温度对3种金缕梅科植物种子萌发特性的影响
2016-12-04广西植物功能物质研究与利用重点实验室广西植物研究所广西桂林54006中国科学院华南植物园广东广州50650国际植物保护联盟BGCI广东广州50650
, , , , (.广西植物功能物质研究与利用重点实验室,广西植物研究所, 广西 桂林 54006;2.中国科学院华南植物园, 广东 广州 50650; 3.国际植物保护联盟(BGCI), 广东 广州 50650)
温度对3种金缕梅科植物种子萌发特性的影响
王满莲1,文香英2,3,韦霄1,蒋运生1,唐辉1
(1.广西植物功能物质研究与利用重点实验室,广西植物研究所, 广西 桂林 541006;2.中国科学院华南植物园, 广东 广州 510650; 3.国际植物保护联盟(BGCI), 广东 广州 510650)
选用金缕梅科濒危植物半枫荷和广布种阿丁枫与枫香树为研究材料,比较研究了不同温度(15、20、25、30 ℃)对3种植物种子萌发特性的影响,以期从种子生态学的角度探讨半枫荷濒危的机制。结果表明:半枫荷的种子最大,其千粒重显著大于阿丁枫与枫香树。3种金缕梅科植物的种子萌发曲线相似,4个实验温度下均能萌发,并且萌发率高、萌发速度快、萌发持续时间短。3种植物15 ℃下的种子萌发率均最低,但半枫荷与阿丁枫的萌发率均能达到70%以上,枫香树能达到50%以上;其中半枫荷各温度下的种子萌发率均显著高于阿丁枫和枫香树,25 ℃下能达90%。随着温度的升高,3种植物种子的萌发开始时间和萌发持续时间均显著缩短,其中半枫荷15 ℃和30 ℃下的萌发开始时间早于其它2种植物, 15 ℃下的萌发持续时间长于其它2种植物。总体而言,3种金缕梅科植物种子萌发适应的温度范围均较广,其中半枫荷的种子萌发能力最强。推测半枫荷对单个种子的投入大,种子集中在某一时段萌发,种群在受到强烈干扰时难以短期内恢复种群规模,这可能是其濒危的因素之一。
半枫荷; 阿丁枫; 枫香树; 种子萌发; 温度
半枫荷(SemiliquidambarcathayensisH.T.Chang)为金缕梅科半枫荷属植物,高大常绿乔木,国家Ⅱ级保护植物,为我国特有种,它具有枫香属和蕈树属两属间的综合性状,在学术研究、中医药用、林木加工、园林绿化等方面均具极高价值[1]。目前该物种仅残存于我国南部和东南山区, 即广西、湖南、江西、广东等省山区地带,一般分布在海拔1 000 m以下。枫香树(LiquidambarformosanaHance)系金缕梅科枫香树属的高大落叶乔木,我国秦岭及淮河以南各省份均有分布,其生长快、用途广、适应性强、生态效益好,已成为林种、树种结构调整的首选树种[2]。阿丁枫(Altingiachinensis(Champ.) Oliv,ex Hance)为金缕梅科蕈树属常绿乔木,又名蕈树。分布于浙江、福建、湖南、广东、海南、江西、广西、云南、贵州等地,常生于海拔200~1 000 m的常绿阔叶林中的山谷、沟边。生长迅速,树形优美,材质致密、坚韧,是优良用材、药用及园林绿化观赏树种[3]。笔者在野外实地调查过程中发现,半枫荷野外多以孤立木存在,结实有大小年之分,种子产量低,树下没有幼苗更新,而枫香树与阿丁枫种群均成片存在,林下有幼苗更新。半枫荷与枫香树和阿丁枫亲缘关系较近,生态习性相近,分布区有重叠,并且同样作为经济树种受到大量采挖,为什么半枫荷与枫香树和阿丁枫在种群大小上有如此巨大的差别?
