脉冲降雨对土壤异养呼吸影响机制的模拟研究
2016-12-14李一强王义东王辉民王中良
李一强,王义东,王辉民,王中良
(1.天津师范大学a.天津市水资源与水环境重点实验室,b.城市与环境科学学院,天津 300387;2.中国科学院地理科学与资源研究所,北京 100101)
脉冲降雨对土壤异养呼吸影响机制的模拟研究
李一强1,王义东1,王辉民2,王中良1
(1.天津师范大学a.天津市水资源与水环境重点实验室,b.城市与环境科学学院,天津 300387;2.中国科学院地理科学与资源研究所,北京 100101)
为研究脉冲降雨对土壤异养呼吸的影响机制,通过设置土壤+凋落物+降雨(A)、土壤+降雨(B)、土壤+凋落物+灭菌+降雨(C)、土壤+灭菌+降雨(D)和土壤+凋落物+无降雨(CK)共5组处理,采用室内培养法分析了脉冲降雨对中亚热带湿地松林下凋落物土壤异养呼吸的影响.结果表明:模拟降雨不同程度地激发了凋落物呼吸和矿质土壤呼吸,两者在0.25~1 h达到峰值,后逐渐恢复到降雨前水平,模拟降雨引起的物理排气过程持续时间较短.凋落物呼吸对降雨激发的土壤异养呼吸的平均贡献率为93.1%,而矿质土壤呼吸仅为6.9%.由此可知,降雨不同程度地激发了凋落物呼吸和矿质土壤呼吸,物理排气过程对土壤呼吸贡献有限,而凋落物呼吸对激发土壤异养呼吸贡献很大,且作用时间较长(48 h),是主导土壤异养呼吸对降雨响应的关键.
土壤CO2排放;土壤呼吸;凋落物分解;模拟降雨
近年来,人类的工业化活动导致大气中温室气体浓度不断增加,由此引起的全球性气候变暖和其他环境变化受到广泛关注[1].CO2是温室效应最大的贡献者,对全球变暖的贡献率达到60%以上[2].全球的森林覆盖率约为陆地总面积的1/4,是全球碳库的重要组成部分,该生态系统碳贮量约为1 146 Pg(1 Pg=109t),
其中超过2/3的碳储存在土壤中[3].作为森林生态系统碳交换的重要环节,土壤呼吸约占森林生态系统总呼吸量的50%~80%[4-7],因此,土壤呼吸的微小变化就可以显著改变大气中CO2的浓度[8],进而影响气候变化.
土壤异养呼吸是指在微生物参与下土壤碳以CO2形式流向大气圈的矿化过程,主要包括矿质土壤呼吸和凋落物层呼吸[9].土壤异养呼吸是一个较为复杂的过程,受土壤微生物群落组成及活性、土壤温度和湿度、植被类型、土壤养分供给条件等多种因素影响[10].在受到干旱胁迫时,微生物的数量会减少,个体呼吸速率也会下降,对土壤异养呼吸速率的贡献降低.近年来的研究表明,降雨可改善土壤水分状况,加快土壤CO2的产生,从而提高土壤异养呼吸速率,如Gordon等[11]研究结果表明土壤水分改善后微生物的活性增强,Gestel等[12]报道了不同气候区的土壤在水分条件改善后生物量增加.凋落物呼吸是土壤异养呼吸的重要组成部分[13],在全球森林系统中,凋落物对土壤呼吸总量的贡献率约为33%,且土壤呼吸速率在一定范围内会随着凋落物的增加而升高[14].此外,凋落物分解可以改变土壤的养分条件并影响碳循环[15],但地表凋落物蒸发越强烈,持水能力越弱,其分解越容易受到水分不足的胁迫[16-17].降雨可以迅速改变地表凋落物的水分状况,水分增加促使凋落物分解速率加快,继而使土壤呼吸速率增大[18].同时,凋落物分解的加快又为微生物提供了大量的营养物质和相对合适的生存环境,使得物理排气过程基本结束后土壤呼吸速率在雨后短时间内快速升高[19-20].在大多数土壤异养呼吸与水分关系的研究中,凋落物呼吸与矿质土壤呼吸对土壤异养呼吸的贡献并没有分开讨论,而是笼统地归因于土壤的干湿程度[20-22],凋落物呼吸对降雨的响应过程及对土壤异养呼吸的影响还有待探讨.本研究选取具有典型代表性的中亚热带湿地松林,通过设置5组不同处理,采用室内培养法探讨脉冲降雨对土壤异养呼吸的影响机制.
