温度对Bt水稻及其非靶标害虫褐飞虱生态适应性的影响
2016-12-05杨亚军吴志红徐红星郑许松田俊策鲁艳辉吕仲贤
杨亚军, 吴志红, 徐红星, 郑许松, 田俊策, 鲁艳辉, 吕仲贤
浙江省农业科学院植物保护与微生物研究所,浙江省植物有害生物防控重点实验室省部共建国家重点实验室培育基地,浙江 杭州 310021
温度对Bt水稻及其非靶标害虫褐飞虱生态适应性的影响
杨亚军, 吴志红, 徐红星, 郑许松, 田俊策, 鲁艳辉, 吕仲贤*
浙江省农业科学院植物保护与微生物研究所,浙江省植物有害生物防控重点实验室省部共建国家重点实验室培育基地,浙江 杭州 310021
【背景】温度是影响植物生长发育最为重要的环境因子之一。明确温度对Bt水稻及其非靶标害虫的影响有助于今后Bt水稻的推广应用。【方法】设置不同的温度梯度(26、30和34 ℃)分别处理Bt水稻T2A-1(表达Cry2A蛋白),在处理后不同天数内分别测定Cry蛋白、叶绿素、草酸、可溶性糖和总氮含量,同时评价不同温度处理下Bt水稻上褐飞虱的生态适应性。【结果】Cry蛋白含量显著受温度影响,随着温度升高Cry蛋白含量下降。温度、处理时间及两者相互作用均显著影响Bt水稻T2A-1的叶片叶绿素、叶鞘草酸、可溶性糖和总氮含量。叶绿素含量在26 ℃处理20 d时高于30、34 ℃的处理。随着温度的升高,Bt水稻叶鞘草酸含量升高。可溶性糖含量高温(30 和34 ℃)处理后20 d低于26 ℃处理。Bt水稻叶鞘内可溶性氮含量随着温度的升高而升高。温度对非Bt水稻MH63上叶绿素、草酸、可溶性糖和总氮含量的影响与T2A-1趋势一致。高温(34 ℃)处理的Bt水稻上的褐飞虱雌雄虫寿命、每雌产卵量及卵孵化率显著低于26 ℃处理。温度对非Bt水稻MH63上褐飞虱生态适应性参数的影响与T2A-1趋势一致。【结论与意义】温度对Bt水稻具有显著影响,且对Bt水稻上非靶标害虫褐飞虱的生态适应性产生不利影响。本研究有利于今后Bt稻田的害虫综合治理措施的制定。
温度;Bt水稻; 生理; 褐飞虱; 生态适应性
苏云金芽胞杆菌(Bacillusthurigiensis,Bt)是一种革兰氏阳性菌,其在孢子发育过程中可产生具有杀虫作用的Cry蛋白(Chenetal., 2011)。随着现代生物技术的发展,已有多种Bt基因转入水稻(Chenetal.,2011; Cohenetal.,2008)。Bt水稻的使用能减少环境污染,促进粮食安全生产,对发展优质无公害稻米生产具有重要的经济效益、社会效益和显著的生态效益,可为无公害稻米的生产提供保障和支撑(Huangetal.,2005; Lu,2010; Tanetal.,2011; Zhangetal.,2007)。环境因子是影响植物生长的重要因素之一,目前,已有多篇报道涉及温度、肥料、旱涝、盐胁迫等因素对Bt作物的游离氨基酸、可溶性糖及Bt蛋白表达量的影响((张顺等,2011; Bruns & Abel,2002; Chenetal.,2005; Coviellaetal.,2002; Jiangetal.,2006; Luoetal.,2008; Kauretal.,2011; Wangetal.,2012)。
温度是重要的农业生态因子之一,其变化将会对农作物的生产产生重大影响。IPCC(2007)预计,2016—2035年全球平均地表温度与1986—2005年相比将升高0.3~0.7 ℃。而浙江省绝大部分地区(岱山、普陀和武义除外)温度呈升高趋势,大部分地区(衢州、金华和丽水西部地区除外)超过35 ℃的天数也为增加趋势(宁金花和申双和,2009; 唐国利等,2009)。温度上升将会改变农作物种植区域、种植时间、生长季,对农作物的生长发育、产量等产生重要影响(徐铭志和任国玉,2004; 赵俊芳等,2009; Songetal.,2010; Taoetal.,2006)。极端高温还会造成农作物的热害,经济损失严重(秦叶波和张慧,2015)。
