文冠果叶碳氮含量对人为拉枝干扰的响应机制
2016-12-19苏曼琳马履一苏淑钗
苏曼琳 ,马履一 ,段 劼 ,吴 尚,苏淑钗 ,敖 妍
(1.北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083;2.国家能源非粮生物质原料研发中心,北京 100083)
文冠果叶碳氮含量对人为拉枝干扰的响应机制
苏曼琳1,2,马履一1,2,段 劼2,吴 尚1,2,苏淑钗1,2,敖 妍1,2
(1.北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083;2.国家能源非粮生物质原料研发中心,北京 100083)
碳氮含量与花芽分化有密切的关系,枝条着生角度影响了碳氮含量在叶片中的分布,进而影响了花芽分化。为了提出有利于花芽分化的枝条着生角度,对文冠果样树的枝条分别进行了45 °、60 °、90 °角度的拉枝处理,不拉枝为对照,分析拉枝后叶片的可溶性糖含量、淀粉含量、氮含量和碳氮比。结果表明:拉枝角度为60 °和90 °枝条的叶片可溶性糖、淀粉含量和碳氮比都显著高于30 °和45 °枝条(p<0.05),氮含量则相反(p<0.05)。但是30 °和45 °之间,60 °和90 °之间没有显著差异(p>0.05)。综合分析碳氮含量、成花结果和枝条受损等情况后,选择60 °为文冠果的最佳拉枝角度。
拉枝;碳氮;文冠果;花芽分化
在全球能源危机的背景下,各国开始寻找新能源,而来源广泛、应用方便、少污染的生物能源受到广泛关注[1-3]。全球的能源发展趋势也逐步由多元化结构向可再生能源转化,逐步替代化石燃料[4]。生物质能源是重要的可再生能源,是继化石能源煤、石油和天然气之后的第4位能源[5]。生物质能极有可能成为未来可持续能源系统的组成部分[6]。
文冠果Xanthoceras sorbifolia Bunge属无患子科,文冠果属,原产我国,主要分布在辽宁、内蒙古、山西、陕西、甘肃一带[7-8]。具有可栽培的面积大、抗旱性强、含油量高、果实采收容易等特点,被国家林业局列为中国7个重点生物质能源发展树种,是我国北方地区主要的生物质能源树种。此外,文冠果具有荒山绿化、水土保持、防风固沙和观赏等诸多生态功能[9-10]。
由于文冠果存在成花而不成果的现象,使得果实产量非常低,严重阻碍了它的应用和发展。有研究表明,果树的花芽分化既与营养水平有关,也受激素的控制[14]。碳水化合物的积累有利于花芽分化,是花芽分化的结果物质和能量物质[15-17]。可溶性糖可以为花芽分化提供营养,在花芽分化中可以直接被利用,淀粉是能量物质,虽然其本身没有活性,但是花芽分化过程中可以分解成糖,供花芽分化利用[18-20]。氮是植物体内蛋白质、酶类和核酸的组成部分,氮素尤其是花和花序生长发育必需的,是根系从土壤中吸收最多的矿质元素之一[23]。而拉枝会影响碳水化合物和氮的分配,进而影响花芽分化水平。修剪和拉枝是调节树体枝组结构及促进果树生殖生长的主要措施。有学者认为,对于促进成花结果,拉枝比修剪更为有效[11-12]。修剪研究十分丰富的苹果已经被证明,拉枝是促进成花结果最为有效的手段[13]。但是,国内外对文冠果整形修剪方面的研究较少,采用疏枝或回缩,剪去下垂枝、病虫枝等结论都较为笼统[21-23]。鉴于国内外对文冠果修剪的研究主要集中在修剪反应的生物学特征上,从机理上对文冠果整形修剪效果的研究目前还没有。因此,本研究企图通过对文冠果进行不同角度的拉枝处理,研究叶片的可溶性糖、淀粉、氮的含量以及碳氮比的动态变化及其与花芽分化的关系,对拉枝的作用进行量化研究,为文冠果整形修剪技术提供理论依据。