1 材料与方法
1.1 研究区概况
湿地松原产美国东南部,我国引种后广泛种植于长江以南大部分地区,成为营林造林的先锋物种[23].本研究选取亚热带常绿湿地松人工林作为研究区域(26°44′34″N~26°44′43″N,115°03′35″E~115°03′40″E),该人工林位于江西省泰和县中国科学院千烟洲生态试验站内,地貌类型为红壤丘陵地貌,海拔60~115 m.地处中亚热带湿润季风气候区,年平均气温为17.9℃,全年无霜期共290 d,降雨量为1 469 mm,降雨主要集中在3~6月[24].由于长期过度砍伐,该区域内典型地带性亚热带常绿阔叶林基本消失.目前,现有林地多为1985年前后营造的人工林,主要树种为湿地松(Pinuselliottii)、柑橘(CitrusreticulateBlanco)和马尾松(Pinusmassoniana Lamb)等[25].
1.2 实验设计与样品采集
根据相关资料,并结合实际情况,本研究设置土壤+凋落物+降雨(A组)、土壤+降雨(B组)、土壤+凋落物+灭菌+降雨(C组)、土壤+灭菌+降雨(D组)和土壤+凋落物+无降雨(CK组)共5组处理,每组处理设3个重复.取湿地松林下表层0~20 cm土壤,土样在实验室内自然风干,剔除石块和植物残体等杂质,碾磨,过2 mm筛,混匀,密封保存以备使用.使用Vario EL III型元素分析仪测定得到土壤有机质质量分数为1.1%,全氮质量分数为0.1%.凋落物收集采用凋落物收集框法,即在林地内及周围安置3个凋落物收集框(1 m×1 m),每月收集一次,将收集的凋落物分为叶、枝、花和果实,充分混匀,在实验室自然风干,密封备用.
1.3 实验过程与数据测定
选取15个高度约为5 cm、直径为10.6 cm的土壤呼吸环,用AB胶水和塑料板将环底部密封住.称取200 g土样,装入环中,轻压土体,使土体高度约2 cm.依据当时的土壤含水率,加约25 g水,使土壤体积含水率保持在15%,放在培养箱内,温度控制在25℃,培养7d,此间维持水分含量在15%.取出需要做灭菌处理的2组(C组和D组),共6个土壤呼吸环,用氯仿(CHCl3)熏蒸灭菌2 d,具体操作为将已装好土的土壤呼吸环放入3个大的干燥器中,将装有30 mL无水氯仿的小烧杯(带有沸石)、装有30 mL NaOH的小烧杯和少许水一并置于干燥器内,密闭抽气直至氯仿沸腾3 min,然后放置于装有空调的小实验室(温度控制在25℃)内,在黑暗中静置2 d.随后取出装有氯仿和NaOH的烧杯,再用真空压缩泵将干燥器内的空气抽至无味,取出土壤呼吸环,静置2~3 h后测定土壤呼吸本底值以检验灭菌效果.在模拟降雨前,取干重20 g的凋落物覆在3组(A组、C组和CK组)共9个土壤呼吸环的土面上,模拟地表凋落物层,另外2组(B组和D组)6个土壤呼吸环则裸露在地表,以此研究凋落物对降雨的响应.对于模拟降雨处理,分别加蒸馏水10 g,并在模拟降雨的-72 h(灭
菌处理前)、-2h(灭菌后)、0 min(开始模拟降雨)、15 min、30 min、1 h、2 h、4 h、8 h、16 h、24 h、36 h和48 h利用LI-8100型土壤碳通量自动测量仪测定土壤呼吸速率,同时测定未加水处理的土壤呼吸(CK组).每个时间点测定2次,2次测定的时间间隔为1 min;每次测定总时间为2 min,前30 s舍去,不进行回归计算,仪器自动取2次测定的平均值,作为计算平均呼吸速率的3个重复之一.整个实验操作均在该实验室内进行,室内温度始终控制在25℃.
1.4 数据分析
实验过程中,凋落物呼吸、物理排气和矿质土壤呼吸对土壤呼吸激发部分的瞬时贡献率(%)分别为
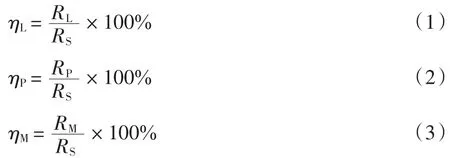
式(1)~式(3)中:ηL为凋落物呼吸对土壤呼吸激发部分的瞬时贡献率(%);ηP为物理排气对土壤呼吸激发部分的瞬时贡献率(%);ηM为矿质土壤呼吸对土壤呼吸激发部分的瞬时贡献率(%);各组分瞬时呼吸速率RL、RS、RP和RM分别为[26]:
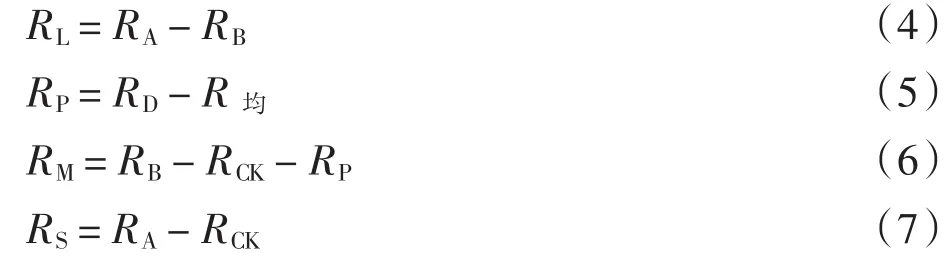
式(4)~式(7)中:RL为凋落物呼吸产生的平均呼吸速率(μmol·(m-2·s-1));RA为A组平均呼吸速率(μmol·(m-2·s-1));RB为B组平均呼吸速率(μmol·m-2·s-1);RP为平均物理排气速率(μmol·m-2·s-1);RD为D组平均呼吸速率(μmol·m-2·s-1);R均为D组8~48 h的平均呼吸速率(μmol·m-2·s-1);RM为降雨激发的矿质土壤呼吸速率(μmol·m-2·s-1);RCK为CK组平均呼吸速率(μmol·m-2·s-1);RS为降雨激发的平均呼吸速率(μmol·m-2·s-1).