在全球气温变暖的背景下,水稻生产容易受到温度变化的影响。王保菊等(2010)研究发现,3个不同水稻品种中的可溶性糖含量均随温度升高而增加,而草酸含量则随温度升高先增加后下降,在25 ℃时最高。温度升高也会对水稻的抗虫性产生影响,IR26和IR36的抗虫性从25 ℃起随温度升高而减弱,特别是在自然条件下具中等抗性的IR36,在31和34 ℃下与感虫品种TN1一样抗性完全丧失(王保菊等,2010)。温度变化可以对Bt作物产生影响,如高温使Bt棉花蛋白酶和肽酶活性提高,丙酮酸转氨酶活性下降,但氨基酸含量增加,说明可溶性蛋白分解增强(陈德华等,2003)。Bt水稻育成以来,关于Bt水稻对非靶标害虫褐飞虱NilaparvatalugensStål影响的研究也逐渐增多,从单纯研究Bt水稻上褐飞虱的生态适应性到环境因子的介入研究。研究表明,Bt水稻对褐飞虱生态适应性指标无显著影响,药剂对Bt水稻上褐飞虱再猖獗的诱导能力与非Bt水稻上无异(李向冬等,2014; Chenetal.,2011; Luetal.,2015; Yangetal., 2013)。温度变化对Bt水稻及其非靶标害虫的影响关系到Bt水稻的发展与应用。本文通过设置不同的温度梯度,研究了温度对Bt水稻生理状况的影响,同时评价了不同温度处理下Bt水稻上非靶标害虫褐飞虱的生态适应性,可为气候变化特别是全球气候变暖背景下Bt水稻上害虫种群动态预测、防治策略的制定提供依据,为Bt水稻的推广与应用奠定基础。
1 材料与方法
1.1 供试水稻品系
Bt水稻品系T2A-1(含cry2A基因)及其亲本非Bt水稻品种MH63由华中农业大学提供。TN1由国际水稻研究所提供。水稻品种(系)定期种植于浙江省农科院试验田内,常规管理,种植期间不使用任何农药。
1.2 供试虫源
褐飞虱:2014年采自杭州市郊常规水稻田,养虫室内用TN1苗连续饲养,同日初羽化成虫供试验。
1.3 试验设计
恒温培养箱设3个温度梯度(26、30、34 ℃),选取长势相近的水稻植株(T2A-1),每个品种选15盆,每盆7~8株,每个温度下培养4盆。待恒温培养箱达到预定温度后,移入盆栽水稻并开始计时,不同天数后取样试验。
1.4 不同温度处理水稻体内生理生化指标变化
1.4.1 Cry蛋白含量测定 T2A-1水稻经不同温度处理后0、5、10、20 d,取样测定Cry2A蛋白含量。取相近位置的叶片0.1 g,液氮磨碎后采用Envirologix公司试剂盒所示方法测定Cry蛋白含量。
1.4.2 叶绿素含量 测定不同温度处理后0、10、20 d的Bt水稻叶片的叶绿素含量,以其亲本非Bt水稻MH63为对照。选取水稻植株相近位置的倒叶,用叶绿素测定仪测定其叶片中叶绿素含量。每片叶片测定3次取均值,重复6次。
1.4.3 草酸含量 测定不同温度处理后0、10、20 d的Bt水稻叶鞘的草酸含量,以其亲本非Bt水稻MH63为对照。草酸含量测定采用三氯化钛显色法(展海军等,2006)。取稻苗1 g,剪碎后充分研磨,用20 mL超纯水冲洗至50 mL三角瓶中,加1/3体积活性炭,摇匀后静置30 min脱色,脱色后离心分离活性炭,一次脱色不完全时可重复脱色直至溶液呈无色或略呈乳白色。取上清液2 mL,于4 ℃、3000 r·min-1下离心15 min。取0.5 mL上清液,加40 μL 3%三氯化钛(10%稀盐酸配制),加超纯水至1 mL,取300 μL,测D400 nm。用草酸(分析纯)作标准曲线,计算各样品中草酸的含量。
1.4.4 可溶性糖含量 测定不同温度处理后0、10、20 d的Bt水稻叶鞘的可溶性糖含量,以其亲本非Bt水稻MH63为对照。采用蒽酮比色法(王强盛等,2008)测定可溶性糖含量。将各处理稻苗样品烘干至恒重,取0.1 g左右放入大试管中,加入15 mL蒸馏水,沸水浴20 min,取出冷却,过滤入100 mL容量瓶中,用蒸馏水冲洗残渣数次,定容。取待测样品提取液1.0 mL加蒽酮试剂5 mL,将各管快速摇动混匀后,沸水浴10 min,取出冷却后测D620 nm。用无水葡萄糖(分析纯)作标准曲线,计算各样品中可溶性糖的含量。
1.4.5 总氮含量 测定不同温度处理后0、10、20 d的Bt水稻叶鞘的总氮含量,以其亲本非Bt水稻MH63为对照。将水稻样品烘干,用粉碎机粉碎均匀后,用EURO元素分析仪测定总氮含量。
