木薯MePIL1基因克隆与表达分析
2016-12-12丁泽红铁韦韦付莉莉颜彦夏志强王文泉胡伟
丁泽红 铁韦韦 付莉莉 颜彦 夏志强 王文泉 胡伟
(中国热带农业科学院热带生物技术研究所,海口 571101)
木薯MePIL1基因克隆与表达分析
丁泽红 铁韦韦 付莉莉 颜彦 夏志强 王文泉 胡伟
(中国热带农业科学院热带生物技术研究所,海口 571101)
旨在克隆木薯MePIL1基因,揭示其在木薯遮荫等非生物逆境胁迫中的作用。用RT-PCR的方法从木薯栽培种Ku50叶片中克隆MePIL1基因,对其进行序列比对和系统进化树分析,研究其在木薯野生种和栽培种之间的结构变异,并应用荧光定量PCR技术分析其表达特性。结果显示,克隆获得木薯MePIL1基因,该基因具有一个1 578 bp的开放阅读框,编码525个氨基酸,含有一个HLH保守结构域。系统进化树分析表明,MePIL1与其在杨树和杞柳中的同源基因聚在一起,亲缘关系较近。基因结构变异分析显示,MePIL1在HLH结构域非常保守,其结构变异主要来自于栽培种与野生种之间的差异,且整体上分布比较均匀。表达分析结果显示,MePIL1在叶片中高表达,而且其表达量受到遮荫和渗透胁迫的诱导。结果表明,成功克隆了木薯MePIL1基因,并且MePIL1在转录水平参与木薯对遮荫和渗透胁迫的应答。
木薯;MePIL1;遮荫;基因克隆;表达分析
阳光充足是提高木薯产量的重要保证。作为短日照热带作物,木薯喜阳不耐遮荫,它对光照强度十分敏感。在拉丁美洲、非洲等热带干旱或半干旱地区,农民以求达到土壤经济效益最大化,木薯常常与其它早熟的作物(如玉米、水稻、橡胶和椰子树等)间作在一起[1,2]。大多数时候,特别是植物
生长发育的早期,由于间种的作物长得较快,致使木薯经常遭受到不同程度的遮荫,光照强度降低[3];其次,随着木薯栽培水平的提高,木薯生产区域的扩展和种植密度的增加,使得遮荫的情况也时有发生。遮荫(光照不足)会导致木薯茎叶陡长、节间伸长、茎干细弱、块根变小,最终造成木薯产量显著减少[3]。因此,筛选和鉴定与木薯遮荫相关的重要基因,深入研究其生物学功能,对提高木薯产量具有重要意义。
在遮荫条件下,光强和光质是改变最大的环境因子:如远红光(FR)增加,红光(R)和蓝光减少[4],它们可以作为信号分子诱导植物发生一系列的生理变化。相应地,许多基因的表达量也会发生改变。其中,PIF3-LIKE 1(PIL1)是遮荫条件下加速植物生长所必需的基因之一,编码一个bHLH蛋白[5]。PIL1最早是在拟南芥中通过基因芯片发现的,qRT-PCR证实其表达量在遮荫环境(低R:FR)下可以迅速被诱导;更重要的是,当植物从低R:FR转移到高R:FR时,其表达量则迅速下降[5]。研究表明,PIL1可以被其他基因(如cry1、cry2、pif4、pif5、pif7等)调控[6,7],共同参与遮荫反应。而且,PIL1还能自我反馈调节(自身的表达量)以应对遮荫胁迫[8]。
木薯栽培种是野生种从热带雨林边缘向稀树草原进化而来[9]。经过约7 000-12 000年的驯化,与野生种相比,木薯栽培种具有更好的逆境(如抗病性、耐旱性和耐贫瘠)耐受能力,因而具有更高的生物量和淀粉产量[10]。PIL1被看作是研究植物遮荫反应的标记基因[11]。然而,目前与PIL1相关的研究主要集中在模式植物拟南芥,PIL1在热带作物中的研究还较少,有关木薯PIL1基因克隆及其对遮荫反应的相关分子作用机制尚未见报道。本研究从木薯栽培种Ku50叶片中克隆一个PIL1基因(命名为MePIL1),从系统进化树、基因结构变异、启动子顺式元件和基因表达等方面对它展开了分析,为进一步研究MePIL1的调控功能奠定基础。
1 材料与方法
1.1 材料
供试木薯野生种W14、栽培种Ku50和Arg7由中国热带农业科学院热带生物技术研究所提供。
1.2 方法
1.2.1 材料种植与处理 在木薯种植季节,将Ku50种茎切成长度约15 cm 的茎段,挑选芽眼(3-4个/茎段)且粗细均匀一致的茎段扦插于塑料盆(高18.8 cm,上直径18.5 cm,下直径14.8 cm)中,1茎段/盆。基质采用营养土与蛭石以1∶1的体积比进行混合。木薯种植约10 d后进行间苗,保留1苗/盆。