植物濒危的原因除了人类过度利用及生境的破坏外,还有自身生物学方面的因素[4]。种子萌发是植物生活史中最重要的环节之一,是其种群自然更新的基础[5]。对许多濒危植物的研究发现,种子质量差、萌发率低、被取食严重、萌发所需的条件与其生境不协调等是造成其濒危的重要原因[6],种子萌发特性的系统研究对于濒危植物种群的保护与恢复及其人工繁殖具有重要意义。影响植物种子萌发的因素有温度、储藏条件、种子大小以及母株生境等[7-8],其中温度对种子萌发有着十分重要的影响,可以直接影响无休眠种子的萌发速率,还可以通过影响种子的休眠状态,间接地影响种子萌发[9-11]。本试验比较研究半枫荷与同科植物半枫荷和枫香树3种植物种子萌发对温度处理的应答,从种子生态学的角度探讨温度对3个金缕梅科植物种子萌发的影响,试图为探讨半枫荷濒危机制和人工繁育提供理论依据。
表1 3种金缕梅科植物种子的形态特征
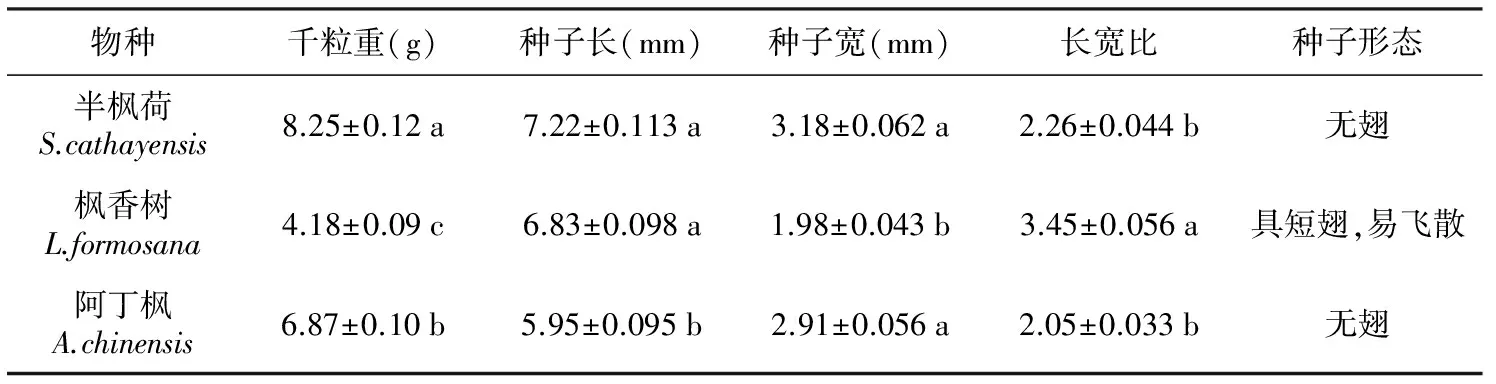
物种千粒重(g)种子长(mm)种子宽(mm)长宽比种子形态半枫荷S.cathayensis8.25±0.12a7.22±0.113a3.18±0.062a2.26±0.044b无翅枫香树L.formosana4.18±0.09c6.83±0.098a1.98±0.043b3.45±0.056a具短翅,易飞散阿丁枫A.chinensis6.87±0.10b5.95±0.095b2.91±0.056a2.05±0.033b无翅
注:同列不同小写字母表示种间差异显著(plt;0.05)。
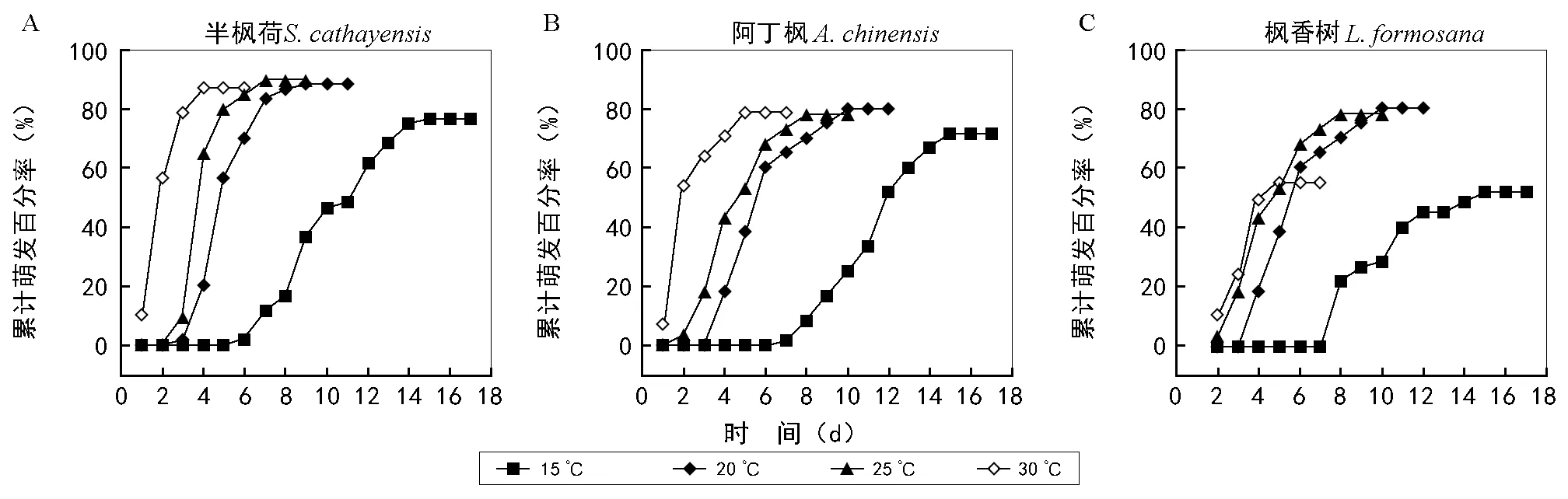
图1 不同温度下3种金缕梅科植物种子的萌发过程
1 材料与方法
1.1 植物材料及种源地概况
研究地区位于广西壮族自治区桂林市,研究地区概况见马姜明等研究[12]。半枫荷、阿丁枫和枫香树种子均于2014年11月采自桂林周边山区,分别是灵川县大镜乡、公平乡和雁山区的自然居群。
1.2 形态指标测定
每一物种成熟的种子采自同一母株,蒴果风干后开裂,稍加敲打,使种子散落,去杂后得纯净种子并干藏。每个物种随机抽取30粒种子,用游标卡尺分别测量种子长和宽, 重复3次,并计算种子长宽比。每个物种随机抽取1 000粒种子,用万分之一天平分别进行称重,重复3次,计算千粒重。
1.3 种子萌发试验