1 材料与方法
1.1 试验地概况及试验材料
试验地位于我国文冠果栽培和研究历史最悠久的内蒙古自治区赤峰市的阿鲁科尔沁旗(43°21 ′43 ″~ 45°24′20″N,119°02′15″~ 121°01′E)。该地区年均气温5.5 ℃,年日照时数2 760~3 030 h,年日照率 68%,极端最高气温40.6 ℃,极端最低气温-32.7 ℃,年平均积温2 900~3 400 ℃,是典型的大陆型气候。年均降水量300~400 mm,夏季降水集中,湿润度为0.32,冬春以西风、西北风为主,夏秋以南风、东南风为主,年可利用的风能176 d。土壤有机质平均含量为2.9%,无霜期95~ 140 d。
试验对象为2010年实生苗栽植的文冠果树,整地方式为水平带状,林木采取均匀栽植模式,林下有稀疏草本植物,株距为2 m×2 m,2014年3月林分平均胸径9.6 cm,平均树高1.61 m,平均冠幅1.59 m(南北方向)。
1.2 试验方法
2014年3月(文冠果休眠期),选取生长势、负载量一致的文冠果树30株,将长势、生长方向、角度(30°)一致的一年生枝条侧拉成45°、60°、90°,为一组,不拉枝为对照,即对照为30°。每株树均处理8组。拉枝3个月后,文冠果叶片完全展开,分别于6月15日、7月1日、7月15日、8月1日、8月15日、9月1日、9月15日、10月1日采样,共8次。每处理每个角度每次采枝条顶部2~3个叶片,放入冰盒带回实验室用去离子水冲洗3次。冲洗干净后,105 ℃杀青30 min,80 ℃烘干至恒质量,待测定可溶性糖、淀粉和氮含量。
1.3 项目测定
可溶性含量糖采用蒽酮比色法测定;淀粉含量采用高氯酸水解后,蒽酮比色法测定[24];氮含量采用凯氏定氮仪测定[25]。
1.4 数据分析
采用spss18.0软件对氮含量、可溶性糖含量、淀粉含量、碳氮比等数据进行One-way ANOVA分析,主要影响因子设为:拉枝角度(4个水平)。因素不同水平间差异显著性采用Duncan检验(检验水平为p<0.05)。分析前,数据经Levene检验满足方差齐性等方差分析的前提条件。采用Excel 2010 软件进行图表绘制。
2 结果与分析
2.1 不同拉枝角度氮含量变化
由图1可知,总体来看,随着时间进展,各拉枝角度的枝条上的叶片氮含量呈递减的趋势。6月15日~8月1日,各拉枝角度枝条上叶片的氮含量下降急速,而在8月1日以后下降较为平缓,10月1日,拉枝60°枝条叶片氮含量最低,为0.91 g/kg,其他日期各拉枝角度枝条上叶片的氮含量均是30°> 45°> 60°> 90°。6月份 60°和 90°拉枝枝条的叶片氮含量均亦显著低于30°枝条叶片(p<0.05),90°拉枝枝条含量最低,为2.30 kg/g,但60°和90°之间没有显著差异(p>0.05)。7月份,同6月份,也是拉枝90°枝条叶片氮含量最低,分别为1.92、1.42 kg/g,与60°无显著差异,但是均极显著小于30°和45°枝条叶片氮含量(p<0.01)。8月份,拉枝90°枝条叶片氮含量最低,为1.21、1.11 g/kg,与60°枝条无显著差异(p>0.05),但是均显著低于30°枝条叶片氮含量(p<0.05)。9月份和10月份,4个拉枝处理的枝条叶片氮含量之间均没有显著差异(p>0.05)。总之,随着拉枝角度的增大,叶片氮含量逐渐减少,60°和90°之间均无显著差异(p>0.05)。