遮荫处理:遮荫处理的木薯种植于树荫底下,对照处理的木薯种植于室外。种植约2个月后,选择生长状况较一致的植株,收集不同叶片,包括未展开叶、第一片完全展开叶和老叶的样品,-80℃超低温冰箱保存。
PEG处理:种植60 d后,选取长势一致的植株用20%的PEG6000溶液模拟干旱胁迫,以浇灌自来水(不施PEG)为对照。在处理0、3和24 h后,分别收集叶片(包括未展开叶、第一片完全展开叶及老叶)和根的样品,-80℃超低温冰箱保存。
为了考察木薯野生种(W14)和不同栽培种(Ku50和Arg7)中MePIL1基因的表达变化,我们收集了正常大田种植环境下叶片(90 d)、茎(90 d)和储藏根(150 d)的样品,-80℃超低温冰箱保存、备用。
1.2.2 RNA提取与cDNA合成 用RNA提取试剂盒(天根生化科技有限公司)提取不同木薯组织总RNA。用 Revert Aid First Strand cDNA Synthesis Kit(Fermentas公司)将总RNA反转录成cDNA,储存于-20℃备用。
1.2.3 基因克隆 以木薯Ku50叶片的cDNA第一链为模板,用primer6.0软件设计的MePIL1基因全长扩增引物(L1:5'-CAATCGGGTCGTGTTTTGGC-3';R1:5'-TCTTTGAGCCTATGGCGTGT-3')进行PCR扩增。PCR产物经回收、连接和转化后,挑选阳性TA克隆送往华大基因生物科技公司进行测序。
1.2.4 生物信息学分析 用BLASTP搜索Phytozome数据库(https://phytozome.jgi.doe.gov/),获取在其他物种中同源性较高的序列;用ClustalX进行序列比对;用MEGA5.2软件[12]构建进化树(Bootstrap= 1 000);用DnaSPv5[13]进行SNP分析及Ka/Ks计
算;用PlantCARE(http://bioinformatics.psb.ugent.be/ webtools/plantcare/html/)进行启动子元件分析;用CDD数据库(http://www.ncbi.nlm.nih.gov/cdd/)进行保守结构域分析。木薯栽培种Ku50和野生种W14中MePIL1基因序列由前期全基因组测序确定[9];栽培种AM560中MePIL1序列从Phytozome数据库下载。
1.2.5 基因表达分析 用primer6.0软件设计MePIL1基因qRT-PCR特异性引物(L2:5'-TGACTTGTTTCCCGGTTCGT-3';R2:5'-CGTCTTCGCATTCGGGTACT-3')。actin基因引物(L3:5'-TGATGAGTCTGGTCCATCCA-3';R3:5'-CCTCCTACGACCCAATCTCA-3')由前人研究[14]确定。引物均由上海生工生物工程股份有限公司合成。qRT-PCR采用 SYBR Green I试剂盒(TaKaRa公司),按照操作说明在Mx 3005P荧光定量PCR仪(Stratagene,美国)上进行:95℃ 30 s,1个循环;95℃ 30 s,55℃ 30 s,72℃30 s,40个循环;95℃ 1 min,55℃ 30 s,95℃ 30 s,1个循环。每个样品3次生物学重复,表达量按照2-ΔΔCt进行计算[15]。
2 结果
2.1 MePIL1基因的克隆
根据拟南芥AtPIL1序列(AT2G46970),搜索木薯数据库寻找其同源序列(Manes.05G008000.1),之后设计基因特异性引物进行PCR扩增(图1)。测序后得到一个长为1 639 bp的序列,包括33 bp的5' UTR,28 bp的3' UTR及1 578 bp的开放阅读框,将其命名为MePIL1(GenBank登录号:KX147466)。MePIL1编码525个氨基酸。经与木薯参考基因组(https://phytozome.jgi.doe.gov/)比对,MePIL1共有5个外显子,4个内含子(图2)。保守结构域分析表明,MePIL1包含一个HLH domain,位于第3、4外显子上(图2)。