1.5 温度对Bt水稻上褐飞虱生态适应性的影响
评价不同温度处理后Bt水稻上褐飞虱的生态适应性,以非Bt水稻MH63作对照。将水稻植株清洗干净,剥去最外层叶鞘后,置于试管中,接1对初羽化褐飞虱成虫,试管以棉花封口,置于原温度的培育箱中。每个温度设置10个重复。每天观察水稻及褐飞虱存活情况,记录产出若虫数量,直至连续5 d未观察到褐飞虱若虫再孵出时为止。解剖水稻茎秆记录未孵化卵数。计算褐飞虱雌雄成虫寿命、产卵量和卵孵化率。
1.6 数据分析
采用软件SPSS 18.0进行方差分析,并用Tukey多重比较法测验各参数不同处理间的差异显著性。百分率先进行反正弦平方根转换后再作方差分析。
2 结果与分析
2.1 不同温度处理水稻体内生理生化指标变化
经不同温度处理后Bt水稻T2A-1体内Cry蛋白的变化见图1。随着温度升高Bt蛋白含量下降,其中处理20 d后3个温度下T2A-1水稻体内Bt蛋白含量差异最大。方差分析结果表明,T2A-1叶片Cry蛋白含量显著受温度(P<0.001)、处理时间(P<0.001)及温度×处理时间(P<0.001)的影响。
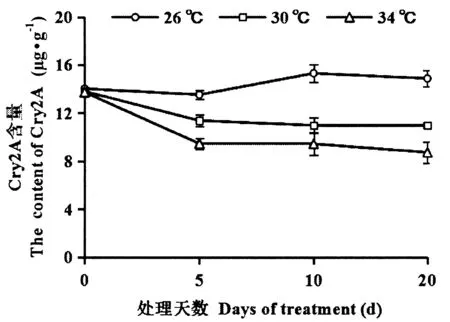
图1 经温度处理的T2A-1水稻叶片中Cry蛋白的变化
Bt水稻在30和34 ℃下经历20 d后,叶片叶绿素含量显著低于26 ℃处理,非Bt水稻MH63上也观察到同样的趋势(表1)。方差分析表明,Bt水稻上叶绿素含量显著受温度(P=0.008)、处理时间(P<0.001)及温度×处理时间(P=0.043)的影响。非Bt水稻MH63叶片叶绿素含量也显著受温度(P=0.019)、处理时间(P<0.001)及温度×处理时间(P=0.048)的影响。
Bt水稻经30和34 ℃处理10 d后叶鞘草酸含量高于26 ℃处理,34 ℃处理20 d后草酸含量高于26和30 ℃处理,非Bt水稻MH63经34 ℃处理10 d后草酸含量高于26 ℃处理,34 ℃处理20 d后草酸含量高于26 ℃处理(表2)。方差分析表明,Bt水稻上草酸含量显著受温度(P<0.001)、处理时间(P<0.001)及温度×处理时间(P<0.001)的影响。非Bt水稻叶鞘草酸含量显著受温度(P<0.001)、处理时间(P<0.001)及温度×处理时间(P=0.004)的影响。
Bt水稻T2A-1经30和34 ℃处理10、20 d后叶鞘可溶性糖含量均低于26 ℃处理(表3)。非Bt水稻MH63叶鞘中34 ℃处理20 d后叶鞘可溶性糖含量均低于26和30 ℃处理。方差分析表明,Bt水稻T2A-1叶鞘可溶性糖含量显著受温度(P<0.001)、处理时间(P<0.001)及温度×处理时间(P<0.001)的影响,非Bt水稻MH63叶鞘可溶性糖含量显著受温度(P<0.001)、处理时间(P<0.001)及温度×处理时间(P<0.001)的影响。
Bt水稻T2A-1经26和30 ℃处理10 d后叶鞘氮含量低于34 ℃处理,处理20 d氮含量随温度降低而降低(表4)。非Bt水稻MH63上叶鞘氮含量受温度的影响趋势与T2A-1一致。方差分析表明,Bt水稻T2A-1和非Bt水稻MH63叶鞘可溶性氮含量均显著受温度(P<0.001)、处理时间(P<0.001)及温度×处理时间(P<0.001)的影响。
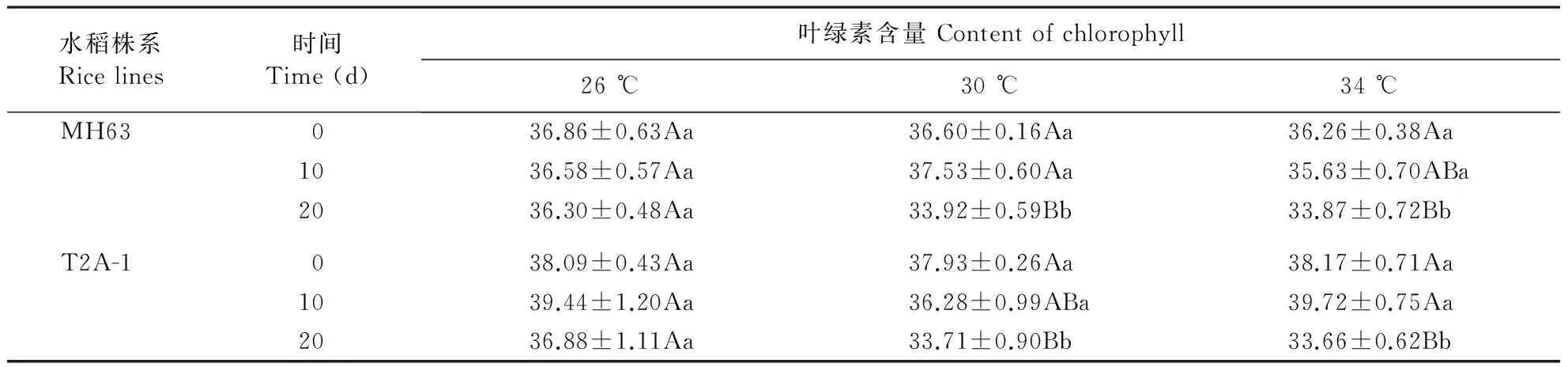
表1 不同温度处理下Bt水稻T2A-1和其亲本非Bt水稻MH63叶片叶绿素含量(SPAD值)
不同小写字母表示不同温度之间具有显著差异。不同大写字母表示同一品种不同处理天数之间具有显著差异。
Different lowercase stands for significant differences between treatments with different temperatures. Different uppercase stands for significant differences between data of same rice line with different treating times.
