珍稀树种花榈木种子休眠破除方法研究
2016-12-04贵州大学林学院贵阳550025长顺县林业局贵州长顺550700
, (.贵州大学林学院, 贵阳 550025; 2.长顺县林业局, 贵州 长顺 550700)
·研究报告·
珍稀树种花榈木种子休眠破除方法研究
邓兆1,2,韦小丽1
(1.贵州大学林学院, 贵阳 550025; 2.长顺县林业局, 贵州 长顺 550700)
为了破除花榈木种子的休眠,对花榈木种子进行药剂处理、混沙湿藏处理以及激素+混沙湿藏处理,对处理后的种子进行发芽试验,测定种子的发芽指标,并测定混沙湿藏不同阶段种子内部脱落酸、赤霉素、酸性磷酸酯酶、淀粉酶、蛋白酶、ATP等指标的变化。结果表明:激素+混沙湿藏处理的种子萌发效果优于混沙湿藏和药剂处理。用500 mg/L赤霉素浸泡花榈木种子12 h,再低温(4 ℃)混沙湿藏45 d是破除花榈木种子休眠的最佳方法,其发芽开始时间、结束时间和高峰期分别比不处理的种子提早10,17 d和14 d,发芽率、发芽势分别比对照提高36.6%、32.7%。混沙湿藏处理的花榈木种子的酸性磷酸酯酶、淀粉酶和蛋白酶活性、GA3和ATP含量随着混沙湿藏的时间延长而增加,ABA含量则随着混沙湿藏时间的延长而减小,混沙湿藏对破除花榈木种子休眠起关键作用。
花榈木; 种子休眠; 药剂处理; 混沙湿藏; 激素处理
花榈木(OrmosiahenryiPrain)是珍贵用材树种和优良的园林绿化树种,同时是高级的家具、装饰品、高级地板的原材料。由于品质优良,花榈木遭大量砍伐,人类的不合理利用使其资源十分稀少,处于濒危状[1],为国家二级保护植物。为保护和培育花榈木资源,近年来人们加强了对花榈木的研究,主要在种子休眠[2]、无性繁殖与组织培养[3-4]、人工林生长规律[5-6]、播种育苗技术[7]、对不同水分、光照的响应[8-9]及容器苗基质筛选[10]等方面,但对破除花榈木种子休眠的方法尚未见研究。种子休眠是植物在长期演化过程中为了不断适应环境而形成的生存对策,种子休眠对植物个体生存、物种的延续和进化具有积极的作用,但对林木的生产带来较大的影响。本试验采用赤霉素、6-BA、化学药剂、混沙湿藏等方法对花榈木种子进行处理,以探讨破除花榈木种子休眠的方法,提高花榈木种子的发芽率和发芽势,使其发芽整齐,为大量繁殖花榈木提供理论依据。
1 材料与方法
1.1 试验材料
试验用的花榈木种子于2014年11月中旬采自贵州关岭县白水镇,去除荚果皮后,放在实验室阴干,保存于4 ℃左右的冰箱中。种子千粒重455.36 g,含水率为12.03%。
1.2 试验设计及方法
设计4个处理,Ⅰ: 0.2%硝酸钾处理种子12 h;Ⅱ: 0.5%高锰酸钾处理种子12 h;Ⅲ: 0.2%硝酸钾处理种子12 h+100 mg/L赤霉素处理12 h;Ⅳ: 0.5%高锰酸钾处理种子12 h+100 mg/L赤霉素处理12 h,以清水浸泡12 h作对照。每个处理3次重复,每次重复100粒种子,进行发芽试验,测定发芽指标。
1.2.2 混沙湿藏处理种子试验
设计3个处理,Ⅴ:持续暖温(15~20 ℃)层积,先对种子进行消毒灭菌,浸种;然后把种子装入小尼龙网袋中,每袋100粒种子,用湿沙混匀后,置于塑料袋中进行层积45 d,并经常翻动种子;Ⅵ: 持续低温(0~4 ℃)层积;Ⅶ:变温层积,先在15~20 ℃条件下层积1个月,然后转入0~4 ℃的条件下层积15 d。
上述贮藏温度分别采用人工气候箱和冰箱控制,每个处理3次重复,每次重复100粒种子,对其进行发芽试验,测定发芽指标。
1.2.3 不同激素处理种子试验