图1 木薯MePIL1基因扩增结果
2.2 MePIL1基因结构变异
为了揭示MePIL1基因结构的变异,将3个木薯品种(包括一个野生种W14,两个栽培种Ku50和AM560)中的MePIL1序列进行比对分析(图2-A),共发现46个SNP,4个Indel(图2-B)。整体上,这些变异分布比较均匀。有趣的是,在HLH保守结构域并没有发现任何变异,暗示HLH区域可能对MePIL1基因功能非常重要。

图2 木薯MePIL1基因比对结果(A)与其结构变异示意图(B)
序列两两比较表明,MePIL1的结构变异主要来自于栽培种与野生种之间的差异:栽培种与栽培种之间的差异很小,只有4个SNP,其中1个在非编码区,3个在编码区;除此之外,其他变异均来自于栽培种与野生种之间。栽培种与野生种Ka/Ks的值介于0.519-0.578之间,暗示MePIL1基因在进化的过程中受到了纯化选择。
2.3 MePIL1进化树分析
经Phytozome数据库搜索,获得了与MePIL1同源性较高的其他物种的序列。进化树分析表明,MePIL1蛋白与杨树(Potri.014G111400.1)和杞柳(SapurV1A.0321s0050.1)聚在一起,亲缘关系较近(图3)。此外,我们还发现,C4植物包括柳枝稷、谷子、高粱、二穗短柄草、玉米和狗尾草聚在一起,而C3植物如白菜型油菜和水稻聚在一起,说明PIL1基因具有C3-C4分化的特性。然而,作为C3植物,拟南芥并没有与白菜型油菜和水稻聚在一起,说明PIL1在C3植物内存在不同的分化。
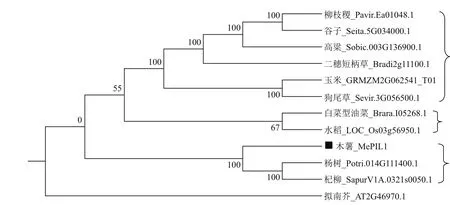
图3 MePIL1系统进化树分析
2.4 启动子元件分析
启动子是基因的一个重要组成部分。我们选取MePIL1起始密码子上游1 500 bp进行启动子元件分析,发现了与ABA作用的元件(如ABRE)、与光相关的元件(如ATCT-motif、Box I和Box 4)、与热胁迫相关的元件(如HSE)和与干旱诱导相关的元件(如MBS)。而且,这些作用元件在野生种和栽培种中都存在。这些结果表明,MePIL1可能参与干旱、光、热等胁迫的响应。
2.5 MePIL1表达分析
本研究考察了MePIL1基因在野生种和栽培种不同组织中的表达量变化。结果(图4-A)表明,在野生种W14中,MePIL1在叶片中表达最高,茎秆其次,根中最低。与野生种相比,MePIL1在栽培种中呈现出相似的表达趋势,然而其在叶片中的表达量要高3-4倍,且表现出叶片特异性表达的特点。这些结果表明,在木薯进化的过程中,MePIL1基因主要倾向于通过提高叶片中的表达量来适应新的生长环境。这可能与该基因在野生种和栽培种之间的碱基变异有关。
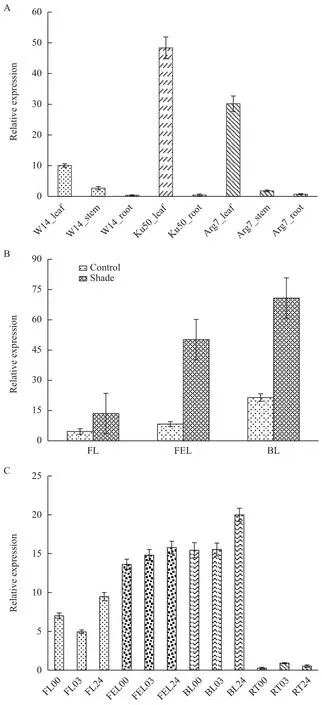
图4 木薯MePIL1基因表达量分析
木薯栽培种是野生种从热带雨林边缘向稀树草原进化而来,其中光(包括光强和光质)是最重要的环境因子。随后,我们在正常和遮荫条件下,分别检测了栽培种Ku50不同叶片中MePIL1基因的表达量。结果(图4-B)表明,MePIL1在遮荫条件下的表达量均显著的高于其在正常条件下的表达量,说明该基因受遮荫胁迫诱导。此外,我们还发现,MePIL1的表达量随着叶片衰老而逐渐增加,暗示该基因可能还参与木薯叶片衰老过程。
在MePIL1启动子区域,我们发现了与ABA作用的元件(如ABRE)和与干旱诱导相关的元件(如MBS),暗示MePIL1可能还参与干旱胁迫。为了初步研究MePIL1对干旱胁迫的响应,我们在PEG 6000模拟条件下,考察了MePIL1在不同组织中表达的动态变化(图4-C)。结果表明,在未展开叶中,MePIL1的表达量呈现先下降(3 h)再上升(24 h)的趋势;在第一片完全展开叶中,MePIL1的表达量呈现逐渐上升的趋势,但差异并不明显;在老叶中,MePIL1的表达量在PEG胁迫3 h后基本没有变化,但在24 h显著上升;在根中,MePIL1的表达量很低,呈现先上升后下降的趋势。这些结果表明,在PEG胁迫处理后,MePIL1在不同叶片和根中的表达量呈现多样性的变化,这将为后续研究其功能提供较好的参考。