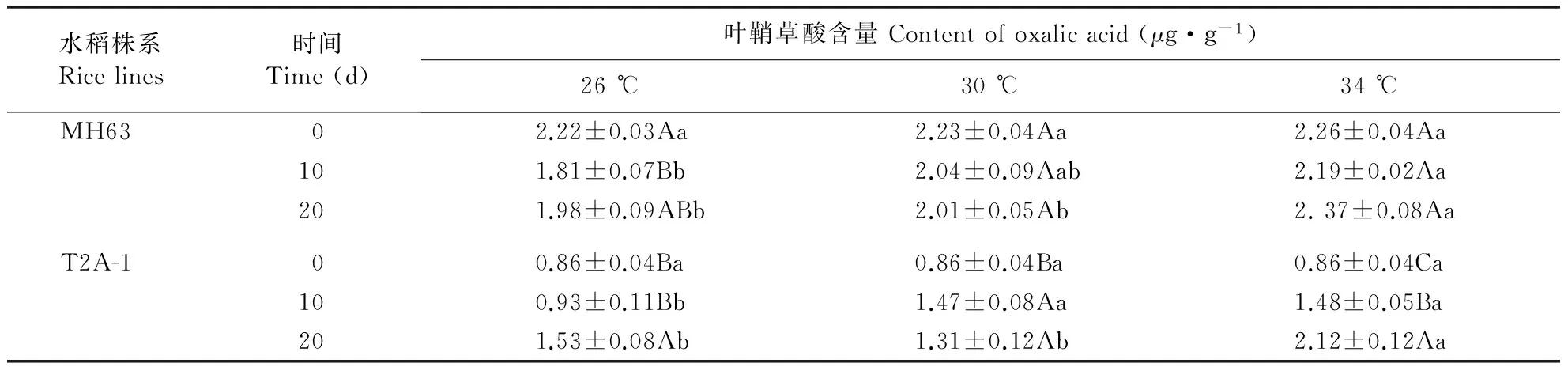
表2 不同温度处理下Bt水稻T2A-1和其亲本非Bt水稻MH63叶鞘草酸含量
不同小写字母表示不同温度之间具有显著差异。不同大写字母表示同一品种不同处理天数之间具有显著差异。
Different lowercase stands for significant differences between treatments with different temperatures. Different uppercase stands for significant differences between data of same rice line with different treating times.
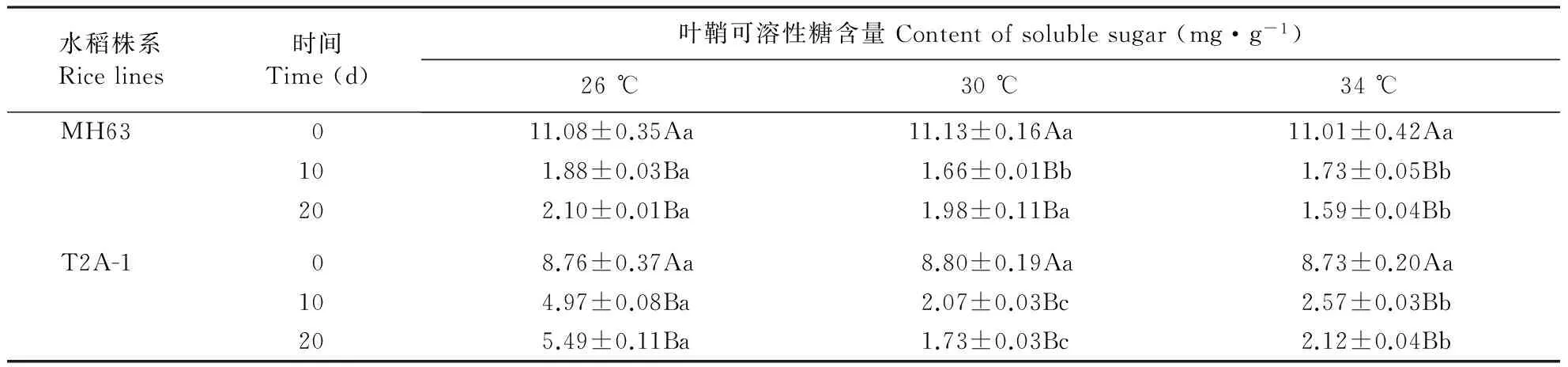
表3 不同温度处理下Bt水稻T2A-1和其亲本非Bt水稻MH63叶鞘可溶性糖含量
不同小写字母表示不同温度之间具有显著差异。不同大写字母表示同一品种不同处理天数之间具有显著差异。
Different lowercase stands for significant differences between treatments with different temperatures. Different uppercase stands for significant differences between data of same rice line with different treating times.
