海岛棉抗枯萎病相关基因实时荧光定量PCR分析
2016-11-26杜荣光姚正培陈全家张杰杜丽丽苏秀娟曲延英
杜荣光,姚正培,陈全家,张杰,杜丽丽,苏秀娟,曲延英
(新疆农业大学农学院/农业生物技术重点实验室,乌鲁木齐 830052)
海岛棉抗枯萎病相关基因实时荧光定量PCR分析
杜荣光,姚正培,陈全家,张杰,杜丽丽,苏秀娟,曲延英
(新疆农业大学农学院/农业生物技术重点实验室,乌鲁木齐 830052)
【目的】以不同抗性海岛棉为材料,利用实时荧光定量PCR技术对11个棉花抗枯萎病相关基因进行表达量分析,为海岛棉抗枯萎病育种及抗病机理提供候选基因资源。【方法】选择海岛棉抗枯萎病材料06-146、易感材料新海14号,及以其为父母本杂交的RIL系高代材料10893(超抗)、10895(高抗)、10897(感病)、10796(易感)为实验材料,进行接菌处理。分别取接菌0、4、10、18、28和40 h后的幼苗下胚轴,提取mRNA并反转录。根据已知陆地棉抗枯萎病相关的EST序列和海岛棉抗病转录组测序筛选到的抗病相关基因序列设计引物,进行实时荧光定量 PCR 反应。根据各基因在不同材料中的表达值,分析各基因与海岛棉抗枯萎病间的关系。【结果】CFW3、CFW10、comp62810、comp71372四个基因在海岛棉抗枯萎病过程中可能起着比较重要的作用,尤其是利用抗病材料和感病材料转录组测序后得到的与抗病性有关的comp62810、comp71372基因。【结论】不同棉花品种对病菌侵染的灵敏程度,不同的品种的灵敏度不同,用以判断基因的作用。
海岛棉;枯萎病;抗性基因;实时荧光定量PCR
0 引 言
【研究意义】海岛棉(GossypiumbarbadenseL.)纤维是世界上最优良的棉纤维,是主要的纺织工业原料。新疆是我国唯一的海岛棉产区 ,优越的自然生态条件为新疆海岛棉的发展提供了得天独厚的优势。棉花枯萎病(Fusariumwilt)是影响棉花经济性状最主要的病害之一,在棉花生长前期,枯萎病可造成死苗,中后期则影响蕾铃生长发育直至脱落, 导致棉花产量严重下降, 纤维品质变劣。随着新疆海岛棉种植面积的逐渐扩大,各棉区常年连作,棉花相关的病虫害有呈逐年加剧的趋势,特别是落叶型枯萎病迅速蔓延,生产中又需丰产、优质、早熟,必须在不同生态条件下具备逆抗性的海岛棉品种。因此,开展海岛棉抗枯萎病相关基因筛选并验证其功效,对新疆海岛棉育种具有重要意义。【前人研究进展】棉花抗枯萎病方面在陆地棉中已有大量研究报道。海岛棉中有关枯萎病抗性研究却鲜见报道[5]。【本研究切入点】研究使用实时荧光定量PCR技术分析相关基因在抗病性不同的材料中的表达特性。【拟解决的关键问题】以海岛棉抗枯萎病材料06-146、易感材料新海14及以其为父母本杂交的RIL系高代材料10893(超抗)、10895(高抗)、10897(感病)、10796(易感)为实验材料,进行不同时间的接菌处理。利用实时荧光定量PCR技术对棉花抗枯萎病相关基因进行表达量分析,为筛选海岛棉枯萎病抗性基因提供依据,进而为海岛棉抗枯萎病分子机制提供依据。
1 材料与方法
1.1 材 料
根据在大田调查枯萎病情。并进行抗病性评价后获得抗、感病海岛棉材料,分别是06-146、新海14号及以06-146、新海14号为父母本杂交筛选的RIL(Recombinant Inbred Lines,重组自交系)高代自交系10893、10895、10897、10796,其中06-146、10893、10895为抗病材料,新海14号、10796、10897为感病材料。试验材料由新疆农业大学农学院提供。
