产β-葡萄糖苷酶酵母菌的分离鉴定及酶学特性
2016-11-21徐亚男史学伟安洋洋谈思维童军茂
徐亚男,史学伟, 安洋洋, 谈思维,肖 婧,童军茂*
(1.石河子大学 食品学院,新疆 石河子832000;2.包头职业技术学院 经管系,内蒙古 包头010043)
产β-葡萄糖苷酶酵母菌的分离鉴定及酶学特性
徐亚男1,史学伟1, 安洋洋2, 谈思维1,肖 婧1,童军茂*1
(1.石河子大学 食品学院,新疆 石河子832000;2.包头职业技术学院 经管系,内蒙古 包头010043)
在葡萄酒自然发酵的初期,非酿酒酵母菌占主导地位,其产生的β-葡萄糖苷酶独特的水解活性,能够赋予葡萄酒特殊的香气。在新疆赤霞珠葡萄酒发酵过程中,通过对非酿酒酵母菌的采集与筛选,得到1株产β-葡萄糖苷酶能力较强的菌株;紫外-微波复合诱变后,酶活力增加1.81倍;经26S rRNA基因序列分析,鉴定为克鲁维毕赤酵母,并命名为XYN086(P.kluyveri XYN086);酶学性质研究表明:该菌株所产β-葡萄糖苷酶最适pH为6.0,在pH 6.0~8.0时酶活力保持60%以上;酶的最适作用温度为60℃,在60℃酶活力保持较好;金属离子依赖性实验表明,Fe2+和Cu2+对该菌株所产β-葡萄糖苷酶的酶活力均有激活作用,Mg2+、Ca2+、K+和Na+均有抑制作用,说明此菌株所产的β-葡萄糖苷酶对金属离子具有依赖性。据以上数据确认,该菌株为产β-葡萄糖苷酶克鲁维毕赤酵母。
β-葡萄糖苷酶;非酿酒酵母菌;酶学特性
β-葡萄糖苷酶在生物能源、食品工业、保健品、农业、医药等领域有着广泛的应用。β-葡萄糖苷酶属纤维素酶类,在生物能源领域,纤维素发酵生产燃料乙醇等过程中,添加一定的β-葡萄糖苷酶或产β-葡萄糖苷酶菌株,可提高纤维素降解的酶反应速率,为生物能源的利用与开发奠定了基础;在食品工业领域,β-葡萄糖苷酶作用于水果中的芳香前体物质,利于果汁脱苦和果汁增香,并可用作食品添加剂;在保健品行业中,β-葡萄糖苷酶应用于大豆异黄酮等,可有效增加产量和降低生产成本,其工业化应用前景较高;在农业领域,生氰β-葡萄糖苷酶在植物病虫害的绿色防治上具有重大意义;在医药领域,β-葡萄糖苷酶可应用于某些疾病的诊断和治疗。
非酿酒酵母菌是一类自然存在于葡萄表皮、酿酒环境中,能分泌多种胞外酶,通过代谢和自溶提高发酵食品的感官特性,参与复杂新鲜的风味物质形成的酵母菌[1]。葡萄酒的主要香气成分取决于葡萄中挥发性次级代谢产物的数量和化学性质[2]。其中,以自由分子或非芳香糖基化前体物质存在的烷烃贡献最大[3]。β-葡萄糖苷酶能够显著增加葡萄酒中非芳香糖基化前体物质的水解和芳香物质的释放[4],在葡萄酒的香气形成过程中起重要作用。在葡萄酒自然发酵过程的初期,非酿酒酵母菌占主导地位,其产生的β-葡萄糖苷酶独特的水解活性,能够赋予葡萄酒特殊的香气[5-6]。但是,目前已知菌株产β-葡萄糖苷酶的酶活力普遍比较低,在一定程度上限制了β-葡萄糖苷酶的工业化生产。因此,直接从自然界中筛选出产β-葡萄糖苷酶且酶活较高的菌株是最直接并且最有效的方法。
在新疆赤霞珠葡萄酒发酵过程中,经鉴别培养基采集非酿酒酵母菌,得到一株能分泌β-葡萄糖苷酶的野生菌株,通过分子生物学鉴定为克鲁维毕赤酵母;并对其酶学特性进行初步研究,证实其在食品工业领域有着良好的应用前景,进一步应用于新疆特色酿造发酵行业中,以期改善产品质量,提高设备利用率,同时为后期的研究提供理论基础。
1 材料与方法
1.1 材料
1.1.1 试验葡萄 赤霞珠葡萄,采自新疆葡萄园。
1.1.2 培养基 酵母浸出粉胨葡萄糖培养基(g/dL):葡萄糖2.0,酵母提取物 2.0,蛋白胨2.0;pH 7.0,121℃灭菌20 min。赖氨酸培养基[7](g/dL):赖氨酸0.56,葡萄糖1.0,KH2PO40.1,MgSO40.05,琼脂2.0;121℃灭菌20 min。筛选培养基(g/dL):葡萄糖2.0,蛋白胨2.0,酵母浸粉1.0,p-NPG 1.0;自然pH值,121℃灭菌20 min。发酵培养基(g/dL):葡萄糖 2.0,蛋白胨 2.0,酵母浸粉 1.0,NH4NO30.3,KH2PO40.4;自然pH值,121℃灭菌20 min,在温度降到60~70℃左右加入0.1 g/dL p-NPG。