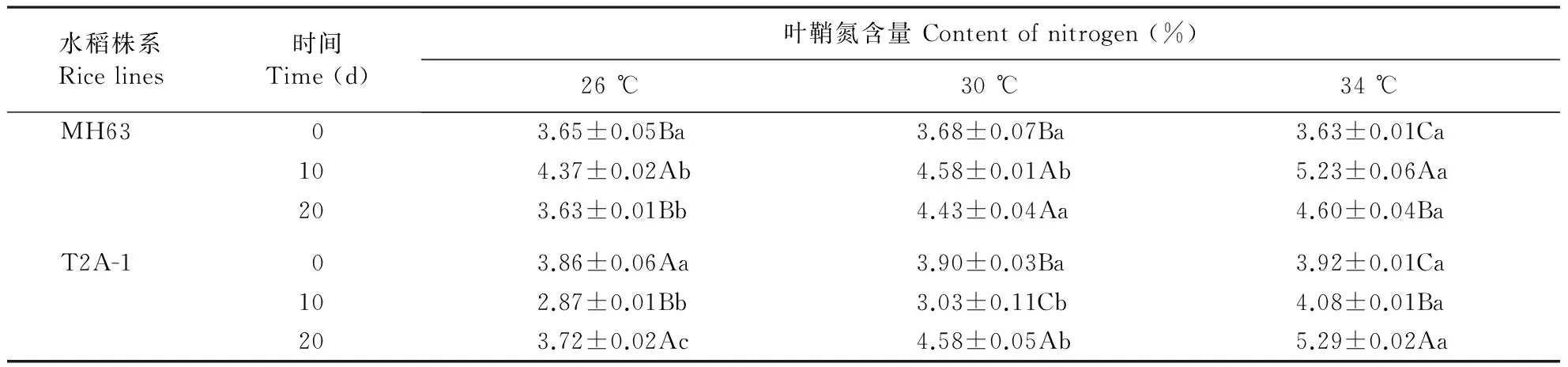
表4 不同温度处理下Bt水稻T2A-1和其亲本非Bt水稻MH63叶鞘氮含量
不同小写字母表示不同温度之间具有显著差异。不同大写字母表示同一品种不同处理天数之间具有显著差异。
Different lowercase stands for significant differences between treatments with different temperatures. Different uppercase stands for significant differences between data of same rice line with different treating times.