2015年3月5日,挑选半枫荷、阿丁枫和枫香树籽粒饱满、大小基本一致的种子,洗净后用0.1%的KMnO4溶液消毒15 min,再用纯净水洗3~4遍,用滤纸吸干表面水分后,放入直径为12 cm铺有2层滤纸的培养皿内,加入5 mL纯净水,并加皿盖保湿,放入RSZ智能型人工气候箱 。设置 4 个恒温(15、20、25、30 ℃)条件,每个温度3次重复,每个重复随机选取 20 粒种子,所有萌发在 12 h/12 h(光照/黑暗) 的培养条件下进行,光照强度为35μmol/m2。每天在相同的时间观察记录发芽种子数,以胚根突破种皮1 mm作为种子发芽的标准,当连续3 d没有种子再萌发时,视为发芽结束。主要萌发指数:萌发率,即萌发结束后总的萌发百分数(%);萌发开始时间,从播种到第1粒种子萌发所需的时间(d);萌发持续时间,即开始萌发到萌发结束所需时间(d);萌发速率指数(GR)=G1/T1+G2/T2+…+Gt/Tt,Tt为相应的萌发日数,Gt为t时间的萌发百分数。
1.4 数据分析
数据处理采用SPSS 13.0软件,种间与温度间差异显著性用单因素方差分析(One-way ANOVA)检验,用LSD显著性检验方法比较不同处理间的差异性。用Sigma Plot 11.0(SPSS Inc.,USA)绘图。
2 结果与分析
2.1 3种金缕梅科植物种子形态比较
由表1可见, 半枫荷种子的千粒重显著大于阿丁枫与枫香树,种子长与枫香树无显著差异、但显著大于阿丁枫,种子宽与阿丁枫无显著差异、但显著大于枫香树,半枫荷与阿丁枫的种子长宽比无显著差异,但均显著小于枫香树。枫香树的种子具短翅,易飞散传播,半枫荷与阿丁枫的种子无翅。
2.2 温度对3种金缕梅科植物种子萌发进程的影响
如图1所示,3种金缕梅科植物的种子萌发曲线十分相似,尤其是半枫荷与阿丁枫的种子萌发曲线相似度更高。总体而言,随着温度的升高3种植物种子的开始萌发时间越早,萌发结束的时间也越早。半枫荷和阿丁枫在20、25、30 ℃3个温度下的最终萌发率相差不大, 15 ℃下的最终萌发率均低于前3个温度;枫香树在20 ℃和25 ℃ 2个温度下的最终萌发率基本接近,15 ℃和30 ℃下的最终萌发率低于前2个温度。总体而言,3种金缕梅科植物的萌发开始时间和持续时间均较短,30 ℃下半枫荷与阿丁枫种子1 d便能萌发,枫香树种子2 d便能萌发,萌发均持续5 d左右;25 ℃下3种植物均在第2~3天开始萌发,萌发均持续5~7 d;20 ℃下第3~4天开始萌发,萌发均持续6~7 d;15 ℃下半枫荷种子6 d左右就能萌发,枫香树和阿丁枫种子第7~8天能萌发,3种植物萌发均持续8~10 d。