1.1.3 仪器 Seven Excellence S400型pH计,梅特勒托利仪器(上海)有限公司制造;SYQ-DSX-280B手提式不锈钢电热蒸汽灭菌器,上海申安医疗器械厂制造;Mini-14K高速离心机,上海昨非实验室设备有限公司制造;数显气浴恒温振荡器,金坛市精达仪器制造厂制造;7200型可见分光光度计,尤尼柯(上海)仪器有限公司制造;Power Cycler Gradient SL PCR仪,德国耶拿分析仪器股份公司制造;IEFSYS琼脂糖凝胶电泳仪,英国Biochrom有限公司制造。
1.2 方法
1.2.1 菌株分离纯化 葡萄样品经破碎后带皮进行自然发酵,发酵启动后在整个发酵过程中定期取样,以不同浓度梯度涂布于赖氨酸培养基,28℃培养48 h,划线分离;重复多次直至得到单菌落后,接种于酵母浸出粉胨葡萄糖培养基,28℃培养48 h后,保存于-80℃冰箱。
1.2.2 产酶菌株的筛选 取试验菌株的菌悬液0.1 mL,以10倍系列稀释至10-1~10-6,分别取各个梯度稀释液1 mL接种于筛选培养基上,培养72 h后,喷1 mol/L碳酸钠,选择其中显示黄色光圈的菌株作为初筛菌株;初筛菌株接种于发酵培养基,置于温度30℃的摇床,150 r/min培养72 h后,发酵液于8 000 r/min离心10 min,除菌,收集上清液,于紫外分光光度计400 nm下测定吸光值;筛选酶活力大的菌株,备用。
1.2.3 产酶菌株诱变选育 将筛选出的菌株发酵液在离心机中5 000 r/min离心8 min,弃去上清液,用无菌水重新悬浮;吸取2 mL放置于4℃冰箱中作为对照;剩余的菌液,吸取5 mL加入无菌平板中,平板置于紫外灯已照射30 min的磁力搅拌器上,等距离放置距灯30 cm处,照射20 min,得诱变菌株1;诱变菌株1放入微波源为2 450 MHz、850 W的微波炉中,每照射10 s,冷却10 s后再照射10 s,冷却后在37℃培养60 h,得到最终诱变菌株。
1.2.4 粗酶液的提取和 β-葡萄糖苷酶活性测定
取诱变菌株和对照菌株发酵液于4℃8 000 r/min离心10 min,上清液即为粗酶液。取0.05 mL酶液,将50 mL 0.2 g/dL p-NPG溶液,0.15 mL pH值为5.0的醋酸缓冲液混合后,30℃下水浴预热10 min后立即加入碳酸钠中止反应。室温放置5 min,400 nm处测吸光值,同时做空白样品;酶活力单位(IU)定义:每分钟生成1 μmol对硝基苯酚用酶量为1个酶活单位。酶活力计算公式:
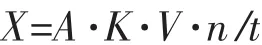
式中:X为酶活力 (IU/mL);A为吸光值;K为吸光常数;V为反应总体积;n为稀释倍数;t为反应时间。
1.2.5 菌株的分子生物学鉴定 采用酵母基因组DNA提取试剂盒提取试验菌株DNA,以所得基因组DNA为模板,利用ITS1和ITS4为引物扩增ITS区的基因,PCR反应体系参照文献[8],将扩增产物送到北京三博远志生物技术有限责任公司进行测序;将ITS序列输入NCBI(美国国家生物信息中心)的GenBank数据库中进行比对,分析序列同源性,确定该菌株的分类地位。
1.2.6 粗酶性质研究
1)pH对酶活的影响:分别配制pH为3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0的柠檬酸缓冲溶液,测定酶活力,以最高点酶活为100%,其他与之相比得相对酶活,绘制酶最适pH值曲线;
2)pH稳定性研究:粗酶液分别与等量不同pH缓冲液混匀,置于30℃下水浴保温1 h后,测定剩余酶活,以最高点酶活为100%,其他pH条件下与之相比为相对酶活,绘制酶的pH值稳定性曲线;