2.2 温度对Bt水稻上褐飞虱生态适应性的影响
不同温度处理下Bt水稻T2A-1上褐飞虱的生态适应性见图2。雌成虫寿命随着处理温度的升高呈降低趋势,雄成虫寿命在26 ℃最高。每雌产卵量26 ℃下最高,30 ℃次之,34 ℃下最低。卵孵化率26 ℃下高达93.01%,30 ℃下降为59.85%,34 ℃下仅为25.20%。在非Bt水稻对照MH63上,褐飞虱生态适应性参数随着温度的变化趋势与T2A-1上相同。
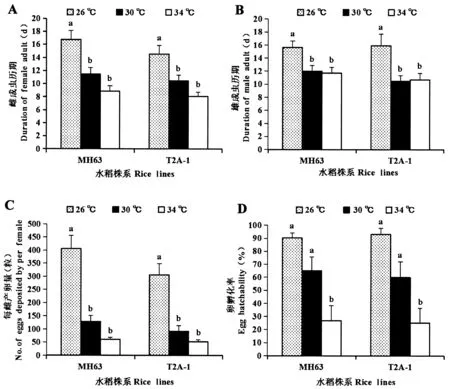
图2 温度对Bt水稻T2A-1和其亲本非Bt水稻MH63上褐飞虱生态适应性的影响
3 讨论
温度是植物生长发育必需的环境因子之一,在全球气候变暖的背景下,温度变化对植物的影响尤其受到关注。本文研究表明,Bt水稻T2A-1体内Cry蛋白在高温处理下较常温处理表达量低,说明Bt水稻体内Cry蛋白表达量受温度变化的影响。同样,在Bt棉花和Bt玉米上也有环境因子影响Bt蛋白含量的报道(陈德华等,2003; 张桂玲和温四民,2011)。Bt蛋白是Bt水稻中的主要杀虫成分之一,目前,Bt水稻体内设计的Cry蛋白表达量均比较高,但暂时还未进行商业化推广。稻纵卷叶螟CnaphalocrocismedinalisGuenee 等害虫对其还处于敏感阶段,由高温引起的Cry蛋白含量的降低尚不足以降低水稻对稻纵卷叶螟等害虫的抗性。然而,今后随着Bt水稻的推广应用,环境因子对Bt蛋白含量的影响需引起高度重视,以便及时采取措施延缓害虫对Bt水稻产生抗性。
叶绿素是水稻叶片进行光合作用的重要物质之一。Bt水稻在26 ℃下处理20 d时比其他2个温度(30、34 ℃)高,长时间高温会影响叶绿素的生成。草酸含量与褐飞虱取食量有关,草酸含量增加可降低褐飞虱取食量,增强水稻的抗虫性(Yoshiharaetal.,1980)。本文结果显示,Bt水稻体内草酸含量随温度的升高而升高。王保菊等(2010)也发现,22~34 ℃范围内随着温度升高草酸含量先增加后减少,25 ℃时最高,从而水稻的抗虫性较高。Bt水稻T2A-1叶鞘可溶性糖含量高温(30、34 ℃)处理后20 d含量低于26 ℃处理,与罗泽民等(1982)研究结果一致。Bt水稻T2A-1叶鞘内可溶性氮含量随着温度的升高而升高。褐飞虱喜欢在含氮量高的水稻植株上产卵,高氮含量植株上褐飞虱卵孵化率高于低氮含量植株(吕仲贤等,2005)。温度对Bt水稻T2A-1生理参数变化趋势影响与非Bt水稻MH63上一致。
温度是褐飞虱种群生命系统中一个重要的因子,温度变化能通过影响其生长发育、生殖来影响种群的发生动态(石保坤等,2014; 祝树德等,1994)。王保菊(2010)研究了非Bt水稻上温度变化对褐飞虱生态适应性的影响,发现温度过高会降低褐飞虱的生态适应性。褐飞虱成虫在39.5 ℃暴露2 h,其存活率显著降低44.4%,缓和高温(31、33 ℃)对褐飞虱成虫hsp70表达具有诱导作用(季璐等,2015)。本文研究表明,高温条件下褐飞虱雌雄虫寿命、每雌产卵量及卵孵化率低于常温处理。不同温度下非Bt水稻MH63上褐飞虱的生态适应性与Bt水稻T2A-1上变化趋势一致。近年来,全球气候变暖,特别是暖冬或者夏凉秋暖的温度格局有利于褐飞虱的生长发育与繁殖。多个研究表明,Bt水稻对褐飞虱生态适应性几乎无影响,也不会引起褐飞虱大发生,Bt水稻种植条件下药剂诱导褐飞虱再猖獗的能力与非Bt水稻上无异(李向冬等,2014; Chenetal.,2011; Luetal.,2015; Yangetal.,2013)。本文研究表明,温度对Bt水稻上褐飞虱的影响与非Bt水稻上趋势一致。本文在全球气候变暖的背景下研究温度对Bt水稻及非靶标害虫褐飞虱的影响,为今后Bt稻田非靶标害虫褐飞虱防治策略的制定奠定基础。然而,Bt水稻田褐飞虱种群数量变化还受许多生物与非生物因子的影响,且多种情况下为多种因子相互作用的结果。今后,环境因子—Bt水稻—褐飞虱之间相互关系的研究还需加强。
陈德华, 杨长琴, 陈源, 聂安全, 吴云康, 2003. 高温胁迫对Bt棉叶片杀虫蛋白表达量和氮代谢影响的研究. 棉花学报, 15(5): 288-292.
色谱柱:ZorbaxSB-C8(4.6mm×75mm,3.5 μm)或效能相当的色谱柱,辛烷基硅烷键合硅胶为填充剂;流动相:磷酸盐缓冲液(取磷酸二氢钾3.4 g与三乙胺15 mL,加1 000 mL水使其溶解,用磷酸调节pH至 5.5)-甲醇-四氢呋喃(体积比为 587∶323∶90);流速:1.5 mL/min;紫外检测波长:230 nm;进样量:10 μL;柱温:45℃;出峰顺序:度洛西汀峰、杂质Ⅴ峰、杂质Ⅲ峰与杂质Ⅷ峰;理论板数:不低于2500。
季璐, 崔旭红, 朱敏, 2015. 短时高温胁迫对褐飞虱存活及其热激蛋白基因hsp70表达的影响. 植物保护学报, 42(4): 619-625.