此外,凋落物呼吸和降雨激发矿质土壤呼吸对土壤异养呼吸的贡献率(%)为

式(8)和式(9)中:λL为凋落物呼吸对土壤异养呼吸的贡献率(%);λM为降雨激发矿质土壤呼吸对土壤异

式(10)~式(13)中:TL为凋落物呼吸产生的呼吸累积量(mmol·m-2);TA为A组呼吸累积量(mmol·m-2);TB为B组呼吸累积量(mmol·m-2);TP为物理排气累积量(mmol·m-2);TD为D组呼吸累积量(mmol·m-2);T均为D组8~48 h的平均呼吸累积量(mmol·m-2);TCK为CK组呼吸累积量(mmol·m-2);TM为降雨激发的矿质土壤呼吸累积量(mmol·m-2);TH为土壤异养呼吸累积总量(mmol·m-2).
使用 Origin9.0软件中“Mathematics” 下的“integrate”程序对各组土壤呼吸速率的均值进行“Mathmatical Area”的面积积分,得到5组处理不同阶段的土壤呼吸累积量.养呼吸的贡献率(%);TL、TM和TH为
2 结果与分析
2.1 土壤异养呼吸对降雨的响应
实验测定所得各组土壤呼吸速率随时间的变化关系如图1所示,其中图1(b)为图1(a)中虚线部分的局部放大图.
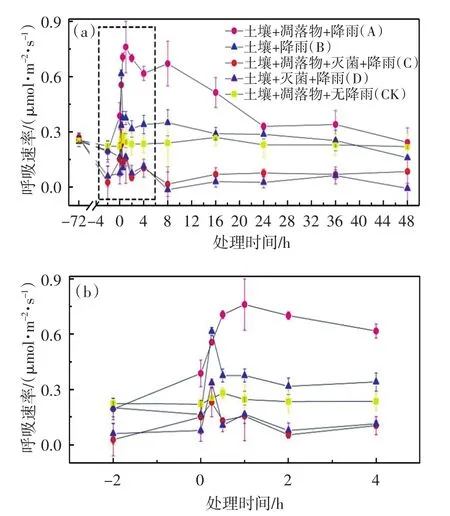
图1 土壤呼吸速率瞬时变化图Fig.1 Instantaneous variation of soil respiration rate
由图1可以看出,在进行灭菌处理前(-72 h),各组样品的土壤呼吸速率没有显著差异(0.25~0.27 μmol/
(m2·s)).灭菌处理结束后(-2 h),进行灭菌的C组(土壤+凋落物+灭菌+降雨)和D组(土壤+灭菌+降雨)的土壤呼吸速率与未经灭菌的A(土壤+凋落物+降雨)、B(土壤+降雨)和CK(土壤+凋落物+无降雨)3组的土壤呼吸速率产生明显区别,其中C组和D组的平均呼吸速率下降明显,而A、B和CK组仅出现少许下降.微生物呼吸是土壤异养呼吸的重要组成部分,灭菌处理杀死微生物后便不再有CO2释放,但含碳矿物质的化学氧化作用在灭菌处理后依然会进行[9],因此C组和D组的平均呼吸速率尽管出现大幅度下降,分别下降至0.03 μmol/(m2·s)和0.06 μmol/(m2·s),但并没有直接下降为0.微生物群落的营养物质主要来源于土壤中的有机质,活性有机质的减少会限制微生物的活动,减少CO2的产生.因此,在-72~-2 h这段时间内,A、B组及CK组的土壤呼吸速率也出现了少许下降,但数值仍很接近(0.19~0.22 μmol/(m2·s)).CK组虽并未进行模拟降雨,但凋落物分解与土壤有机质的胁迫可能产生了相互抵消,其平均呼吸速率在0 min~48 h时间段内基本没有变化,为后续比较提供参考.
开始模拟降雨(0 min)后,雨水与土壤孔隙的物理替代作用使土壤孔隙中的CO2最先排出,使土壤呼吸量增加[19].因此,受物理排气过程的影响,A、B、C和D组的土壤呼吸速率在0~15 min时间段内均明显增加,且15 min时,B、C和D组的平均呼吸速率均达到最大值,分别为0.62、0.23和0.34 μmol/(m2·s).