1.2 方 法
1.2.1 试验设计
将收集的种子消毒处理后,在发芽盒内催芽,之后在事先准备好的盆内进行水培育苗,待幼苗长至第三片真叶时,进行接菌。接菌时温度在25℃左右,湿度在60%以上。光照培养16 h/d,暗培养8 h/d。浸染棉花幼苗的菌液浓度为1×106个/mL,处理2 d后感病棉苗表现出叶片萎蔫脱落的枯萎病发病症状。分别取接菌0、4、10、18、28和40 h之后的幼苗下胚轴,每个至少3个重复。液氮速冻后置于-80℃冰箱保存备用。
1.2.2 RNA提取与反转录
实验采用北京全式金RransZolTMPlant试剂盒提取RNA。由赛若公司提供的反转录试剂盒来进行反转录。
1.2.3 引物设计
通过查找国内外有关抗性基因方面的文献,在棉花EST文库中查找与棉花抗枯萎病抗病相关序列。筛选与防御相关(Defense Relate)、胁迫相关(Stress Relate)、抗病相关(Disease-related)、细胞壁结构(Cell war structure)、生长(Growth,photosynthesis)等有关的5段EST序列;以及通过转录组测序所获得的与抗病性有关的类黄酮合成相关(Flavonoids biosynthesis-related)、转录因子(Transcription factor)、棉酚合成相关(Gossypol biosynthesis-related)、谷胱甘肽s转移酶(Glutathione S-transferase)、糖基转移酶(Glycosyl transferase)、致病相关蛋白(Pathogenesis-related)等基因序列。设计荧光定量引物序列。表1
表1 使用的引物序列
Table 1 Primer sequences used in this study
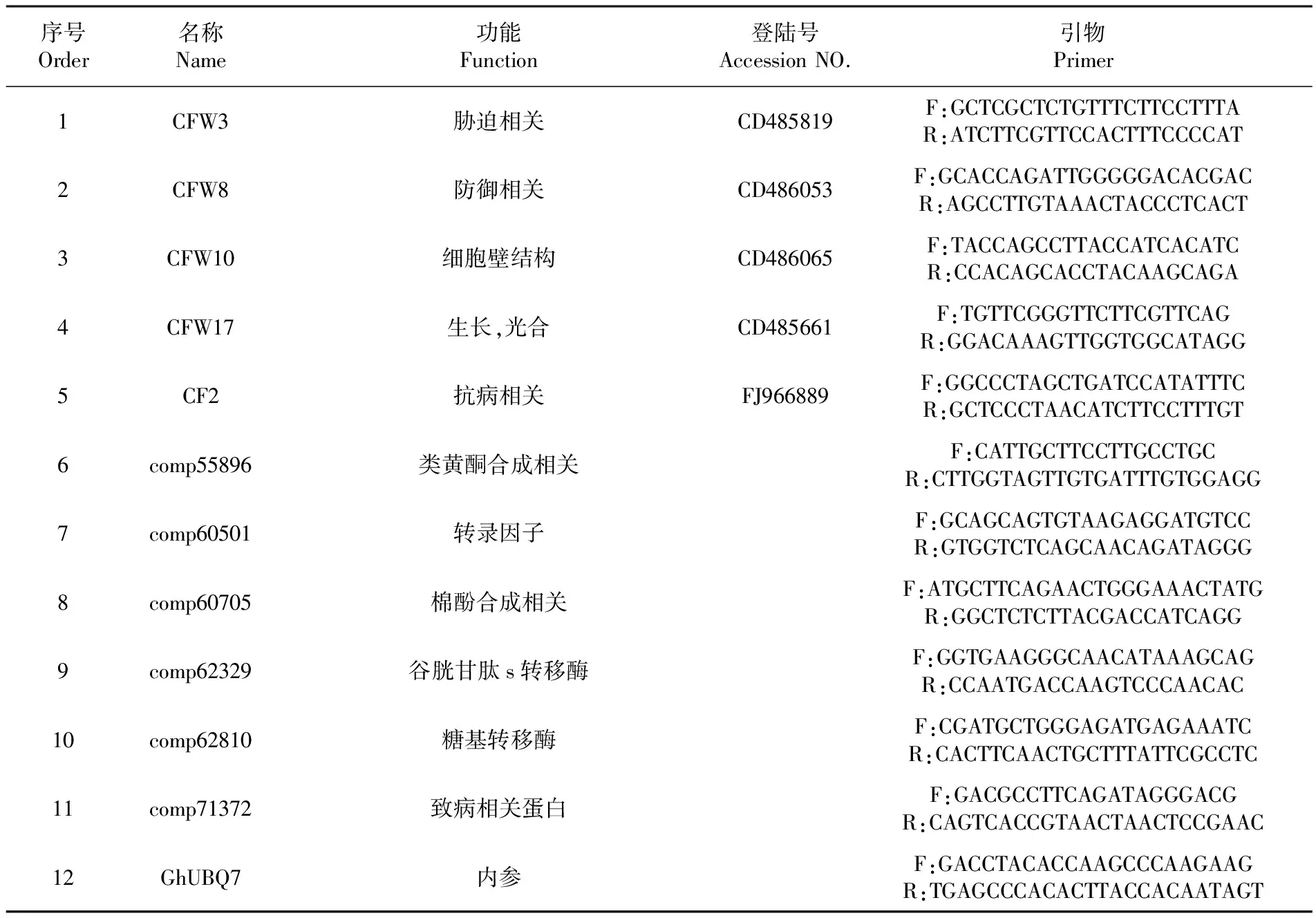
序号Order名称Name功能Function登陆号AccessionNO.引物Primer1CFW3胁迫相关CD485819F:GCTCGCTCTGTTTCTTCCTTTAR:ATCTTCGTTCCACTTTCCCCAT2CFW8防御相关CD486053F:GCACCAGATTGGGGGACACGACR:AGCCTTGTAAACTACCCTCACT3CFW10细胞壁结构CD486065F:TACCAGCCTTACCATCACATCR:CCACAGCACCTACAAGCAGA4CFW17生长,光合CD485661F:TGTTCGGGTTCTTCGTTCAGR:GGACAAAGTTGGTGGCATAGG5CF2抗病相关FJ966889F:GGCCCTAGCTGATCCATATTTCR:GCTCCCTAACATCTTCCTTTGT6comp55896类黄酮合成相关F:CATTGCTTCCTTGCCTGCR:CTTGGTAGTTGTGATTTGTGGAGG7comp60501转录因子F:GCAGCAGTGTAAGAGGATGTCCR:GTGGTCTCAGCAACAGATAGGG8comp60705棉酚合成相关F:ATGCTTCAGAACTGGGAAACTATGR:GGCTCTCTTACGACCATCAGG9comp62329谷胱甘肽s转移酶F:GGTGAAGGGCAACATAAAGCAGR:CCAATGACCAAGTCCCAACAC10comp62810糖基转移酶F:CGATGCTGGGAGATGAGAAATCR:CACTTCAACTGCTTTATTCGCCTC11comp71372致病相关蛋白F:GACGCCTTCAGATAGGGACGR:CAGTCACCGTAACTAACTCCGAAC12GhUBQ7内参F:GACCTACACCAAGCCCAAGAAGR:TGAGCCCACACTTACCACAATAGT