李向冬, 杨亚军, 田俊策, 徐红星, 郑许松, 吕仲贤, 2014. 转Bt基因及其亲本水稻上褐飞虱对三唑磷和溴氰菊酯的反应. 应用生态学报, 25(10): 3006-3010.
罗泽民, 唐明远, 彭忠魁, 1982. 杂交水稻抗稻褐飞虱的机制. 湖南农业科学 (1): 9-12.
吕仲贤, Heong K L, 俞晓平, 胡萃, 2005. 稻株含氮量和密度对褐飞虱存活、发育和生殖特性的影响. 生态学报, 25(8): 1838-1843.
宁金花, 申双和, 2009. 气候变化对中国农业的影响. 现代农业科技 (12):251-254, 256.
秦叶波, 张慧, 2015. 水稻高温热害发生规律及防御措施. 浙江农业科学, 56(9): 1362-1365.
石保坤, 胡朝兴, 黄建利, 侯茂林, 2014. 温度对褐飞虱发育、存活和产卵影响的关系模型. 生态学报, 34(20): 5868-5874.
唐国利, 丁一汇, 王绍武, 任国玉, 刘洪滨, 张莉, 2009. 中国近百年温度曲线的对比分析. 气候变化研究进展, 5(2): 71-78. 王保菊, 徐红星, 郑许松, 傅强, 吕仲贤, 2010. 温度对水稻抗褐飞虱特性的影响. 中国水稻科学, 24(4): 443-446.
徐铭志, 任国玉, 2004. 近40年中国气候生长期的变化. 应用气象学报, 15(3): 306-312.
展海军, 周长智, 张立, 弓彦忠, 2006. 啤酒中草酸含量的测定. 酿酒科技 (4): 102-104.
张顺, 陈刚, 房卫平, 马宗斌, 谢德意, 李伶俐, 朱伟, 2011. 施氮量对抗虫棉Bt蛋白表达和降解的影响. 华北农学报, 26(6): 148-153. 张桂玲, 温四民, 2011. 盐胁迫对转Bt基因棉苗期Bt蛋白表达量和氮代谢的影响. 西北农业学报, 20(6): 106-109.
赵俊芳, 杨晓光, 刘志娟, 2009. 气候变暖对东北三省春玉米严重低温冷害及种植布局的影响. 生态学报, 29(12): 6544-6551.
祝树德, 陆自强, 杭衫保, 徐海, 1994. 温度对褐飞虱种群调控作用研究. 华东昆虫学报, 3(1): 53-59.
Bruns H A and Abel C A, 2002. Nitrogen fertility effects on Bt δ-endotoxin and nitrogen concentrations of maize during early growth.AgronomyJournal, 95(1): 207-211.
Chen D H, Ye G Y, Yang C Q, Chen Y and Wu Y K, 2005. The effect of high temperature on the insecticidal properties ofBtcotton.EnvironmentalandExperimentalBotany, 53(3): 333-342.
Chen M, Shelton A and Ye G Y, 2011. Insect-resistant genetically modified rice in China: from research to commercialization.AnnualReviewofEntomology, 56: 81-101.
Cohen M B, Chen M, Bentur J S, Heong K L and Ye G Y, 2008.Btrice in Asia: potential benefits, impact, and sustainability∥Romeis J, Shelton A M and Kennedy G G.IntegrationofInsect-ResistantGeneticallyModifiedCropswithinIPMPrograms. New York: Springer Science+Business Media B V: 223-248.
Coviella C E, Stipanovic R D and Trumble J T, 2002. Plant allocation to defensive compounds: interactions between elevated CO2and nitrogen in transgenic cotton plants.JournalofExperimentalBotany, 53: 323-331.
Huang J K, Hu R F, Rozelle S and Pray C, 2005. Insect-resistant GM rice in farmers′fields: assessing productivity and health effects in China.Science, 308: 688-690.
IPCC, 2007.ClimateChange2007,thePhysicalScienceBasis,ContributionofWorkinggroupItotheFourthAssessmentReportoftheIntergovernmentalPanelonClimateChange. Cambridge: Cambridge University Press.
Jiang L J, Duan L S, Tian X L, Wang B M, Zhang H F, Zhang M C and Li Z H, 2006. Nacl salinity stress decreasedBacillusthuringiensis(Bt) protein content of transgenicBtcotton (GossypiumhirsutumL.) seedlings.EnvironmentalandExperimentalBotany, 55(3): 315-320.
Kaur N, Sohal B S and Singh K, 2011. Biochemical and physiological changes onBacillusthuringiensiscotton after imidacloprid foliar spray.PesticideBiochemistryandPhysiology, 99(3): 280-284.
Lu C M, 2010. The first approved transgenic rice in China.GMCrops, 1(3): 113-115.