注:不同小写字母表示同种不同温度间差异显著,不同大写字母表示相同温度下不同种间差异显著(plt;0.05)。图2 不同温度下3种金缕梅科植物种子的萌发指数
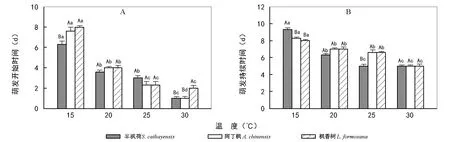
图3 不同温度下3种金缕梅科植物种子的萌发时间
2.3 温度对3种金缕梅科植物种子萌发指数的影响
进一步统计分析表明,温度对3种金缕梅科植物种子的最终萌发率和萌发速率指数均具有显著的影响(图2)。半枫荷和阿丁枫15 ℃下的最终萌发率和萌发速率指数均显著小于20~30 ℃,20、25、30 ℃ 3个温度下的最终萌发率无显著差异,但30 ℃下的萌发速率指数显著高于20 ℃和25 ℃;枫香树在20 ℃和25 ℃ 2个温度下的最终萌发率和萌发速率指数均无显著差异,但均显著高于30 ℃和15 ℃。3种植物相比较,15 ℃下,半枫荷的最终萌发率和萌发速率指数均显著高于阿丁枫,而阿丁枫的萌发率又显著高于枫香树;20 ℃和25 ℃2个温度下,半枫荷的最终萌发率均显著高于阿丁枫和枫香树(后两者无显著差异),但3种金缕梅科植物的种子萌发速率指数均无显著差异;30 ℃下,半枫荷的最终萌发率均显著高于阿丁枫,而阿丁枫又显著高于枫香树,半枫荷的萌发速率指数与阿丁枫无显著差异,但均显著高于枫香树。总体而言,半枫荷与阿丁枫的最适萌发温度为20~30 ℃,最大萌发率分别为90%和80%,枫香树的最适萌发温度为20~25 ℃,最大萌发率为80%,也就是说,半枫荷与阿丁枫种子耐高温能力强于枫香树,半枫荷的最大萌芽率高于阿丁枫和枫香树。
2.4温度对3种金缕梅科植物种子萌发时间的影响
统计分析表明,温度对3种金缕梅科植物种子的萌发开始时间和萌发持续时间均具有显著的影响(图3)。随着温度的升高,3种植物种子的萌发开始时间和萌发持续时间均显著缩短。3种植物相比较,15 ℃下,半枫荷的萌发开始时间显著早于阿丁枫和枫香树,但萌发持续时间显著长于阿丁枫和枫香树,阿丁枫与枫香树的萌发开始时间和萌发持续时间均无显著差异;20 ℃下,3种金缕梅科植物种子的萌发开始时间和萌发持续时间均无显著差异;25 ℃下,3种植物种子的萌发开始时间无显著差异,半枫荷萌发持续时间显著小于阿丁枫和枫香树;30 ℃下,枫香树的萌发开始时间显著晚于半枫荷和阿丁枫,但三者的萌发持续时间无显著差异。
3 结论与讨论