参数设置为:引物长度为20~25 bp,产物长度为100~250 bp,GC含量在40%~60%。荧光定量PCR引物由北京华大基因公司合成。
1.2.4 实时荧光 PCR 反应
分别在接菌0、4、10、18、28和40 h取抗病材料10893、10895、06-146和感病品种10897、10796、新海14号的下胚轴,提取总RNA,反转录为cDNA。进行实时荧光定量PCR分析。
荧光定量PCR反应体系(20 μL): SYBR Green super Mix 10 μL、引物各0.5 μL、 样本cDNA 1 μL、校正液1 μL、用ddH2O补至20 μL。在实验中,先进行95℃ 10 min,一个循环,进行预变性;之后再95℃ 30 s、60℃ 30 s、72℃ 30 s,40 个循环,是产物扩增的阶段。
1.3 数据统计
实时荧光定量反应结束后,通过相连的机器每个样品的荧光信号会自动收集,对溶解曲线的图来判断是否为同一个基因,剔除无用数据。利用SPSS算出标准误,根据其Ct值计算出相对表达量,计算公式如下:
目的基因相对表达量= 2-△△Ct
△△Ct=[(Ct目的基因-Ct内参基因)处理组-(Ct目的基因-Ct内参基因)对照组].
2 结果与分析
2.1 与棉花抗病相关EST序列荧光定量表达
研究表明,CFW3(胁迫相关)在4 h时,除10897和新海14号外其他材料表达量增加,在18 h时06-146和10893拥有最大值;新海14号、10796、10895和10897在28 h拥有最大值。随着时间延长,表达量在达到最高值后开始下降,但是抗病材料06-146和10893基因表达量在各阶段均高于感病材料新海14号和10897,感病材料在这个基因上相对于抗病材料在最大值有滞后现象。图1
研究表明,CFW8(防御相关)在接菌4 h之后,10796和父本(06-146)瞬时表达量增加明显,10893、10897和新海14号该基因在各时期表达量基本无变化。在18 h时06-146表达量最高,40 h时10786表达量很高,10895在28 h表达量达到最高,但是显著低于10796和06-146的表达量。图2
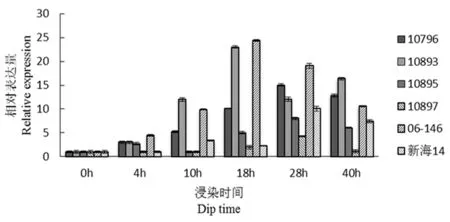
图1 CFW3基因荧光定量
Fig.1 Fluorescence quantitative diagram of CFW3
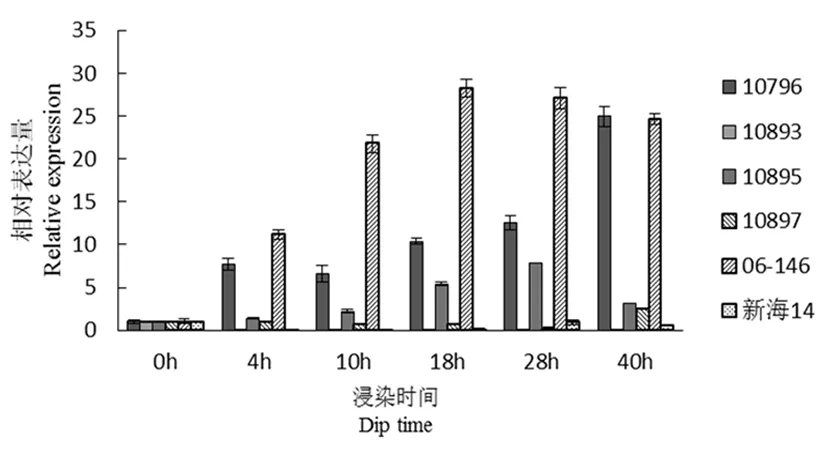
图2 CFW8基因荧光定量
Fig.2 Fluorescence quantitative diagram of CFW8
研究表明,CFW10(细胞壁结构相关)在接菌4 h之后,除新海14号外,其他材料表达量均增加,其中抗病材料06-146该基因表达量持续增高,10893、10895在10 h表达量达最高,之后表达量开始下降。感病材料10796中CFW10基因表达量也有一定程度升高,且在10 h达到最大量,但是其表达量显著低于其他抗病材料。感病材料10897该基因在4 h表达量增加后即下降。图3
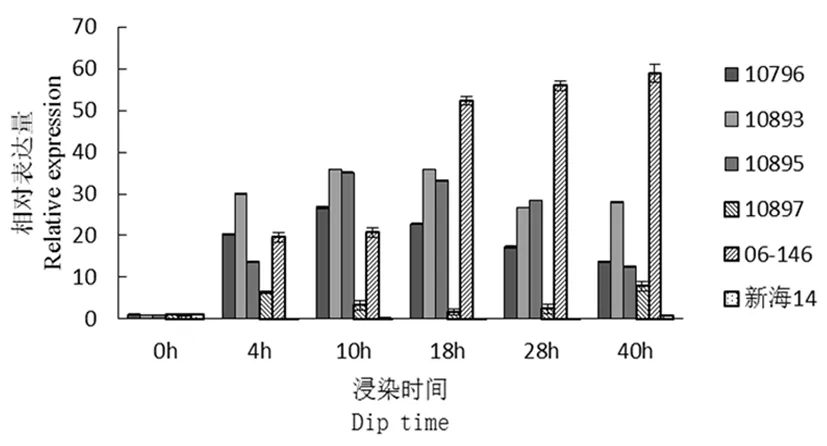
图3 CFW10基因荧光定量