Lu Z B, Liu Y E, Han N S, Tian J C, Peng Y F, Hu C, Guo Y Y and Ye G Y, 2015. Transgenic cry1C or cry2A rice has no adverse impacts on the life-table parameters and population dynamics of the brown planthopper,Nilaparvatalugens(Hemiptera: Delphacidae).PestManagementScience, 71(7): 937-945.
Luo Z, Dong H Z, Li W J, Ming Z and Zhu Y Q, 2008. Individual and combined effects of salinity and waterlogging on Cry1Ac expression and insecticidal efficacy ofBtcotton.CropProtection, 27(12): 1485-1490.
Song Y L, Linderholm H W, Chen D L and Walthe A, 2010. Trends of the thermal growing season in China, 1951—2007.InternationalJournalofClimatology, 30(1): 33-43.
Tan T, Zhao J T and Chen C, 2011. The impact of commercialization of GM rice in China.American-EurasianJournalofAgricultural&EnvironmentalSciences, 10(3): 296-299.
Tao F L, Yokozawa M, Xu Y L, Hayashi Y and Zhang Z, 2006. Climate changes and trends in phenology and yields of field crops in China, 1981—2000.AgriculturalandForestMeteorology, 138: 82-92.
Wang F, Jian Z P, Nie L X, Cui K H, Peng S B, Lin Y J and Huang J L, 2012. Effects of N treatments on the yield advantage ofBt-SY63 over SY63 (Oryzasativa) and the concentration of Bt protein.FieldCropsResearch, 129: 39-45.
Yang Y J, He J J, Dong B Q, Xu H X, Fu Q, Zheng X S, Lin Y J and Lu Z X, 2013. Effects of twoBtrice lines T2A-1 and T1C-19 on the ecological fitness and detoxification enzymes ofNilaparvatalugens(Hemiptera: Delphacidae) from different populations.JournalofEconomicEntomology, 106(4): 1887-1893.
Yoshihara T, Sogawa K, Pathak M D, Juliano B O and Sakamura S, 1980. Oxalic acid as a sucking inhibitor of the brown planthopper in rice (Delphacidae, Homoptera).EntomologiaExperimentalisetApplicata, 27(2): 149-155.
Zhang Q F, 2007. Strategies for developing green super rice.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 104(42): 16402-16409.
(责任编辑:郭莹)
Effects of temperature onBtrice and ecological fitness of the non-target insect pestNilaparvatalugens
Ya-jun YANG, Zhi-hong WU, Hong-xing XU, Xu-song ZHENG,Jun-ce TIAN, Yan-hui LU, Zhong-xian LÜ*
StateKeyLaboratoryBreedingBaseforZhejiangSustainablePestandDiseaseControl,InstituteofPlantProtectionandMicrobiology,ZhejiangAcademyofAgriculturalSciences,Hangzhou,Zhejiang310021,China
【Background】 Temperature is one of the most important factors that influences the growth and development of crop plants. Understanding the effects of temperature onBtrice and its non-target insects is helpful for the implement and management ofBtrice in future. 【Method】 The contents of Cry protein, chlorophyll, oxalic acid, soluble sugar and nitrogen were investigated in theBtrice plants that were exposed to different temperatures (26, 30 and 34 ℃) for different durations. Ecological fitness of brown planthopperNilaparvatalugenswas evaluated under these different temperatures. 【Result】 The Cry protein content was significantly affected with decreasing Cry protein as the temperature increased. Temperature, treatment duration and their interaction were significantly for the content of chlorophyll, oxalic acid, soluble sugar and nitrogen inBtrice. The content of chlorophyll inBtrice and soluble sugar ofBtrice shezth treated for 20 d at 26 ℃ was higher than at 30 and 34 ℃. The content of oxalic acid and nitrogen inBtrice sheath increased with increasing temperatures. The effect of temperature on the chlorophyll, oxalic acid, soluble sugar and nitrogen in non-Btrice MH63 were similar to T2A-1. Longevity of male and female adults, number of eggs deposited per female and egg hatchability ofN.lugensonBtrice was lower at 34 ℃ than at 26 ℃. The effect of temperature on the ecological fitness of the nontarget insect pestN.lugensin non-Btrice MH63 was similar to T2A-1. 【Conclusion and significance】 Temperature could influence the physiology ofBtrice and negatively affect the ecological fitness of the nontarget insect pestN.lugensinBtrice. This study will facilitate the makeup of the integrated management of insect pests in theBtrice in future.
temperature;Btrice; physiology;Nilaparvatalugens; ecological fitness
2016-03-15 接受日期(Accepted): 2016-05-23
国家转基因生物新品种培育重大专项(2014ZX08001-001); 国家水稻产业技术体系(CARS-01-17); 浙江省自然科学基金(LY15C140003)
杨亚军, 男, 副研究员。 研究方向: 水稻害虫综合治理。 E-mail: yargiuneyon@163.com
*通讯作者(Author for correspondence), E-mail: luzxmh2004@aliyun.com
10. 3969/j.issn.2095-1787.2016.04.008