3 讨论
木薯是喜阳作物,光照不足(遮荫)显著地减少了木薯产量。作为遮荫反应的标记基因,PIL1在植物遮荫胁迫中扮演着重要角色,但在木薯中尚未见报道。本研究从木薯中克隆了PIL1的同源基因MePIL1。序列分析表明,MePIL1编码525个氨基酸,有5个外显子,4个内含子,亲缘关系与杨树和杞柳较近。与其他PIL1基因类似[5],MePIL1也含有一个保守的HLH结构域。
木薯栽培种是野生种从热带雨林边缘向稀树草原进化而来[9]。与热带雨林相比,稀树草原所处的纬度更高,太阳光要相对弱一些。为了更好地适应新的生长环境,许多与光、高温和水分胁迫等相关的基因在栽培种中的表达均较高[9]。在本研究中,我们发现MePIL1在两个栽培种(Ku50和Arg7)叶片中的表达量均显著高于野生种,暗示这可能是木薯在进化的过程中增强环境适应性的结果。此外,我们还发现MePIL1在野生种和栽培种之间存在大量的碱基变异,但具体哪个变异与基因的表达量相关,目前还不清楚。
与预期相同,在遮荫条件下,MePIL1的表达量被显著诱导。而且,它还能被PEG6000处理诱导,说明MePIL1可能参与除遮荫之外的其他胁迫(如干旱)响应。这一点在水稻中观察到了相似的结论:在干旱条件下,OsPIL1可以作为一个关键调控因子参与节间伸长[16]。除此之外,冷胁迫也显著抑制了OsPIL1的表达量,然而高盐、高温、ABA、GA对OsPIL1的表达量没有影响[16]。
表达分析表明,MePIL1在叶片中特异性表达,而且其表达量随着叶片衰老而逐渐增加,暗示该基因可能参与木薯叶片衰老过程。虽然目前还没有PIL1参与衰老的直接报道,但有研究表明,PIF4和PIF5在拟南芥中能诱导叶片衰老[17];而且PIF4和PIF5能调控其下游靶基因PIL1的表达[18],暗示PIF4和PIF5可能通过PIL1调控木薯叶片衰老。这些结果为进一步研究MePIL1的功能奠定了基础。
4 结论
本研究克隆了木薯PIL基因MePIL1。该基因具有1 578 bp的开放阅读框,编码525个氨基酸,含有HLH保守结构域。系统进化树分析表明,MePIL1与杨树和杞柳的亲缘关系较近。基因结构变异分析显示MePIL1在HLH结构域非常保守,其结构变异主要来自于栽培种与野生种之间的差异。表达分析结果显示,MePIL1在叶片中高表达,而且其表达量受到遮荫和渗透胁迫的诱导。
[1] Mutsaers H, Ezumah H, Osiru D. Cassava-based intercropping:a review[J]. Field Crops Research, 1993, 34(3-4):431-457.
[2] Johnston M, Onwueme I. Effect of shade on photosynthetic pigments in the tropical root crops:yam, taro, tannia, cassava and sweet potato[J]. Experimental Agriculture, 1998, 34(3):301-312.
[3] Okoli PS, Wilson G. Response of cassava(Manihot esculenta Crantz)to shade under field conditions[J]. Field Crops Research, 1986, 14:349-359.
[4] Vandenbussche F, Pierik R, Millenaar FF, et al. Reaching out of the shade[J]. Current Opinion in Plant Biology, 2005, 8(5):462-468.
[5] Salter MG, Franklin KA, Whitelam GC. Gating of the rapid shadeavoidance response by the circadian clock in plants[J]. Nature, 2003, 426(6967):680-683.