Fig.3 Fluorescence quantitative diagram of CFW10
研究表明,CFW17(生长,光合相关)在接菌后,材料总体呈下降趋势,母本(新海14号)几乎不表达该基因,感病材料10796表达也很少,在这个基因在该感、抗材料的差异不是很明显,该基因在10897和06-146中表达量比较高。图4
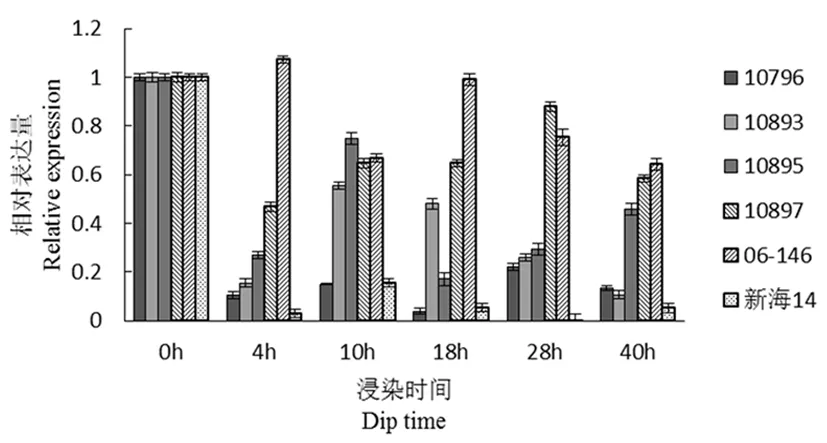
图4 CFW17基因荧光定量
Fig.4 Fluorescence quantitative diagram of CFW17
研究表明,CF2(抗病相关)在4 h时所有材料表达量有不同的增加,抗病材料06-146和感病材料10796在10 h是该基因表达量最高,且10796表达量高于06-146。在28 h时10895拥有最大表达量。该基因表达量在抗病材料和感病材料间没有明显规律性差异。图5
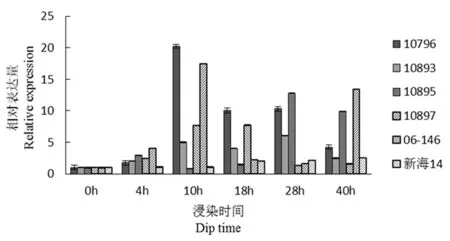
图5 CF2基因荧光定量
Fig.5 Fluorescence quantitative diagram of CF2
2.2 转录组测序获得与抗病相关基因的荧光定量表达
研究表明,comp55896(类黄酮合成相关)在4 h时,10796与父本(06-146)的相对表达量提升显著,在4 h时10796的相对表达量升到最高,之后表达量逐步下降。但在28 h时,10895和06-146表达量达到最大。而感病材料的相对表达量几乎被抑制。抗病材料与感病材料之间存在显著性差异。图6
研究表明,comp60501(转录因子基因)在接菌4 h时,10893与父本的瞬时表达增加明显,28 h时,表达量最高。抗病品种是上调表达。其他感病品种在接菌40 h之前表达量被抑制,在40 h表达量增高。图7
研究表明,comp60705(棉酚合成相关基因)在4 h时,除新海14号外,所有品种的表达量均有升高,抗病材料10893和父本(06-146)、感病材料10796在18 h时的表达量最大,之后随着时间推移,表达量逐渐下降。其中以10796表达量最高。图8
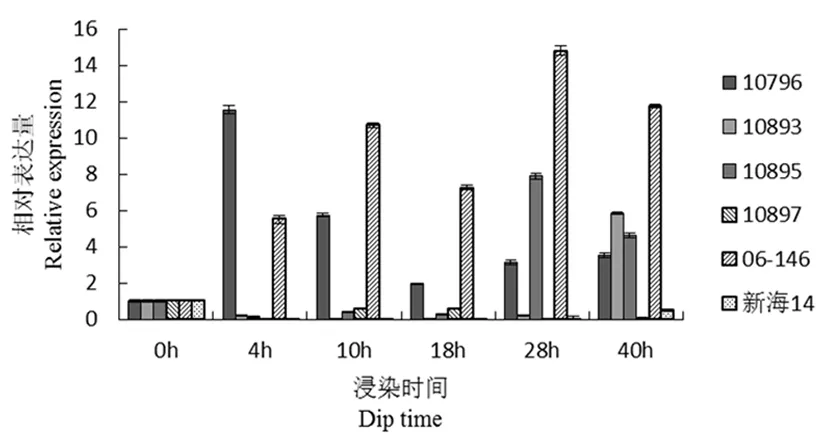
图6 comp55896基因荧光定量
Fig.6 Fluorescence quantitative diagram of comp55896
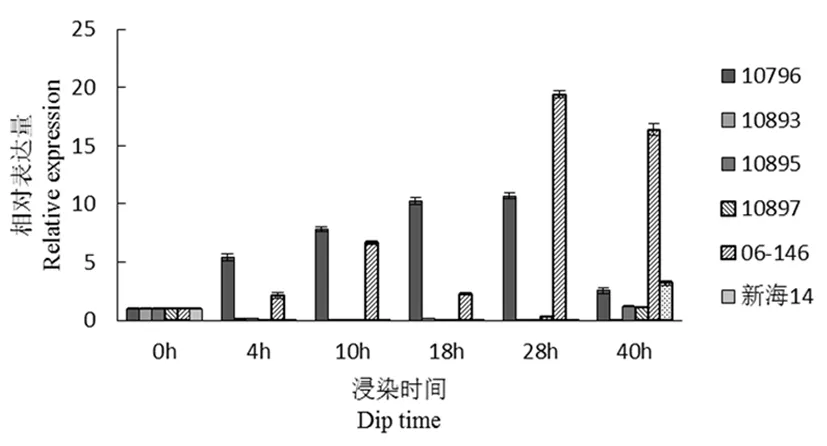
图7 comp60501基因荧光定量
Fig.7 Fluorescence quantitative diagram of comp60501
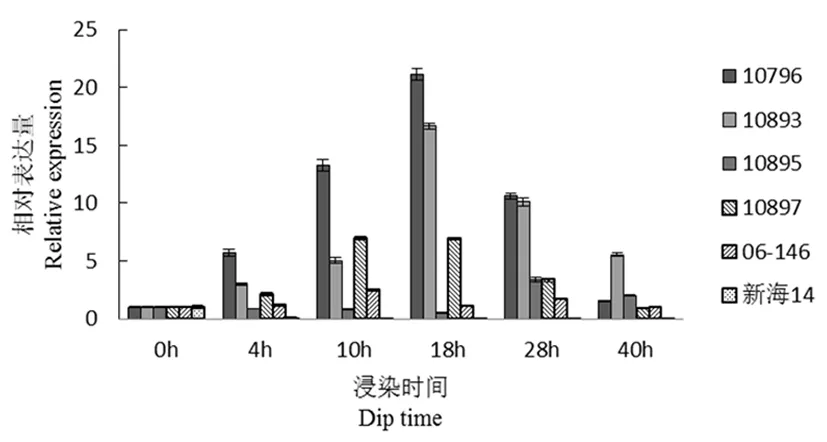
图8 comp60705基因荧光定量
Fig.8 Fluorescence quantitative diagram of comp60705
研究表明,comp62329(谷胱甘肽s转移酶)接菌4 h时10796、10895、10897、06-146的瞬时表达量增加,28 h时达到最大。抗病材料10893在各时间段该基因表达量均未增加。图9
研究表明,comp62810(糖基转移酶)在4 h是抗病品种10893与06-146表达量增加明显,感病品种10897、新海14号的表达量被抑制,且10893在10 h是表达量达到最高,其中母本新海14号的表达量先降低,后略有增高,在感、抗材料之间,该基因的表达量存在着很显著的差异。图10