凋落物分解是土壤有机质的重要来源,也是土壤异养呼吸的重要组成部分[27].蒸发强烈与持水能力弱造成凋落物分解更容易受到水分不足的胁迫,而与温度相比,凋落物分解更依赖于水分的供给[16-17],雨后凋落物的分解可以显著提高土壤呼吸速率[19,28].因此,A组的平均呼吸速率并没有在15 min时达到最大值,而是继续升高,在1 h时达到0.79 μmol/(m2·s)的最大值.达到最大值后,受到水分降低的胁迫,A组的平均呼吸速率持续缓慢下降,并在48 h后与CK基本相同,分别为0.24和0.22 μmol/(m2·s).
降雨在一定范围内可以增加微生物的生物量[12,29],增强土壤微生物活性[11,30],水分条件改善后微生物可迅速利用底物进行新陈代谢[31],促使CO2释放加快,因此B组的土壤呼吸速率在15 min后出现大幅下降,但30 min时仍高于模拟降雨前(-72 h~0 min)自身的平均呼吸速率以及同一时间点CK组的平均呼吸速率,且此状态一直持续了相当长的时间.约40 h后,B组的土壤呼吸速率逐渐低于有凋落物覆盖的CK组,同时也低于模拟降雨前(-72 h~0 min)自身的平均呼吸速率,这很有可能是因为受到营养物质供给的胁迫.Wu等[32]利用不同类型的森林土壤在不同湿度下进行模拟实验,表明随着土壤中所添加营养物质的不断消耗,不同处理条件下土壤异养呼吸速率均不断降低,为本研究上述结果提供支持.
灭菌后,微生物不再产生CO2,与微生物呼吸相比,含碳矿物质的化学氧化作用比较微弱[9].因此,C组和D组的平均呼吸速率在15 min~8 h时间段内的变化与物理排气过程息息相关.15 min后,2组的平均呼吸速率出现大幅下降但并不为0;8 h后,物理排气过程基本结束,2组的平均呼吸速率也降至模拟降雨(0 min)后的最低点;由于氯仿熏蒸灭菌不彻底,因此C组和D组的平均呼吸速率在8 h后又逐渐升高.凋落物在雨后分解可为微生物生长繁殖提供丰富的营养物质[20],因此C组微生物的生存环境优于D组,造成C组的平均呼吸速率逐渐高于D组.尽管C组和D组的平均呼吸速率在8~48 h时间段内有了一定的升高,但始终远低于同一时间点未灭菌的A组和B组.
凋落物呼吸、物理排气和矿质土壤呼吸对土壤呼吸激发部分的瞬时贡献率如图2所示.
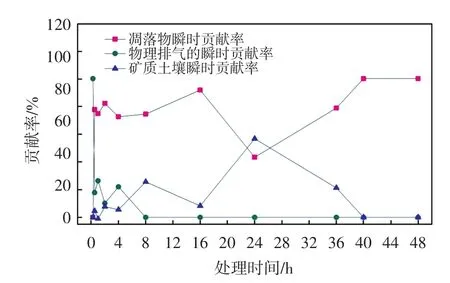
图2 土壤呼吸瞬时贡献率图Fig.2 Instantaneous contribution rate of soil respiration
由图2可以看出,整个模拟过程中(0~48 h),物理排气的瞬时贡献率迅速降低,在8 h后几乎为0;凋落物瞬时贡献率则迅速上升至约80%,从16 h后开始逐渐下降,在24 h时降至最低点(43.3%);矿质土壤呼吸对降雨的响应较慢,在1 h后出现显著变化,这与Xu等[33]在野外观测到微生物对降雨的响应大约需要1 h的研究结果基本一致,此后,矿质土壤呼吸对土壤呼吸激发部分的瞬时贡献率逐步升高,在24 h时升至最高点(56.7%).结合图1中信息可知,在24 h时,凋落物分解释放的CO2量下降明显,从而降低了
凋落物呼吸对土壤呼吸激发部分的瞬时贡献率,并相对地提高了矿质土壤呼吸对土壤呼吸激发部分的瞬时贡献率.24 h后,受土壤有机质胁迫,激发部分的矿质土壤呼吸下降明显,降低了对土壤呼吸激发部分的瞬时贡献率,并在40 h时下降为0,而此时凋落物呼吸下降放缓,对土壤呼吸激发部分的瞬时贡献率逐渐升高至100%.
2.2 降雨对土壤异养呼吸的影响
各组土壤呼吸累积量随时间的变化关系如图3所示.