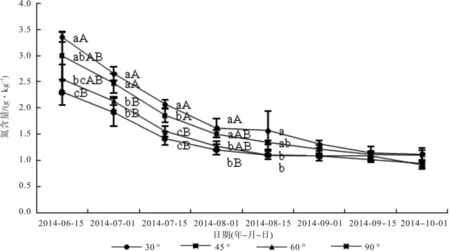
图1 文冠果叶片氮含量变化Fig. 1 Change of nitrogen content in Xanthoceras sorbifolia Bunge leaves
2.2 不同拉枝角度可溶性糖含量变化
由图2可知,随时间的推移,不同拉枝角度可溶性糖含量变化呈现出两方面的规律,一方面,各个拉枝角度枝条上的叶片可溶性糖含量变化趋势一致,均呈现递减趋势,6月15日—7月15日各拉枝角度的叶片可溶性糖含量下降急速,而在7月15日以后,各拉枝角度的叶片可溶性糖含量下降平缓。另一方面,6月15日、7月1日、7月15日、8月1日、9月1日60°枝条叶片可溶性糖含量最高,分别为19.35%、16.00%、13.15%、13.05%、9.73%; 9月15日、10月1日90°枝条叶片可溶性糖含量最高,达到9.57%、9.04%。拉枝60°枝条与90°枝条可溶性糖含量无显著差异(p>0.05),并极显著高于30°和45°(p<0.01)。因此说,随着拉枝角度的增大,叶片可溶性糖含量逐渐增大,但是60°和90°之间无显著差异(p>0.05)。
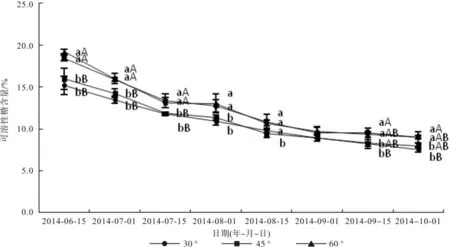
图2 文冠果叶片可溶性糖含量变化Fig. 2 Change of soluble sugar content in Xanthoceras sorbifolia Bunge leaves
2.3 不同拉枝角度淀粉含量的动态变化
由图3可知,不同拉枝角度叶片上的淀粉含量在6月15日—7月15日呈递增趋势,7月15日以后呈递减趋势,并且7月15日—8月15日下降急速,而8月15日以后下降较为平缓。6月份和7月份,拉枝90°枝条叶片淀粉含量最高,达到2.47%(6月15日)、2.74%(7月1日)、3.26%(7月15日),和60°枝条无显著差异(p>0.05),并均显著高于30°和45°枝条叶片淀粉含量(p<0.05)。8月份,30°枝条叶片淀粉含量最低,为1.45%、1.32%,显著低于60°和90°枝条叶片淀粉含量(p<0.05)。9月份和10月份,各拉枝处理枝条叶片淀粉含量之间均无显著差异(p>0.05)。总之,随着拉枝角度的增大,叶片淀粉含量逐渐增大,但是60°和90°之间没有显著差异(p>0.05)。
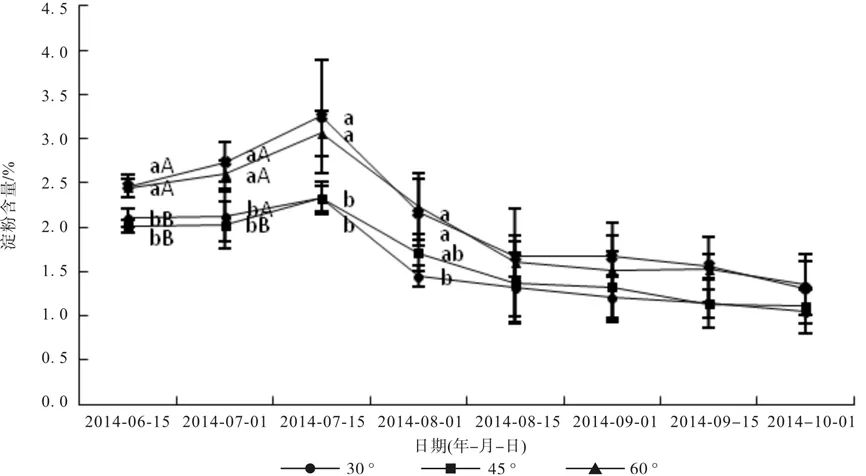
图3 文冠果叶片淀粉含量变化Fig. 3 Change of starch content in Xanthoceras sorbifolia Bunge leaves
2.4 各拉枝角度碳氮比的动态变化