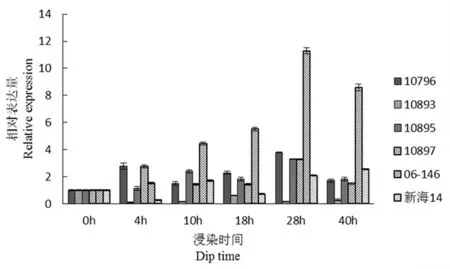
图9 comp62329基因荧光定量
Fig.9 Fluorescence quantitative diagram of comp62329
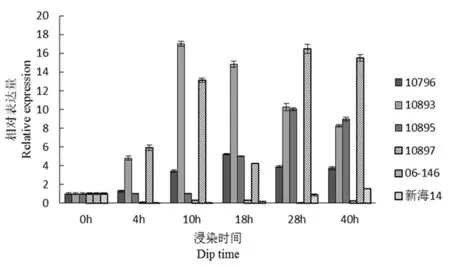
图10 comp62810基因荧光定量
Fig.10 Fluorescence quantitative diagram of comp62810
研究表明,comp71372(致病相关蛋白)在4 h时瞬时表达量增加,18 h是相对表达量达到最高,之后随着时间的增加,表达量递减。该基因在抗病材料中均表现上调表达,且表达量明显高于感病基因的,可能在海岛棉抗枯萎病中起着较重要的作用。图11
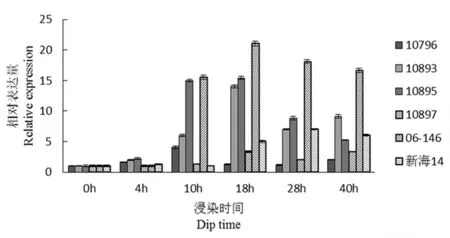
图11 comp71372基因荧光定量
Fig.11 Fluorescence quantitative diagram of comp71372
3 讨 论
植物抗病信号传导两种重要形式包括系统本身的抗性和诱导获得的抗性,植物会在不同的刺激下进行表达调控来防卫病原物。根据研究表达量的变化可知:CFW3、CFW10、comp62810、comp71372等抗病相关基因,在枯萎病病菌侵染的过程中,感、抗材料之间表达有差异,其差异主要体现在:相对于感病材料,抗病材料的最大表达量出现的时间比感病材料早,表达量远高于感病材料的,所以CFW3、CFW10、comp62810、comp71372这四个基因在抗病过程中可能起着比较重要的作用,尤其是利用抗病材料和感病材料转录组测序后得到的与抗病性有关的comp62810、comp71372基因。
CFW17在海岛棉棉花枯萎病侵染过程中随着时间的推移,表达量会减少,推断在受到病菌胁迫时植物光合能力和生长会受到影响。comp60501和comp62329基因在抗病材料中的表达量会比感病材料的多,最大表达量出现的时间,感、抗材料基本一致,推测这些基因对抗病有一定影响,但是对病菌的反应不如CFW3、CFW10、comp62810、comp71372基因敏感。comp60705和CF2两基因表现出在感病材料10796中表达量高于抗病材料的,可能暗示海岛棉抗病性与棉酚合成有关基因的表达有负相关,CF2由于是从陆地棉抗病相关EST库中筛选到的,可能暗示陆地棉和海岛棉枯萎病抗性基因的作用不一致。
CFW3、CFW10、comp62810、comp71372在海岛棉中抗病材料4 h表达量增加明显,且感病材料表达被抑制或者滞后表达,推测抗病材料对枯萎病的抗性是瞬时表达的,说明抗病材料对枯萎病菌的侵染灵敏,但在CFW8、comp55896中海岛棉抗病品种表达结果比较滞后。
利用抗病材料06-146和新海14号获得的转录组数据,在基因表达量上与抗病性的关系更为密切。comp60501转录因子中,抗病材料表现出非常高的相对表达量。comp60501转录因子调控植物的抗逆性,当植物受到侵染时,通过内部的反应,磷酸化级联反应刺激转录因子,让抗逆基因进行表达,由此提高植物抗性。所以,抗病品种的反应很快,这有利于植物进行阻止病菌的侵染,而感病材料的表达量不高,可能在病菌侵染时改转录因子并不能调控这类材料的抗病相关基因。comp62329中,谷胱甘肽s转移酶的作用是催化内源性或外源性有害物质的偶联,增加疏水性,将有害物质排出。接菌4 h时,10796、10895、10897、06-146的瞬时表达量增加,在28 h时达到最大,说明该类基因在海岛棉抗病中可能起到一定作用。comp62810中,接菌4 h时,表达量在抗病材料中极为明显。抗病材料的表达量基本是感病材料的4倍表达量,同时Comp71372基因在抗病或者是感病材料在10或18 h都有达到最高表达量的,但是抗病材料整体的表达量要高于感病材料,说明在海岛棉中这两个基因与海岛棉康枯萎病行关系密切。