将赤霉素和细胞分裂素分别配制成0,50,100,150,250,500,1 000 mg/L的浓度,分别浸泡种子12 h后进行混沙湿藏,温度控制在0~4 ℃,每个处理3次重复,每次重复100粒种子,贮藏45 d后从沙中取出洗净,然后用80 ℃的高温水浸泡24 h,再进行发芽试验。发芽床为沙床,置床前用0.15%的甲醛溶液对发芽盒、种子、镊子进行消毒处理,沙放在130 ℃的烘箱内高温烘24 h进行杀菌处理。每个处理置床3个,置床后分别放在25 ℃恒温的光照培养箱中进行发芽实验。从置床之日起开始观察记录,直到发芽结束为止,每天统计发芽数。
(2)船钞的进一步降低。第二次鸦片战争后,美国抢在英法之前,强迫清政府签订中美《天津条约》,条约第二十六款明确提出:“凡在一百五十吨以上者,每吨纳银四钱,不及一百五十吨者,每吨纳银一钱。”要求清政府降低船钞标准,随后中英《天津条约》第二十九款也有类似规定。
1.2.4 混沙湿藏不同时间段种子的生理生化变化测定
取花榈木种子进行混沙湿藏,测定湿藏0,15,30,45 d种子的脱落酸、赤霉素、酸性磷酸酯酶、淀粉酶、蛋白酶、ATP等生理生化指标的变化,每个指标测定3次重复。
酸性磷酸酯酶活性、ATP含量、淀粉酶活性和蛋白酶活性参照陈润政[11]的方法测定,脱落酸和赤霉素参照曹帮华等[12]的方法测定。
1.3 数据处理
1.3.1 主要指标计算方法
发芽率(%)=正常幼苗数/供试种子总数×100%;
发芽势(%)=发芽达到发芽高峰时的累计发芽种子数/供试种子总数×100%。
1.3.2 统计分析方法
试验数据采用SPSS 18.0软件进行方差分析和多重比较。
2 结果与分析
2.1 不同处理破除花榈木种子休眠的效果分析
2.1.1 不同药剂组合处理的效果比较
与对照相比,不同药剂处理的花榈木种子发芽开始时间、发芽结束时间、发芽高峰期均比对照提前,不同处理种子发芽率、发芽势差异显著(p≤0.05)。用0.5%高锰酸钾处理种子12 h+100 mg/L赤霉素处理12 h(Ⅳ)效果最好,其发芽开始时间、结束时间和发芽高峰期分别比对照提前6 d,9 d和9 d,发芽率、发芽势分别比对照提高10.6%、7.7%,且与对照和Ⅰ、Ⅱ处理差异显著(p≤0.05)。
3种不同温度的混沙湿藏处理种子发芽率、发芽势差异显著(plt;0.05)。其中Ⅵ处理(持续低温0~4 ℃层积)的效果最佳,发芽开始时间和发芽结束时间都较其它2个处理短,发芽率、发芽势均最高,持续暖温(15~20 ℃)层积效果最差,变温层积效果居中。但不论是哪种混沙湿藏处理,其效果都优于药剂处理。
2.1.2 不同浓度赤霉素处理的效果比较
赤霉素是有利种子萌发的激素。从表3可知,随着赤霉素浓度的增加,种子的各项发芽指标均呈增加趋势。用100~1 000 mg/L赤霉素处理的种子,发芽率、发芽势分别比ck 1提高4%~14%和3%~12%。以500 mg/L赤霉素浸泡种子12 h后再混沙湿藏45 d,然后用80 ℃高温水浸泡24 h效果最佳,其发芽开始时间、结束时间和高峰期分别比ck 1提早7 d,8 d和8 d,发芽率、发芽势分别比ck1提高14%和12%,比ck 2分别提早10 d、17 d和14 d,提高36.6%、32.7%。赤霉素浓度为1 000 mg/L时,种子的发芽率、发芽势与赤霉素浓度为500 mg/L时差异不显著(plt;0.05),50~150 mg/L赤霉素处理的种子发芽率、发芽势与对照差异均不显著(plt;0.05),说明赤霉素浓度过低,对种子萌发的促进作用极小。
表1 不同药剂处理对花榈木种子发芽的影响
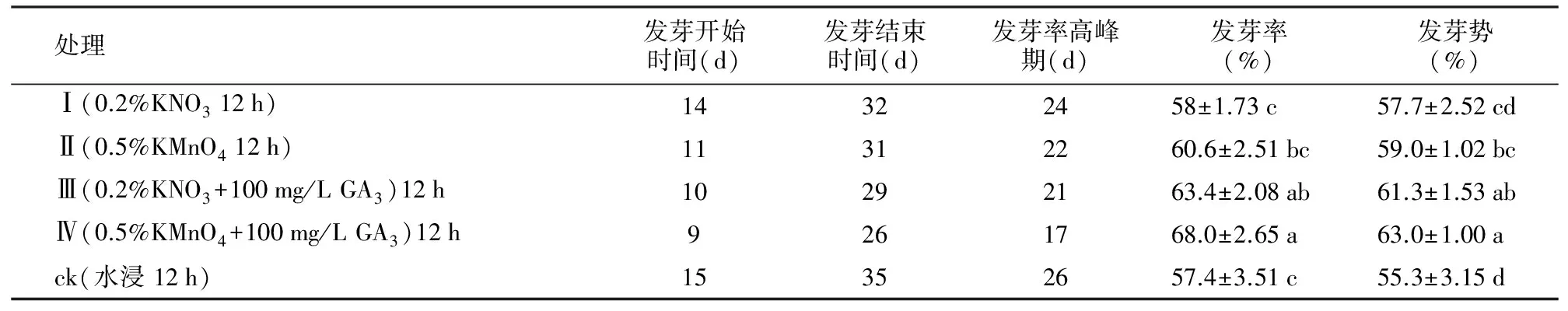