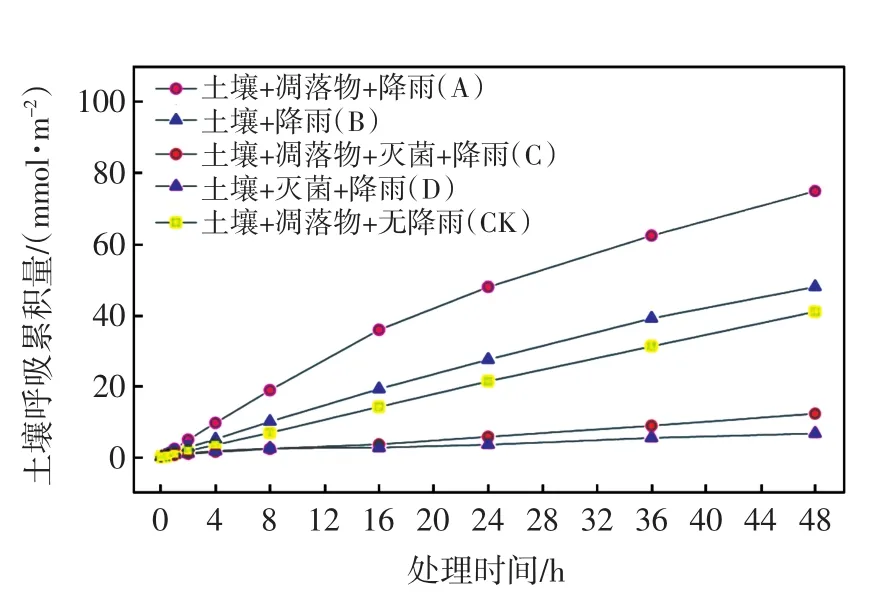
图3 土壤呼吸累积变化Fig.3 Cumulative variation of soil respiration
由图3可以看出,在有凋落物覆盖的条件下,受降雨激发的A组呼吸累积量(75.20 mmol/m2)远大于CK组(41.09 mmol/m2),为CK组的1.83倍;同时降雨也激发了B组(48.16 mmol/m2)的矿质土壤呼吸,但其呼吸累积量远小于A组,仅为CK组的1.17倍,这表明在无凋落物覆盖的情况下,降雨对矿质土壤呼吸的激发作用有限.进行灭菌处理的C组和D组的呼吸累积量较小,远小于同期A组和B组的呼吸累积量.此外,营养物质供给条件的不同造成C组和D组微生物群落恢复速度不同,8 h后,C组(12.27 mmol/m2)的呼吸累积量逐渐大于D组的呼吸累积量(6.69 mmol/ m2),尽管恢复速度较快,但短时间内仍无法恢复到原有水平.
依据各组呼吸累积量数据,雨后凋落物分解使整个过程(0 min~48 h)的土壤平均呼吸速率比无降雨条件下的土壤平均呼吸速率(CK组)提高了65.8%(不包括物理排气),雨后矿质土壤呼吸也使整个过程(0 min~48 h)的土壤平均呼吸速率比无降雨条件下的土壤平均呼吸速率提高了4.9%(不包括物理排气).上述结果的提高率(65.8%)显著高于其他研究结果,如Cisneros-Dozal等[18]指出温带落叶林凋落物在雨后的分解能够使土壤呼吸速率平均增加约24%;混合落叶林凋落物在加入水后也能使原地监测的土壤呼吸速率提高11%~26%[20];Cisneros-Dozal等[17]发现雨后凋落物呼吸对土壤呼吸的贡献率从1%上升到42%;在Ataka等[34]的研究中,凋落物在雨后的呼吸速率迅速提高,在峰值时对土壤呼吸的贡献率为51%.然而,凋落物呼吸对土壤呼吸的贡献并不完全源自自身分解释放的CO2,微生物分解土壤有机质作为土壤异养呼吸中不可忽视的部分[35],有机质的输入也可影响土壤微生物,从而影响CO2的释放[36-37].上述研究结果利用不同的方法研究凋落物呼吸对土壤异养呼吸的贡献,并未涉及土壤微生物活动与凋落物分解的互相促进作用,而本研究的高贡献率表明降雨带来的水分增加提高了凋落物的可利用性,很可能进一步激发土壤的微生物活动,从而促进了凋落物层的CO2释放.
各阶段凋落物呼吸和矿质土壤呼吸所激发的土壤异养呼吸的贡献率如图4所示.