温度对种子萌发的开始、潜能和萌发率均具有重要影响[13],是决定植物定植成功最关键的因素。3种金缕梅科植物种子在本试验设置的温度范围内(15~30 ℃)均能萌发,虽然3种植物15 ℃下的种子萌发率均显著低于20~30 ℃,但半枫荷与阿丁枫15 ℃下的萌发率均能到达70%以上,枫香树能达到50%以上,表明3种金缕梅科植物种子萌发的温度范围较广,而且范围相似。3种植物均为南方树种,我国南方地区春季温度一般在10~20 ℃之间,这种萌发机制确保了大部分种子在合适的季节萌发,增大了幼苗的存活机会。濒危植物半枫荷各温度下的最大萌发率均显著高于阿丁枫和枫香树,适温下(20~30 ℃)其萌发率能达90%左右,并且各温度下其萌发速率显著高于或与另外2个金缕梅科植物无显著差异,说明温度不是影响半枫荷种子萌发的重要因素。
萌发开始时间和萌发持续时间的长短反映了种子萌发的速度和难易程度[14]。研究结果表明,各试验温度下3种金缕梅科植物种子的萌发开始时间和持续时间均较短,并随着温度的升高,3种植物的种子萌发开始时间和萌发持续时间显著缩短。15 ℃和30 ℃下3种植物中半枫荷的种子萌发开始时间最短,分别为6 d和1 d,枫香树萌发开始时间最长,分别为8 d和2 d。15 ℃下半枫荷的萌发持续时间最长,为9~10 d,30 ℃下3种植物无显著差异,均为4~5 d。野生环境中种子集中在某一时段萌发,增大了多变环境造成幼苗大量死亡的风险[15],但这种萌发特性有利于人工种苗繁育时提高种苗整齐度。
研究表明,种子大小会影响种子的萌发速率和萌发百分率[16]。比较3种金缕梅科植物的种子形态特征发现,濒危植物半枫荷的种子最大,其千粒重显著大于阿丁枫与枫香树,本研究3种金缕梅科植物中大种子有更高萌发率和萌发速率、更早的萌发开始时间,但物种间相差不是很大。3种植物中枫香树的种子最轻,长宽比最大,具短翅,易飞散传播,而半枫荷与阿丁枫的种子无翅。野外调查发现半枫荷结果率低于阿丁枫与枫香树,败育的种子多,种子数量少,说明半枫荷对单个种子的投入大,也就是说半枫荷采取高千粒重、低生育率的K-对策。一般情况下,K-对策的物种通常幼苗数量较少,种群在受到强烈干扰时难以短期内恢复种群规模,容易造成濒危。
综上所述,3种金缕梅科植物种子萌发的温度范围较广,萌发率和萌发速度较快,萌发开始时间较早和持续时间较短,温度不是影响3种植物种子萌发的重要因素,但短时间内种子大量萌发易造成因环境条件的突然变化而引起物种局域性的灭绝。半枫荷采取高千粒重、低生育率的K-对策可能是其濒危的重要原因。
[1]陈国德,吴海霞.半枫荷的叶片活性成分测定[J].林业研究,2013,41(4):6-8.