处理发芽开始时间(d)发芽结束时间(d)发芽率高峰期(d)发芽率(%)发芽势(%) Ⅰ(0.2%KNO312h)14322458±1.73c57.7±2.52cd Ⅱ(0.5%KMnO412h)11312260.6±2.51bc59.0±1.02bc Ⅲ(0.2%KNO3+100mg/LGA3)12h10292163.4±2.08ab61.3±1.53ab Ⅳ(0.5%KMnO4+100mg/LGA3)12h9261768.0±2.65a63.0±1.00a ck(水浸12h)15352657.4±3.51c55.3±3.15d
注:不同小写字母分别表示不同地区相同分析指标间存在显著差异(plt;0.05)。下同。
表2 不同混沙湿藏处理对花榈木种子发芽的影响

处理发芽开始时间(d)发芽结束时间(d)发芽率高峰期(d)发芽率(%)发芽势(%) Ⅴ(15-20℃)45d15292375.7±1.52b73±2.00bc Ⅵ(0~4℃)45d12262081.3±2.08a77±1.73a Ⅶ(15~20℃30d,0~4℃15d)14272177.7±2.51ab75±1.00ab
表3 不同浓度的赤霉素处理对花榈木种子发芽的影响

GA3(mg/L)发芽开始时间(d)发芽结束时间(d)发芽率高峰期(d)发芽率(%)发芽势(%)5011231880±1.73d79±1.70de10010221784±3.60cd80±1.00cd1509211686±2.66cd82±1.73bcd2507201488±2.65bc83±1.73bc5005181294±4.58a88±1.65a10005171292±3.60ab85±1.00abck1(混沙湿藏)12252080±2.65d76±2.45eck2(不处理)15352657.4±3.51e55.3±3.15f
2.1.3 不同浓度6-BA处理的效果比较
由表4可知,随着6-BA浓度的增加,花榈木种子的各项发芽指标都逐渐增加。50 mg/L 6-BA处理的种子发芽率与对照差异不显著(plt;0.05),这说明6-BA浓度过低,对种子的发芽基本没有促进作用。用1 000 mg/L 6-BA浸泡花榈木种子12 h后再混沙湿藏45 d,然后用80 ℃高温水浸泡24 h效果最好,其发芽开始时间、结束时间以及发芽高峰期分别比对照提早5 d,6 d和8 d,其发芽率、发芽势分别比对照提高8%、5%。说明6-BA处理有助于花榈木种子提早发芽,提高种子发芽率和发芽势,但效果不及赤霉素处理。
表4 不同浓度的6-BA处理对花榈木种子发芽的影响

6⁃BA(mg/L)发芽开始时间(d)发芽结束时间(d)发芽率高峰期(d)发芽率(%)发芽势(%)5011251980±2.65c77±2.45cd10011241884±2.20b79±2.02cd15010221684±1.00b80±1.02abc2509211586±1.63ab81±2.65ab5007191386±1.73ab83±2.00a10007191288±1.68a83±1.00a0(ck)12252080±2.65c76±1.73d
2.2 混沙湿藏过程中种子生理生化指标的变化
测定结果(表5)表明,花榈木种子的酸性磷酸酯酶、淀粉酶、蛋白酶活性、GA3含量、ATP含量均随混沙湿藏的时间增加呈逐渐增加趋势,且差异显著(plt;0.05)。贮藏45 d时,各指标比贮藏前分别增加115.6%、188.8%、180.7%、71.8%和92%;酸性磷酸酯酶增加量最大,GA3增加量较小;ABA含量随着混沙湿藏时间的增加而减小,比贮藏前降低58.0%,且不同贮藏时间ABA含量差异显著(plt;0.05)。花榈木种子的各种酶活性随着混沙湿藏时间的增加而增强,有助于贮藏物质的分解释放;促进种子萌发激素GA3含量增加,而抑制种子萌发激素ABA含量降低,从而破除种子休眠。