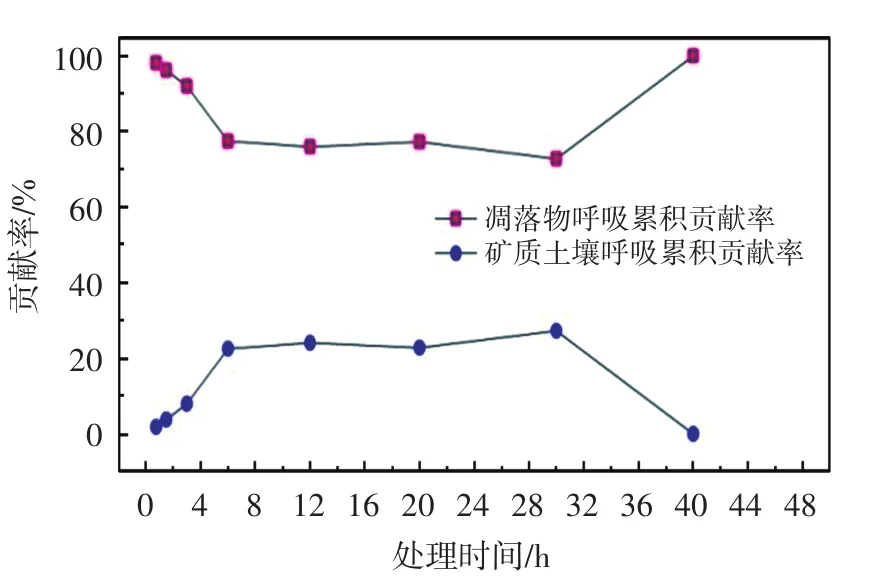
图4 土壤异养呼吸各组分贡献率Fig.4 Each component contribution rate of soil heterotrophic respiration
凋落物分解可为微生物的生长繁殖提供大量营养物质[20],从而促进CO2释放.由图4可以看出,受降雨激发影响,凋落物呼吸对土壤异养呼吸的贡献率在0~1 h内迅速提高,继而逐步下降,在6 h时下降至稳定状态并持续到30 h,之后又逐步升高.降雨同样激发了矿质土壤呼吸但响应较为缓慢,降雨激发的矿质土壤呼吸贡献率也随着时间推移而逐渐上升,在6 h时上升至稳定状态并持续到30 h,由于营养物质的胁迫,30 h后逐渐下降为0.凋落物对土壤异养呼吸的贡献率与矿质土壤呼吸的变化密切相关,矿质土壤呼吸的逐步上升相对降低了凋落物对土壤异养呼吸的贡献率,30 h后矿质土壤呼吸的逐步下降也使凋落物对土壤异养呼吸的贡献率得到提高.在整个模拟过程中,由凋落物分解引起的异养呼吸对降雨激发的土壤异养呼吸的平均贡献率为93.1%(0~48 h);矿质土壤呼
吸仅为6.9%(0~48 h).与A组相比,降雨激发的矿质土壤呼吸对土壤异养呼吸的平均贡献率仅为6.9%(0~48 h)表明无凋落物覆盖时,降雨对土壤异养呼吸的激发作用有限.
凋落物厚度是影响凋落物呼吸及其对土壤异养呼吸贡献的重要因素[38],不同厚度凋落物在雨后的响应有待进一步研究.此外,本研究中C组和D组的呼吸速率均不为0,且呼吸速率和呼吸累积量随时间不断升高,间接表明熏蒸灭菌处理可能不够彻底,即未完全杀死所有微生物,从而在一定程度上影响了本研究的结果,但并未影响本研究的主要结论.此外,室内控制实验具有一定的局限性,下一步将利用野外原位模拟实验进一步验证所得结论.
3 结论
以中亚热带湿地松林为例,通过设置土壤+凋落物+降雨(A)、土壤+降雨(B)、土壤+凋落物+灭菌+降雨(C)、土壤+灭菌+降雨(D)和土壤+凋落物+无降雨(CK)共5组处理,采用室内培养法探讨了脉冲降雨对土壤异养呼吸的影响机制,得到以下结论:
(1)凋落物呼吸对降雨响应迅速,且降雨不同程度激发了凋落物呼吸和矿质土壤呼吸,分别使土壤平均呼吸速率比无降雨条件下提高了65.8%和4.9%,激发效应持续时间较长(40~48 h).
(2)降雨显著提高了凋落物呼吸对土壤异养呼吸的贡献,由凋落物分解引起的异养呼吸对降雨激发的土壤异养呼吸平均贡献率为93.1%(0~48 h),矿质土壤呼吸仅为6.9%(0~48 h),表明凋落物呼吸是雨后土壤异养呼吸速率快速升高的重要原因.
(3)降雨引起的物理排气过程持续时间较短,且对降雨激发的土壤呼吸贡献有限.
[1]张超,闫文德,郑威,等.凋落物对樟树和马尾松混交林土壤呼吸的影响[J].西北林学院学报,2013,28(3):22-27. ZHANG C,YAN W D,ZHENG W,et al.Influence of litter on soil respiration in camphor tree-masson pine mixed forest[J].Journal of Northwest Forestry University,2013,28(3):22-27(in Chinese).
[2]MALHIY,GRACEJ.Tropicalforestsand atmospheric carbon dioxide[J]. Tree,2000,15(8):332-337.
[3]DIXON R K,SOLOMON A M,BROWN S,et al.Carbon pools and flux of global forest ecosystems[J].Science,1994,263:185-190.
[4]LAVIGNE M B,RYAN M G,ANDERSON D E,et al.Comparing nocturnal eddy covariance measurements to estimates of ecosystem respiration made by scaling chamber measurements[J].Journal of Geophysical Research,1997,102(24):28977-28985.