4 结 论
选用的抗病材料10893、10895和感病材料10897、10796是由06-146与新海14号杂交后得到的重组自交系,经过两年两点的病田种植,抗病性鉴定筛选到的抗病、感病材料。其中亲本06-146是抗病材料,新海14号是感病材料。Comp基因是利用06-146和新海14号在枯萎病侵染不同时间,取下胚轴提取RNA后进行转录组测序得到的与抗病有关的基因序列。
棉花枯萎病是根部侵染的系统性病害,在接菌48 h内,是病菌侵染和植株体内受体识别病原物侵染信号并做出防卫的关键时期。所以要在40 h之内采样。与未接菌的(0 h)材料进行实时荧光定量Ct值的比较,观察抗病相关基因在抗、感材料之间瞬时表达量不同,推测基因在当时的上调或下调。对棉花下胚轴组织进行病毒诱导表达特性,不同棉花品种对病菌侵染的灵敏程度,不同的品种的灵敏度不同,用以判断基因的作用。
References)
[1] 袁继红.实时荧光定量 PCR 技术的实验研究[J].现代农业科技,2010,(13):20-22.
YUAN Ji-hong. (2010). Experimental Research on Real-time Fluorescent Quantitative PCR [J].ModernAgriculturalSciencesandTechnology, (13):20-22.(in Chinese)
[2] 赵利新,陆朝晖,戴俊生.借鉴国外经验促进新疆长绒棉生产发展[J].新疆农业科学,2005,42(6):438-440.
ZHAO Li-xin, LU Zhao-hui, DAI Jun-sheng. (2005). Promoting the Development of Long-fibre Cotton in Xinjiang by the Experience Abroad [J].XinjiangAgriculturalSciences, 42(6):438-440. (in Chinese)
[3] 杜丽丽.海岛棉枯萎病抗性遗传分析及抗病相关标记的评价[D].乌鲁木齐: 新疆农业大学硕士论文,2015.
DU Li-li.(2015). Genetic Analysis of Resistance toFusariumwiltand Evaluation of Resistance-related Markers in Gossypium barbadense [D]. Master Dissertation. Xinjiang Agriculture University, Urumqi. (in Chinese)
[4]肖蕊,余真真,Elsharawy AA,等. 土壤中棉花黄萎病菌SYBR Green I荧光 RT-PCR 定量检测技术研究[J].菌物学报,2011,30(4):598-603.
XIAO Rui, YU Zhen-zhen, Elsharawy, A.A., et al. (2011). SYBR Green I real time RT-PCR assay for quantitatively detecting the occurrence ofVerticilliumdahliae of cotton in naturally infested soil [J].Mycosystema, 30(4):598-603. (in Chinese)