3 结论与讨论
花榈木种子具有休眠特性[2],严重影响了种子萌发力,给花榈木的繁育带来不便。本研究采用不同药剂处理、不同混沙湿藏温度处理、不同激素结合混沙湿藏处理花榈木种子,结果表明,混沙湿藏是打破种子休眠、促进种子萌发的必要条件。混沙湿藏过程中,花榈木种子的酸性磷酸酯酶、淀粉酶、蛋白酶等酶活性和GA3、ATP含量均随着混沙湿藏的天数增加而增加,而ABA含量则随着混沙湿藏时间的增加而减小,说明混沙湿藏有效地改善了种子酶活性和激素比例,从而促进种子萌发。混沙湿藏以0~4 ℃低温贮藏45d效果最佳,发芽率达81.3%,发芽势为77%。混沙湿藏再加外源GA3、6-BA处理则进一步加强了种子内部有利于种子萌发的激素比例,其中以500 mg/L赤霉素浸泡种子12 h后再混沙湿藏45 d,然后用80 ℃高温水浸泡24 h效果最佳,发芽率、发芽势分别高达94%、88%,比不作任何处理的种子分别提高36.6%、32.7%,发芽开始时间和结束时间分别提早10,17 d。
表5 不同混沙湿藏时间段花榈木种子中生化指标的变化

生化指标0d15d30d45d酸性磷酸酯酶活性[nmol/(min·g)]15.06±0.02d18.93±0.03c25.36±0.03b32.47±0.02a淀粉酶活性[mg/(min·g)]2.15±0.03d3.42±0.04c5.68±0.05b6.21±0.03a蛋白酶活性(μmol/g)1.82±0.03d2.39±0.05c3.46±0.04b5.11±0.03aABA含量(μg/g)4.29±0.02a3.84±0.03b2.59±0.03c1.81±0.04dGA3含量(μg/g)3.16±0.03d3.95±0.05c4.81±0.04b5.43±0.03aATP含量(μg/g)0.25±0.02d0.39±0.04c0.43±0.04b0.48±0.04a
混沙湿藏处理可促使胚形态发育成熟,抑制物质含量降低,激素含量发生变化,提高酶的活性,促进种子萌发[13]。本研究结果表明,随着种子混沙湿藏时间的延长,花榈木种子的酸性磷酸酯酶、淀粉酶、蛋白酶等酶活性和GA3、ATP含量均随着混沙湿藏时间的延长而增加,而ABA含量则随着混沙湿藏时间的延长而减小,种子发芽率、发芽势得以大幅度提高,这与前人研究的毛梾种子[14]、珙桐[15]、青钱柳[16]等休眠种子特性相似。
赤霉素、ABA和细胞分裂素对种子的休眠和萌发起重要作用。种子休眠的三因素调控的学说中指出:GA是种子萌发的必需激素,种子中无足够量GA,种子不可能萌发;ABA是诱发种子休眠的主要激素,种子中虽有GA但同时存在ABA,种子休眠,因为ABA抑制GA作用的发挥;而细胞分裂素并不单独对休眠与萌发起作用,不是萌发所必需的激素,但能抵消ABA的作用使因存在ABA而休眠的种子萌发[17]。本研究中得出结论,混沙湿藏过程中GA3含量增加,而ABA含量下降,花榈木种子休眠被打破。前人的研究还表明,种子贮藏中施加外源赤霉素、6-BA处理,能有效打破由于ABA等植物激素萌发抑制物造成的种子休眠[13],本研究采用外源赤霉素、6-BA处理种子,再混沙湿藏,比单独的混沙湿藏处理更能有效地打破花榈木种子的休眠,其原因是外源的赤霉素、6-BA与混沙湿藏的作用叠加,加速了种子内部激素比例变化,因而效果最佳。
[1]孟宪帅.珍稀濒危植物花榈木致濒原因及机理研究[D].贵州大学硕士论文,2011.