[2]温利载,温晋强.枫香树生态特性及育苗技术[J].现代农业科技,2013(1):173,184.
[3]龙双畏,刘济祥,郑伟.优良园林绿化树种阿丁枫育苗技术研究[J].北方园艺,2009(5):199-201.
[4]李文,张小平,郝朝运,等.珍稀植物连香树(Cercidiphyllumjaponicum)的种子萌发特性[J].生态学报,2008,28(11):5 445-5 453.
[5]崔现亮,罗娅婷,毕廷菊,等.储藏和萌发温度对青藏高原东缘12种灌木种子萌发的影响[J].生态学杂志,2014,33(1):23-32.
[6]葛滢,常杰,岳春雷,等.杭州石荠种子萌发的生理生态学研究[J].植物生态学报,1998,22(2):171-177.
[7]陈辉,张霜,曹敏.光和温度对西双版纳地区先锋树种对叶榕种子萌发的影响[J].植物生态学报,2008,32(5):1 084-1 090.
[8]GREIPSSON S,DAVY A J.Seed mass and germination behaviour in populations of the dune-building grassLeymusarenarius[J].Ann Bot,1995,76:493-501.
[9]王玉峰.温度对植物种子萌发机制的影响[J].防护林科技,2015(6):76-78.
[10]YAJUAN Z,XUEJUN Y,CAROL C B,et al.Effects of amount and frequency of precipitation and sand burial on seed germination,seedling emergence and survival of the dune grassLeymussecalinusin semiarid China[J].Plant Soil,2014,374(1):399-409.
[11]GUL B,WEBER D J.Effects of salinity,light and temperature on germination in Allenrolfea occidentalis[J].Canad J Bot,1999,77:240-246.
[12]马姜明,占婷婷,莫祖英,等.漓江流域岩溶区檵木群落不同恢复阶段主要共有种生态位变化[J].西北植物学报,2012,32(12):2 530-2 536.
[13]FLORES J,BRIONES O.Plant life-form and germination in a Mexican inter-tropical desert:Effects of soil water potential and temperature[J].J Arid Environ,2001,47:485-497.
[14]李雄,尹欣,杨时海,等.温度对高山植物紫花针茅种子萌发特性的影响[J].植物分类与资源学报,2014,36(6):698-706.
[15]刘文杰,刘坤,张春辉,等.种子萌发的积温效应——以青藏高原东缘的12种菊科植物为例[J].植物生态学报,2011,35(7):751-758.
[16]王桔红,崔现亮,陈学林,等.中、旱生植物萌发特性及其与种子大小关系的比较研究[J].植物生态学报,2007,31(6):1 037-1 045.
Effects of Temperature on Seed Germination of Three Species ofHamamelidaceae
WANGManlian1,WENXiangying2,3,WEIXiao1,JIANGYunsheng1,TANGHui1
2016-05-28
广西科技攻关项目(编号:桂科重1355001-5-5);广西科技成果转化项目(编号:1346004-29);桂林市科技成果转化与应用项目(编号:20140124-2);桂林市科技攻关项目(编号:20130414);国际植物园保护联盟项目(编号:BGCI 30412)。
王满莲(1978—),女,硕士,副研究员,主要从事植物生理生态学研究;E-mail:Wangml1978@163.com。
唐 辉(1972—),男,学士,研究员,主要从事植物引种驯化工作;E-mail:th@gxib.cn。
10.16590/j.cnki.1001-4705.2016.10.079
S 792
A
1001-4705(2016)10-0079-05