[6] Zhang T, Maruhnich SA, Folta KM. Green light induces shade
avoidance symptoms[J]. Plant Physiology, 2011, 157(3):1528-1536.
[7] Nozue K, Tat AV, Devisetty UK, et al. Shade avoidance components and pathways in adult plants revealed by phenotypic profiling[J]. Plos Genet, 2015, 11(4):e1004953.
[8] Li L, Zhang Q, Pedmale UV, et al. PIL1 participates in a negative feedback loop that regulates its own gene expression in response to shade[J]. Molecular Plant, 2014, 7(10):1582-1585.
[9] Wang W, Feng B, Xiao J, et al. Cassava genome from a wild ancestor to cultivated varieties[J]. Nature Communications, 2014, 5:5110
[10] Bredeson JV, Lyons JB, Prochnik SE, et al. Sequencing wild and cultivated cassava and related species reveals extensive interspecific hybridization and genetic diversity[J]. Nature Biotechnology, 2016, 34:562-570.
[11] Wang H, Zhang Z, Li H, et al. CONSTANS-LIKE 7 regulates branching and shade avoidance response in Arabidopsis[J]. Journal of Experimental Botany, 2013, 64(4):1017-1024.
[12] Tamura K, Peterson D, Peterson N, et al. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology and Evolution, 2011, 28(10):2731-2739.
[13] Librado P, Rozas J. DnaSP v5:a software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics, 2009, 25(11):1451-1452.
[14] Xu J, Duan X, Yang J, et al. Enhanced reactive oxygen species scavenging by overproduction of superoxide dismutase and catalase delays postharvest physiological deterioration of cassava storage roots[J]. Plant Physiology, 2013, 161(3):1517-1528.
[15] 胡伟, 颜彦, 韦运谢, 等. 木薯MeASR基因克隆及表达分析[J].生物技术通报, 2015, 31(10):125-130.
[16] Todaka D, Nakashima K, Maruyama K, et al. Rice phytochromeinteracting factor-like protein OsPIL1 functions as a key regulator of internode elongation and induces a morphological response to drought stress[J]. Proceedings of the National Academy of Sciences, 2012, 109(39):15947-15952.
[17] Sakuraba Y, Jeong J, Kang MY, et al. Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis[J]. Nature Communications, 2014, 5:4636.
[18] Hornitschek P, Kohnen MV, Lorrain S, et al. Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling[J]. The Plant Journal, 2012, 71(5):699-711.
(责任编辑 马鑫)
Clone and Expression Analysis of MePIL1 in Cassava
DING Ze-hong TIE Wei-wei FU Li-li YAN Yan XIA Zhi-qiang WANG Wen-quan HU Wei
(Institute of Tropical Bioscience and Biotechnology,Chinese Academy of Tropical Agricultural Science,Haikou 571101)
This work aims to clone MePIL1 gene from cassava and to reveal its functional roles in abiotic stresses(e.g.,shade),which will provide theoretical bases for optimizing the planting density of cassava. MePIL1 gene was cloned from leaf of cassava cultivar ‘Ku50’by RT-PCR method,and the sequences of MePIL1 and its homology genes with other species were aligned and their phylogenetic tree was constructed. Subsequently,structural variations of MePIL1 were revealed between wild and cultivated cassava,and their expression patterns were investigated by quantitative RT-PCR. Results showed that MePIL1,which had a 1 578 bp open reading frame and encoded 525 amino acids and contained a HLH conserved domain,was cloned from cassava. Phylogenetic analysis showed that MePIL1 and homologous genes from Populus trichocarpa and Salix purpurea were clustered together,indicating that they had close genetic relationships. Analysis of gene structural variation revealed that the region of HLH domain was highly conserved,while most variations were derived from the differences between the wild and cultivated cassava species,and the distribution overall was relatively even. Expression pattern analysis showed that MePIL1 was highly expressed in leaf,and its expression was induced by osmotic stress and shade treatments. In conclusion,MePIL1 of cassava was successfully cloned and it was involved in the responses to shade and osmotic stresses at the transcriptional level in cassava.
cassava;MePIL1;shade stress;gene clone;expression analysis
10.13560/j.cnki.biotech.bull.1985.2016.12.013
2016-04-17
海南省自然科学基金项目(20163120,20153048,314123)
丁泽红,男,博士,助理研究员,研究方向:植物分子生物学;E-mail:dingzehong@itbb.org.cn
胡伟,男,博士,副研究员,研究方向:植物分子生物学;E-mail:huwei2010916@126.com