由图4可知,8月1日之前,各拉枝角度的枝条上碳氮比呈上升趋势,而8月1日之后,呈平缓下降的趋势,9月1日和10月1日,拉枝60°的枝条叶片的碳氮比最高,达到8.87、9.91,其余日期均,拉枝90°的枝条叶片碳氮比最高。6月份、7月份和8月份,拉枝90°枝条叶片碳氮比最大,达到8.08、8.38、9.41、10.60、9.80,与 60°没有显著差异(p>0.05),7月15日取样除外(90°显著高于60°),并均极显著高于30°和45°枝条叶片碳氮比(p<0.01)。9月份和10月份,拉枝60°和90°之间叶片碳氮比均无显著差异(p>0.05),但均显著低于30°和45°枝条叶片碳氮比(p<0.01)。总之,随着拉枝角度的增大,叶片碳氮比逐渐增大,并且除了一次取样之外,60°和90°枝条叶片碳氮比均无显著差异(p>0.05)。
3 结论与讨论
目前我国文冠果由于疏于管理,大多为顶端结果,侧芽几乎不能结果,顶端优势非常明显,“大小年”现象也非常突出,因此提高花芽分化的质量具有重要的意义。为了获得稳产,控制树体生长过旺,消除枝条顶端优势是一种常见和重要的技术措施。而通过拉枝处理,可以延缓树势,改善树体营养分配[26]。
3.1 不同拉枝角度对可溶性糖和淀粉含量的影响
不同拉枝角度处理后,拉枝角度越小,顶端优势越明显,大多数时期都是60°和90°枝条的叶片可溶性含量和淀粉含量显著大于30°和45°,说明拉枝角度增大,有利于碳素的积累,为花芽分化提供营养。这可能是因为开张角度增大后削弱了顶端优势,从而削弱了枝条的营养生长,减少了光合产物的外运,开张角度较小的枝条更易形成旺长,代谢较旺,呼吸作用增强,有机物消耗大,碳素的积累就低。也有可能是大角度损伤了枝条基部,影响了光合产物外运,增加了叶片自身的碳素积累。这与其他学者的研究结果是一致的[20,27]。
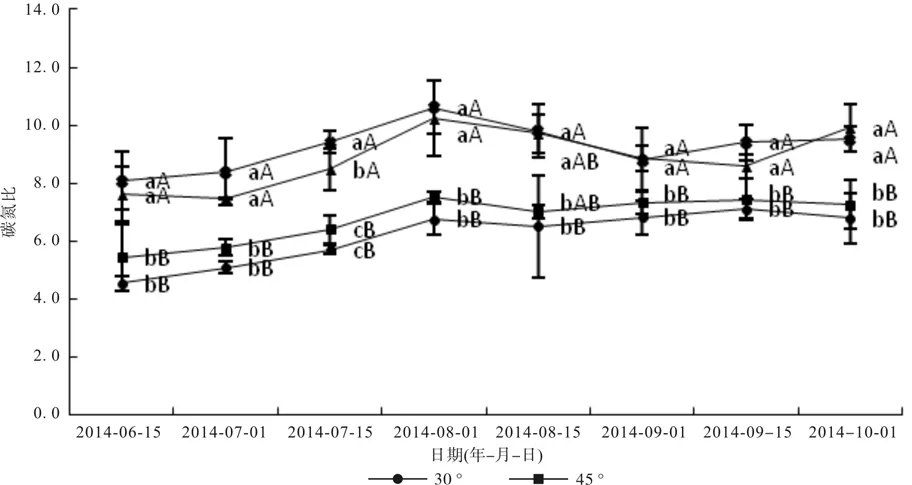
图4 文冠果叶片碳氮比变化Fig. 4 Change of C/N in Xanthoceras sorbifolia Bunge leaves
3.2 不同拉枝角度对氮含量的影响
氮素运输与碳素不同,由根系吸收向上运输,并且氮素的分配方向受生长中心和蒸腾作用的影响[28-30]。叶片中氮含量的高低更能反映植株的真实营养状况[31]。不同拉枝角度处理后,大多数时期都是30°和45°枝条叶片的氮含量显著大于60°和90°,说明拉枝角度的增大不利于氮素的积累。这可能是因为小的开张角度的枝条由于生长旺盛,形成新的生长中心,蒸腾作用强,氮素在竞争上处于优势,氮素得到积累。大开张角度的枝条由于生长不旺,蒸腾作用弱,氮素在竞争上处于劣势,而且,枝条开张角度增大后,极性生长削弱,地上部分碳水化合物向根系运输减少,氮素向地上部分运输量也减少,相互减弱可以使树势缓和[32]。本试验结论与之前其他学者研究结果相似[33-36]。
3.3 不同拉枝角度对碳氮比的影响
单独的碳素含量和氮素含量都不是决定花芽分化的决定因素,碳氮比才是影响花芽分化的关键[37]。大多数时期60°和90°碳氮比都显著大于30°和45°,说明拉枝角度的增大有利于碳氮比的增大。
3.4 实际应用
本试验证明,拉枝角度越大,叶片可溶性糖含量、淀粉含量和碳氮比都有所增加,氮含量有所减少,但是60°和90°之间并无显著差异。通过实际试验发现,文冠果枝条在拉枝90°时,木质部容易受损,枝条也易老化,不利于长期发展。鉴于以上结论,可确定文冠果的最佳拉枝角度是60°,这样的角度可较好地改善光照通风条件,创造多碳少氮的环境,也不会造成树体早衰,更加有利于促进花芽形成,为实现优质高产。
[1] 纪占武, 郑文范. 关于发展生物能源化解能源危机的思考[J].东北大学学报: 社会科学版, 2009, 11(6): 490-495.
