭 彪, 谢世春, 彭东辉, 等. 木荚红豆树木材密度和纤维形态的径向变异[J]. 安徽农业科学, 2010, 38(17): 9326-9328.
[74] 叶庭旺, 冯建国, 樊子才, 等. 浙江龙泉阔叶林与针叶林的碳汇功能差异浅析[J]. 江西林业科技, 2013(2): 47-49.
[75] Lehtonen A, Mäkipää R, Heikkinen J, et al. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests[J]. Forest Ecology and Management, 2004, 188(1-3): 211-224.
Conversion parameters determination for stand biomass estimation of four subtropical forest types based on national forest inventory system
HOU Yan-nan1,2, WU Hui-li1, XIANG Wen-hua1, DENG Xiang-wen1
(1. School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, Hunan, China;2. Silivculture Administration Section, Forestry Department of Hunan Province, Changsha 410004, Hunan, China)
Synthesis of stand biomass data of 4 typical forests (Cunninghamia lanceonata forest, Pinus massoniana forest, deciduous broadleaved forest and evergreen broadleaved forest) from National Forest Inventory in subtropical area, we determine conversion parameters for stand biomass estimation based on stand volumes and analyzed how stand characteristics affect the parameters. The results showed that: (1) The mean values of wood basic density (Wd) of dominant trees of Cunninghamia lanceonata forest, Pinus massoniana forest, deciduous broadleaved forest and evergreen broadleaved forest were 0.313 3, 0.412 5, 0.502 1 and 0.527 4,respectively. The Wd was affected by tree provenance, species, site conditions, stand age (A), stand density(D) and other factors. (2) The mean values of biomass expansion factor (Bef) of Cunninghamia lanceonata forest, Pinus massoniana forest, deciduous broadleaved forest and evergreen broadleaved forest were 1.308 9, 1.265 4, 1.423 3 and 1.308 9, respectively, and the mean values of root: shoot ratio (R) were 0.169 4, 0.177 2, 0.239 1 and 0.263 5, respectively. (3) The Bef and R values of these four forests were increased with the increases of A, average diameter at breast height (Dbh) and average tree height (H), and reduced with the increases of D, excepted the R values of the Pinus massoniana forest had no obvious change with A. Due to obvious differences between these four forests, so we should select conversion parameters according to specific forest when estimating forest biomass.
Cunninghamia lanceonata forest; Pinus massoniana forest; deciduous broadleaved forest; evergreen broadleaved forest;wood basic density; biomass expansion factor; stand factor
S785
A
1673-923X(2016)08-0057-09
10.14067/j.cnki.1673-923x.2016.08.012
2015-12-30
国家林业公益性行业专项项目(201304317)
侯燕南,高级工程师,博士研究生
项文化,教授,博士生导师;E-mail:xiangwh2005@163.com
侯燕南,吴惠俐,项文化,等. 亚热带4种森林生物量估算转换参数的研究[J].中南林业科技大学学报,2016,36(8):57-65.
[本文编校:文凤鸣]