[5]LAW B E,RYAN M G,ANTHONI P M.Seasonal and annual respiration of a ponderosa pine ecosystem[J].Global Change Biology,1999,5:169-182.
[6]DAVIDSON E A,BELK E,BOONE R D.Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest[J].Global Change Biology,1998,4:217-227.
[7]JANSSENS I A,LANKREIJER H,MATTEUCCI G,et al.Productivity overshadows temperature in determining soil and ecosystem respiration across European forests[J].Global Change Biology,2001,7:269-278.
[8]SCHLESINGER W H,ANDREWS J A.Soil respiration and the global carbon cycle[J].Biogeochemistry,2000,48(1):7-20.
[9]李玉宁,王关玉,李伟.土壤呼吸作用和全球碳循环[J].地学前缘,2002,9(2):351-357. LI Y N,WANG G Y,LI W.Soil respiration and carbon cycle[J].Earth Science Frontiers,2002,9(2):351-357(in Chinese).
[10]KNORR W,PRENTICE I C,HOUSE J I,et al.Long-term sensitivity of soil carbon turnover to warming[J].Nature,2005,433(7023):298-301.
[11]GORDON H,HAYGARTH P M,BARDGETT R D.Drying and rewetting effects on soil microbial community composition and nutrient leaching[J].Soil Biology and Biochemistry,2008,40(2):302-311.
[12]GESTEL M V,MERCKX R,VLASSAK K.Microbial biomass responses to soil drying and rewetting:the fate of fast and slow growing microorganisms in soils from different climates[J].Soil Biology and Biochemistry,1993,25(1):109-123.
[13]NGAO J,EPRON D,BRECHET C,et al.Estimating the contribution of leaf litter decomposition to soil CO2efflux in a beech forest using 13C-depleted litter[J].Global Change Biology,2005,11(10):1768-1776.
[14]RAICH J W,NADELHOFFER K J.Below ground carbon allocation in forest ecosystems:global trends[J].Ecology,1989,70(5):1346-1354.
[15]BERG B,MCCLAUGHERTY C,PLANT L.Decomposition,Humus Formation,CarbonSequestration[M].2nded.Germany:Springer,2014.
[16]LEE X,WU H,JEFFREY S,et al.Rapid and transient response of soil respiration to rain[J].Global Change Biology,2004,10(6):1017-1026.
[17]CISNEROS-DOZAL L M,TRUMBORE S,HANSON P J.Partitioning sources of soil-respired CO2and their seasonal variation using a unique radio carbon tracer[J].Global Change Biology,2006,12:194-204.
[18]CISNEROS-DOZAL L M,TRUMBORE S,HANSON P J.Effect of moisture on leaf litter decomposition and its contribution to soil respiration in a temperate forest[J].J Geophys Res Biogeosci,2006,112:1-10.
[19]HUXMAN T E,SNYDER K A,TISSUE D,et al.Precipitation pulses and carbon fluxes in semiarid and arid ecosystems[J].Oecologia,2004,141(2):254-268.
[20]BORKEN W,DAVIDSON E A,SAVAGE K,et al.Drying and wetting effects on carbon dioxide release from organic horizons[J].Soil Science Society of America Journal,2003,67(6):1888-1896.
[21]KIM D G,MU S,KANG S,et al.Factors controlling soil CO2effluxes
and the effects of rewetting on effluxes in adjacent deciduous,coniferous,and mixed forests in Korea[J].Soil Biology and Biochemistry,2010,42(4):576-585.
[22]MUHR J,BORKEN W.Delayed recovery of soil respiration after wettingofdrysoilfurtherreducesClossesfrom a Norway spruce forest soil[J]. Journal of Geophysical Research Atmospheres,2009,114(G4):1088.
[23]张太平,任海,彭少麟,等.湿地松(PinuselliottiiEngelm.)的生态生物学特征[J].生态科学,1999,18(2):8-2. ZHANG T P,REN H,PENG S L,et al.The ecological and biological charactristics of Pinuselliottii[J].Ecologic Science,1999,18(2):8-2(in Chinese).
[24]WANG Y D,WANG ZL,WANG H M,et al.Rainfall pulse primarily drives litterfall respiration and its contribution to soil respiration in a young exotic pine plantation in subtropical China[J].Canadian Journal of Forest Research,2012,42(4):657-666.
[25]刘玲玲,刘允芬,温学发,等.千烟洲红壤丘陵区人工针叶林土壤CH4排放通量[J].植物生态学报,2008,32(2):431-439. LIU L L,LIU Y F,WEN X F,et al.CH4emission flux from soil of pine plantations in the Qianyanzhou red earth hill region of China[J]. Journal of Plant Ecology(Chinese Version),2008,32(2):431-439(in Chinese).
[26]褚金翔,张小全.川西亚高山林区三种土地利用方式下土壤呼吸动态及组分区分[J].生态学报,2006,26(6):1693-1700. CHU J X,ZHANG X Q.Dynamic and fractionalization of soil respiration under three different land use/covers in the subalpine region of western Sichuan Province,China[J].Acta Ecologica Sinica,2006,26(6):1693-1700(in Chinese).