[2] 范辉华, 张天宇, 姚湘明, 等. 高产油脂无患子优树的选择[J].中南林业科技大学学报, 2014, 34(5): 4-8.
[3] 郭伦发, 王新桂, 江新能, 等. 广西主要木本油料植物生态经济综合评价[J]. 经济林研究, 2014, 32(3): 81-86.
[4] Klepeis N E, Nelson W C, Ott W R, et al. The National Human Activity Pattern Survey (NHAPS): a resource for assessing exposure to environmental pollutants[J]. Journal of exposure analysis and environmental epidemiology, 2001, 11(3):231-252.
[5] Li H, Zhang Z, Lin S, et al. Research advances in the study of Pistacia chinensis Bunge, a superior tree species for biomass energy[J]. Forestry Studies in China, 2007, 9(2): 164-168.
[6] Hoogwijk M, Faaij A, van den Broek R, et al. Exploration of the ranges of the global potential of biomass for energy[J]. Biomass and bioenergy, 2003, 25(2): 119-133.
[7] 黄维志.荒山绿化的好树种—文冠果[J]。甘肃林业,2004(1): 35-35.
[8] 杨东霞. 文冠果产业开发初探[J]. 内蒙古林业, 2012(1):34-34.
[9] 康永祥, 康博文, 刘建军, 等. 陕北黄土高原文冠果群落结构及物种多样性[J]. 生态学报, 2010 (16):4328-4339.
[10] 范明浩.北方优良生物质能源树种文冠果[J]. 内蒙古林业,2007(12): 019.
[11] Goldschmidt-Reischel E. Regulating trees of apple and pear by pruning and bending[J]. Swedish Journal of Agricultural Research (Sweden), 1997.
[12] Goldschmidt-Reischel E. Regulating trees of apple and pear by pruning and bending[J]. Swedish Journal of Agricultural Research (Sweden), 1997.
[13] Costes E, Lauri P E, Regnard J L. Analyzing fruit tree architecture: implications for tree management and fruit production[J]. Horticultural Reviews, 2006(32): 1-61.
[14] 陈 清, 周 纯, 周学明. 苹果花芽分化的激素调节机理研究及控制技术应用效果[J]. 山西果树, 2006 (3): 38-39.
[15] Faust M. Physiology of temperate zone fruit trees[M]. John Wiley & Sons, Inc., 1989.
[16] Greenberg J, Goldschmidt E E, Goren R. Potential and limitations of the use of paclobutrazol in citrus orchards in Israel[C]//VII International Symposium on Plant Growth Regulators in Fruit Production 329. 1992: 58-61.
[17] 余拱鑫. 拉枝角度对苹果短枝顶芽碳氮和内源激素含量的影响[D]. 西北农林科技大学, 2012.
[18] 梁朗玛, 曾 辉, 杜丽清, 等. HPLC 测定澳洲坚果枝叶的糖类[J]. 广西热带农业,2007 (2): 4-6.
[19] Menzel C M, Carseldine M L, Simpson D R. effect of fruiting status on nutrient composition of litchi (Litchi chinensis Sonn.) during the flowering and fruiting season[J]. Journal of horticultural science, 1988.
[20] 孙乃波, 张志宏. 草莓花芽分化过程中叶片碳水化合物和蛋白质含量的变化[J]. 安徽农业科学, 2006, 34(11): 2328-2329.
[21] 朝克吐. 浅谈文冠果整形修剪技术要点[J]. 内蒙古林业调查设计, 2013 (5): 48-48.
[22] 慎学根, 张德辉. 文冠果增产技术的研究[J]. 中国科技博览,2009 (09X): 103-103.
[23] 蔡 龙. 文冠果栽培及修剪技术探讨[D]. 北京林业大学,2013.