[5]赵蕊,吕利华,陈婷,等.木尔坦棉花曲叶病毒 SYBR Green I实时荧光定量PCR检测方法[J].华南农业大学学报,2015,36(6):87-90.
ZHAO Rui, LV Li-hua, CHEN Ting, et al. (2015). A SYBR Green I real-time fluorescence quantitative PCR detection method for Cotton leaf curl virus [J].JournalofSouthChinaAgriculturalUniversity, 36(6):87-90. (in Chinese)
[6]沈元劼,齐谢敏,刘标,等.棉花黄萎病菌实时荧光定量PCR检测方法的建立及应用[J].生态学杂志,2015,34(7):2 058-2 063.
SHEN Yuan-jie, QI Xie-min, LIU Biao, et al. (2015). Development and application of a real-time PCR assay for the detection and quantification ofVerticilliumdahlia[J].ChineseJournalofEcology, 34(7):2,058-2,063. (in Chinese)
[7]袁伟,万红建,杨悦俭.植物实时荧光定量PCR内参基因的特点及选择[J].植物学报,2012,47(4):427-436.
YUAN Wei, WAN Hong-jiang, YANG Rui-jian.(2012). Plant Gene real-time PCR internal control characteristics and selection [J].ChineseBulletinofBotany, 47(4):427-436. (in Chinese)
[8]李瑞琴,刘星,邱慧珍,等.发生马铃薯立枯病土壤中立枯丝核菌的荧光定量PCR快速检测[J].草业学报,2013,22(5):136-144.
LI Rui-qin, LIU Xin, QIU Hui-zhen, et al. (2013). Quantitative PCR potato blight Rhizoctonia solani rapid detection [J].ActaPrataculturaeSinica, 22(5):136-144. (in Chinese)
[9] Ponchel, F., Toomes, C., Bransfield, K., Leong, F. T., Douglas, S. H., & Field, S. L., et al. (2003). Real-time pcr based on sybr-green i fluorescence: an alternative to the taqman assay for a relative quantification of gene rearrangements, gene amplifications and micro gene deletions.BmcBiotechnology, 3(1):18.