[2]邓兆,韦小丽,孟宪帅,等.花榈木种子休眠和萌发的初步研究[J].贵州农业科学,2011,39(5):69-72.
[3]姚军,李洪林,杨波,等.花榈木的组织培养和快速繁殖[J].植物生理学通讯,2007,43(1):123-124.
[4]高丽,杨波,李洪林,等.花榈木组培苗茎段低温胁迫培养及耐冷性诱导[J].亚热带植物科学,2009,38(2):19-21,25.
[5]张都海,袁位高,陈承良,等.花榈木人工林生长规律的初步研究[J].浙江林业科技,2003,23(3):9-11,27.
[6]傅林清.花榈木人工林主要测树因子回归模型的研究[J].林业勘察设计,2012(1):24-27.
[7]彭丽.珍贵乡土树种花榈木播种育苗技术[J].林业实用技术,2012(2):30-30.
[8]孟宪帅,韦小丽.不同水分环境对花榈木幼苗生理生化的影响[J].山地农业生物学报,2011,30(3):215-220.
[9]段如雁,韦小丽,孟宪帅.不同光照条件下花榈木幼苗的生理生化响应及生长效应[J].中南林业科技大学学报,2013,33(5):30-34.
[10]段如雁,韦小丽,张怡,等.花榈木容器育苗基质筛选[J].林业科技开发,2015,29(4):27-30.
[11]黄学林,陈润政.种子生理实验手册[M].北京:农业出版社,1990.
[12]曹帮华,蔡春菊.银杏种子后熟生理与内源激素变化的研究[J].林业科学,2006,42(2):32-37.
[13]陈伟,马绍宾,陈宏伟.种子休眠类型及破除方法概述[J],安徽农业科学,2009,37(33):16 237-16 239.
[14]李冬林,张亚楠,金雅琴,等.毛梾种子低温层积过程中内源激素的变化及其与发芽的关系[J].经济林研究,2014,32(3):36-41.
[15]雷泞菲,彭书明,牛蓓.珍稀濒危植物珙桐种子休眠萌发过程中内源激素的变化[J].广西植物,2009,29(1):66-69.
[16]杨万霞,方升佐.青钱柳种子综合处理过程中内源激素的动态变化[J].南京林业大学学报(自然科学版),2008,32(5):85-88.
[17]沈海龙.苗木培育学[M].北京:中国林业出版社,2009.
Study on Method of Breaking Seed Dormancy for Rare Tree SpeciesOrmosiahenryi
DENGZhao1,2,WEIXiaoli1
(1.Forestry College of Guizhou University,Guiyang 550025,China;2.Forestry Bureau of Changshun County,Changshun Guizhou 550700,China)
For breaking seed dormancy ofOrmosiahenryi,theOrmosiahenryiseed were treated by medicament,stratification,hormone combine with stratification.The germination experiment of seed after treatment were determined,and the change of ABA,GA3,acid phosphatase,diastase,protease and ATP of seed were determined at different stage after stratification.The results showed the germinate effect of seed by handing with hormone combine with stratification was better than stratification and medicament.The method that soaked seed for 12 h using 500 mg/L GA3,then stratification for 45 d under 4 ℃ was the best method for breaking dormancy ofOrmosiahenryiseed,and the begin time,end time,peak time of germination ahead of 10 d,17 d and 14 d than control,the germination rate,germination potential improved 36.6% and 32.7%.The acid phosphatase, diastase and protease activity,GA3and ATP content ofOrmosiahenryiseed increased with the time of stratification;and the ABA content decreased;the stratification played a key role for breaking the dormancy ofOrmosiahenryiseed.
Ormosiahenryi; seed dormancy; medicament treatment; stratification;hormone treatment
2016-05-19
国家自然科学基金“花榈木根瘤菌多样性及幼苗形成调控因素研究”(编号:31460193);国际合作项目“珍贵用材树种花榈木精准化容器育苗理论与技术研究”(编号:黔科合外G字[2012]7012号);贵州省林业厅重大攻关项目“贵州优质阔叶用材树种培育技术与示范”(编号:黔林科合[2010]重大02)。
邓 兆(1985—),男(苗族),贵州晴隆人;助理工程师,主要从事林木种苗繁育研究。
韦小丽,E-mail:gdwxl-69@126.com。
10.16590/j.cnki.1001-4705.2016.11.001
Q 945.79
A
1001-4705(2016)11-0001-04