[24] 高俊凤.植物生理学实验技术[M].西安:世界图书出版社,2000: 145-148.
[25] 鲍士旦.土壤理化分析(第三版)[M].北京;中国农业出版社, 2000: 264-271.
[26] 杨 勇. 拉枝角度对苹果芽, 叶 C, N 及内源激素的影响 [D].2010.
[27] 张殿高. 苹果树剪枝图解[M].沈阳:辽宁科学技术出版社,1995, 6.
[28] Bi G, Scagel C F, Fuchigami L H. Effects of spring soil nitrogen application on nitrogen remobilization, uptake, and partitioning for new growth in almond nursery plants[J]. Journal of horticultural science & biotechnology, 2004, 79(3): 431-436
[29] Moing A, Lafargue B, Lespinasse J M, et al. Carbon and nitrogen reserves in prune tree shoots: effect of training system[J].Scientia horticulturae, 1994, 57(1): 99-110.
[30] Millard P, Proe M F. Storage and internal cycling of nitrogen in relation to seasonal growth of Sitka spruce[J]. Tree physiology,1992, 10(1): 33-43.
[31] 彭福田, 姜远茂. 不同产量水平苹果园氮磷钾营养特点研究[J]. 中国农业科学, 2006, 39(2): 361-367.
[32] 杨洪强,束怀瑞. 苹果根系研究[M].北京:科学科学出版社,2007:52-53.
[33] Han H H, Coutand C, Cochard H, et al. Effects of shoot bending on lateral fate and hydraulics: invariant and changing traits across five apple genotypes[J]. Journal of experimental botany,2007, 58(13): 3537-3547.
[34] Scandellari F, Tonon G, Thalheimer M, et al. Assessing nitrogen fluxes from roots to soil associated to rhizodeposition by apple(Malus domestica) trees[J]. Trees, 2007, 21(5): 499-505.
[35] Glover J D, Reganold J P, Andrews P K. Systematic method for rating soil quality of conventional, organic, and integrated apple orchards in Washington State[J]. Agriculture, ecosystems &environment, 2000, 80(1): 29-45.
[36] 李美美, 马华冰, 宋新英, 等. 不同修剪措施对成龄‘绿岭’核桃抗腐烂病能力及树势的影响[J]. 经济林研究, 2014,32(3): 61-67.
[37] 余拱鑫, 韩明玉, 张满让, 等. 不同拉枝角度对 ‘嘎啦’苹果顶芽及叶片碳氮含量的影响[J]. 北方园艺, 2012(15):9-12.
Response mechanism of human disturbance to the content of carbon and nitrogen in the leaves of Xanthoceras sorbifolia Bunge
SU Man-lin1,2, MA Lv-yi1,2, DUAN Jie2, WU Shang1,2, SU Su-chai1,2, AO Yan1,2
(1. Key Laboratory of Silviculture and Conversation, Ministry of Education, Beijing Forestry University, Beijing 100083, China;2. National Energy R&D Center for Non-food Biomass, Beijing 100083, China)
There is close relationship between the content of carbon, nitrogen and flower bud differentiation, the angle of branch influences the content of C,N in leaves and then has effect on lower bud differentiation. The effects of bending branch(30°,45°,60°,90°)on the changes of the content of soluble sugar, starch, the nitrogen in leaves of Xanthoceras sorbifolia Bunge were studied. The result showed that the contents of soluble sugar, starch and C/N of branches angle of 60°and 90°were significantly higher than those of 30°and 45°(p<0.05),but the contents of nitrogen were the opposite. There was no significant difference between 60°and90°,30°and45°(p >0.05). Considering the circumstance of flowering and fruiting and branches damaged, 60° is the best angle for Xanthoceras sorbifolia Bunge.
branch bending; carbon and nitrogen; Xanthoceras sorbifolia Bunge; bud differentiation
S759.3+3
A
1673-923X(2016)08-0027-06
10.14067/j.cnki.1673-923x.2016.08.006
2015-10-22
国家国际科技合作专项项目“高能效先进生物质原料林可持续经营技术合作研究”(2014DFA31140)
苏曼琳,硕士研究生
马履一,教授;E-mail:maluyi@bjfu.edu.cn
苏曼琳,马履一,段 劼,等. 文冠果叶碳氮含量对人为拉枝干扰的响应机制 [J].中南林业科技大学学报,2016, 36(8):27-32.
[本文编校:文凤鸣]