[10]陈旭,齐凤坤,康立功,等.实时荧光定量PCR技术研究进展及其应用[J].东北农业大学学报,2010,41(8):148-155.
CHEN Xu, QI Feng-kun, KANG Li-gong, et al. (2010).Advance and application of real-time fluorescent quantitative PCR [J].JournalofNortheastAgriculturalUniversity, 41(8):148-155. (in Chinese)
[11]汪秀秀,杨捷琳,宋青,等.转基因棉花GHB119品系特异性定量PCR检测方法的建立[J].农业生物技术学报,2014,22(3):380-388.
WANG Xiu-xiu, YANG Jie-lin, SONG Qing, et al. (2014). Establishment of a Novel Event-specific Quantitative PCR Method for Genetically Modified Cotton (Gossypiumsp.) GHB119 Detection [J].JournalofAgriculturalBiotechnology, 22(3):380-388. (in Chinese)
[12]赵爽,罗佳,凌宁,等.基因宏阵列和荧光定量PCR方法对西瓜枯萎病害土壤中尖孢镰刀菌的快速检测和定量[J].土壤学报,2010,47(4):703-708.
ZHAO Shuang, LUO Jia, LING Ning, et al. (2010). Quick Check and Quantification of Fusarium Oxysporum in Soil with Macroarray and Real-time PCR Method [J].ActaPedologicaSinica, 47(4):703-708. (in Chinese)
[13] 朱永军,张西英,李金荣,等.海岛棉枯萎病抗性遗传规律及分子标记的初步筛选[J].新疆农业科学,2010,47(2):268-273.
ZHU Yong-jun, ZHANG Xi-ying, LI Jin-rong, et al. (2010). Inheritance of Resistance toFusariumwiltand their Molecular Marker in Gossypium barbadense [J].XinjiangAgriculturalSciences, 47(2):268-273. (in Chinese)
[14] 武维华. 植物生理学[M].北京:科学出版社,2003:440-448.
WU Wei-hua. (2003).PlantPhsiology[M]. Beijing: Science Press: 440-448. (in Chinese)
Fund project:Supported by National Natural Science Foundation of China(31260361)
A Real-time PCR of Related Marker Gene in Cotton (GossypiumbarbadenseL.)
DU Rong-guang, YAO Zheng-pei, CHEN Quan-jia, ZHANG Jie, DU Li-li,SU Xiu-juan, QU Yan-ying
(College of Agronomy, Xinjiang Agricultural University /Laboratory of Agricultural BiotechnologyofXinjiangAgriculturalUniversity,Urumqi830052,China)
【Objective】 With different resistant island cottons as material, using real-time fluorescence quantitative PCR technology to analyze the expression levels of 11 genes related to wilt resistance in cotton, this project aims to provide candidate gene resources for breeding resistance varieties toFusariumwiltof sea island cotton and its mechanism.【Method】Sea Island cotton resistance toFusariumwiltdisease 06-146, the susceptible materials of Xinhai 14 and the parents of the hybrid RIL high generation materials, 10893 (super resistant), 10895 (supper resistant), 10879 (sensible) and 10796 (susceptible) were chosen as the experimental materials and at the same time, inoculation treatment was carried out. The seedlings of 0 h, 4 h, 10 h, 18 h, h, 28, 40 h, respectively, were taken to extract the mRNA and went through reverse transcription. Based on the known EST sequences ofFusariumwilt, resistance in upland cotton and the resistance related gene sequences screened from the Sea Island cotton disease resistance gene sequences, the real-time fluorescence quantitative PCR reaction was carried out. According to the expression of different genes in different materials, the relationship between different genes and the resistance toFusariumwiltof cotton were analyzed.【Result】Four genes, CFW3, CFW10, comp62810, comp71372 in cotton may play a more important role in the process of fighting againstFusariumwiltof Sea Island cotton. In particular, the comp62810 and comp71372 gene, which were obtained from the transcriptional group of the disease resistant material and the susceptible material, were related to the disease resistance.【Conclusion】Through the analysis, it is concluded that the sensitivity of different cotton varieties to the infection of the pathogen is different, which can be applied to determine the roles of the genes.
cotton (GossypiumbarbadenseL.);Fusariumwilt; real-time PCR
10.6048/j.issn.1001-4330.2016.11.003
2016-05-18
国家自然科学基金项目(31260361)
杜荣光(1991- ),男,新疆博乐人,硕士研究生,研究方向为棉花抗病,(E-mail)89251760@qq.com
曲延英(1962- ),女,山东人,博士,教授,研究方向为棉花遗传育种,(E-mail)xjyyq5322@126.com
S562
A
1001-4330(2016)11-1980-08