[27]SULZMAN E W,BRANT J B,BOWDEN R D,et al.Contribution of aboveground litter,belowground litter,and rhizosphere respiration to total soil CO2efflux in an old growth coniferous forest[J].Biogeochemistry,2005,73:231-256.
[28]JOMURA M,KOMINAMI Y,ATAKA M.Differences between coarse woody debris and leaf litter in the response of heterotrophic respiration torainfallevents[J].JournalofForestResearch,2012,17(3):305-311.
[29]KIEFT T L,SOROKER E,FIRESTONE M K.Microbial biomass response to a rapid increase in water potential when dry soil is wetted[J]. Soil Biology and Biochemistry,1987,19(2):119-126.
[30]WU J,BROOKES P C,WU J,et al.The proportional mineralisation of microbial biomass and organic matter caused by air-drying and rewetting of a grassland soil[J].Soil Biology and Biochemistry,2005,37(3):507-515.
[31]DENOBILI M,CONTIN M,MONDINI C,et al.Soil microbial biomass is triggered into activity by trace amounts of substrate[J].SoilBiology and Biochemistry,2001,33:1163-1170.
[32]WU H H,XU X K,DUAN C T,et al.Effect of vegetation type,wetting intensity,and nitrogen supply on external carbon stimulated heterotrophic respiration and microbial biomass carbon in forest soils[J]. Science China Earth Science,2015,58(8):1446-1456.
[33]XU L,BALDOCCHL D D,TANG J.How soil moisture,rain pulses,and growth alter the response of ecosystem respiration to temperature[J]. Global Biogeochemical Cycles,2004,18(4):187-206.
[34]ATAKA M,KOMINAMI Y,YOSHIMURA K,et al.In situ CO2efflux from leaf litter layer showed large temporal variation induced by rapid wetting and drying cycle[J].Plos One,2014,9(10):108404.
[35]LUO Y,ZHOU X.Soil Respiration and the Environment[M].Burlington:Academic Press,2006.
[36]CROW S E,LAJTHA K,BOWDEN R D,et al.Increased coniferous needle inputs accelerate decomposition of soil carbon in an old-growth forest[J].Forest Ecology and Management,2009,258:2224-2232.
[37]CHEN D,ZHOU L,WU J,et al.Tree girdling affects the soil microbial community by modifying resource availability in two subtropical plantations[J].Applied Soil Ecology,2012,53:108-115.
[38]高强,马明睿,韩华,等.去除和添加凋落物对木荷林土壤呼吸的短期影响[J].生态学杂志,2015,34(5):1189-1197. GAO Q,MA M R,HAN H,et al.Short-term effects of aboveground litter exclusion and addition on soil respiration in a Schimasuperba forest in Zhejiang Province,Eastern China[J].Chinese Journal of Ecology,2015,34(5):1189-1197(in Chinese).
(责任编校 亢原彬)
Simulation research on the impact mechanism of rainfall pulse on soil heterotrophic respiration
LI Yiqiang1,WANG Yidong1,WANG Huimin2,WANG Zhongliang1
(1.a.Tianjin Key Laboratory of Water Resources and Environment,b.College of Urban and Environmental Sciences,Tianjin Normal University,Tianjin 300387,China;2.Institute of Geographic Sciences and Natural Resources Research,Chinese Academy of Sciences,Beijing 100101,China)
In order to investigate the influence mechanism of rainfall pulse on soil heterotrophic respiration,soil+litter-fall+ rainfall(A),soil+rainfall(B),sterilized soil+litter+fall+rainfall+(C),soil sterilization+rainfall(D)and soil+litterfall+no rainfall(CK)treatments were set to study the effects of rainfall pulse on the heterotrophic respiration of the litter in the subtropical pine forest by indoor culture method.The results show that:rainfall stimulated the respirations of litter-fall and mineral soil which both peak at 0.25~1 h,and then gradually recover to the level before rainfall.The duration time of physical exhaust is shorter.The rainfall-induced average contribution of the litter-fall respiration to soil heterotrophic respiration is 93.1%,while the mineral soil respiration is only 6.9%.In summary,the rainfall trigger the litter-fall and mineral soil respirations in varying degrees,but the contribution of physical exhaust to soil respiration is limited,and the litter-fall respiration has a great contribution to soil heterotrophic respiration for a long time(48 h),which serves as the key process for the response of soil heterotrophic respiration to rainfall.
soil CO2emission;soil respiration;litter decomposition;simulated rainfall
P942
A
1671-1114(2016)05-0039-07
2016-05-30
国家自然科学基金资助项目(31300381);天津市高等学校“创新团队培养计划”资助项目(TD12-5037).
李一强(1992—),男,硕士研究生.
王义东(1982—),男,副研究员,主要从事生物地球化学方面的研究.E-mail:wangyidong58@163.com