3)温度对酶活的影响:在温度分别为30、40、50、60、70℃下测酶活,以最高点酶活为100%,其他与之相比得相对酶活,以确定最适反应温度,绘制酶催化温度曲线;
4)热稳定性研究:粗酶液置于50、60、70℃不同温度下保温一定时间,取出后迅速冷却,以pH为7.0 2 g/dL葡萄糖溶液为底物溶液,在30℃下测定剩余酶活,以未经处理的酶溶液酶活为100%,其他温度条件下与之相比为相对酶活,绘制酶热稳定性曲线;
5)金属离子对酶活的影响:反应体系中加1 mL终浓度为 1 mmol/L的各种金属离子 Ca2+、Mg2+、Fe2+、Cu2+、Na+、K+,充分混匀后在30℃下放置24 h,测酶活,以1 mL双蒸水代替金属离子的酶溶液酶活力为100%,其他金属离子与之相比为相对酶活。
1.2.7 数据分析 独立重复进行3次试验,结果是3次测定的平均值±标准偏差,所有曲线以Origin 8作图。
2 结果与讨论
2.1 产酶菌株的筛选
以采集于发酵过程中的新鲜葡萄汁为分离源,分离得到具有水解β-葡萄糖苷酶活性的菌株3株,通过β-葡萄糖苷酶黄色光圈亮度试验,得到黄色光圈较明显的1号和2号菌株,结果如表1所示。
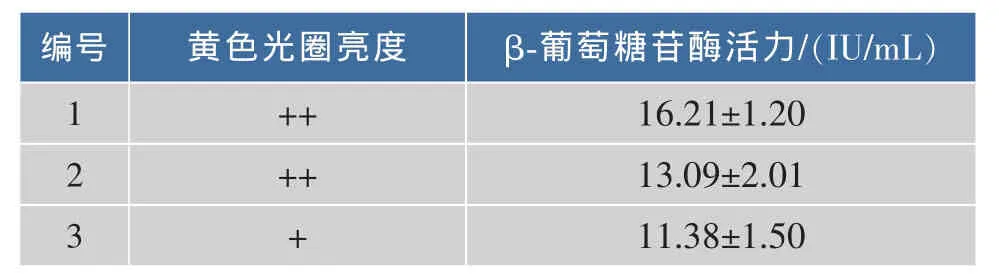
表1 产酶菌株筛选结果Table 1 Results of the screening strain
由表1可知,分离得到的3株菌株均产β-葡萄糖苷酶,通过黄色光圈亮度可知,1号和2号菌株水解能力最强,3号菌株次之;酶活测定结果表明,1号菌株产β-葡萄糖苷酶能力高于其他2株,2号菌株酶活力高于3号菌株,表明1号和2号菌株均具有可能的开发前景,故选择1号和2号菌株为本次试验初次筛选的菌株。
2.2 产酶菌株诱变选育
将产β-葡萄糖苷酶能力较好的菌株进行诱变选育,结果见表2。可知,各菌株经紫外-微波复合诱变后,酶活力均不同程度增大,1号菌株酶活力增加(13.12±1.45)IU/mL,2号菌株酶活力增加(5.05±0.29)IU/mL,经诱变后,1号菌株所产β-葡萄糖苷酶活力明显高于2号菌株,故本次试验所得菌株为1号诱变菌株。

表2 菌株诱变选育对酶活的影响Table 2 Effect of mutation on β-glucosidase activity
2.3 26s rRNA基因序列分析
对1号菌株进行DNA的提取和PCR扩增,通过分子生物学手段鉴定该菌株的ITS核苷酸序列如下:
GTCGCTACTACCGATTGAATGGCTTAGTGAGG CTCCAAGATTGGCGCCGCGGGAGGGGCAACTTTC CCATGGCGCTGAAAATTCAGTCAAACTTGGTCATT TAGAGGTCGTAAAAGTCGTAACAAGGTTTCCGTA GGTGAACCTGCGGAAGGATCATTACTGTGATTTAT ATCTTATACACATGCGTGAGCGCACCAAACACCTA AAATTGTAATACCACAGTCACTAAGTTTTAACAAA ACAAAACTTTCAACAACGGATCTCTTGGTTCTCGC ATCGATGAAGAGCGCAGCGAAATGCGATACCTAG TGTGAATTGCAGCCATCGTGAATCATCGAGTTCTT GAACGCACATTGCGCCCCATGGTATTCCATGGGG CATGCCTGTCTGAGCGTCGTTTCCTTCTTGCGCAA GCAGAGTTGAGAACAGGCTATGCCTTTTTCGAAAT GGAACGTCGTGGACGAAGTGAACTAAACTTTTAG CACGCTTTGGCCGCCGAACTTTTAACTAAGCTCGA CCTCAGATCAGGTAGGAATACCCGCTGAACTTAA GCATATCAATAAGCGGAGGAAAAGAAACCAACA GGGATTGCCTCAGTAGCGGCGAGTGAAGCGGCAA GAGCTCAGATTTGAAATCTCACCTAGTGTGCGAG
见表 3,1号菌株的 26s rRNA基因序列和BLAST比对结果显示,它与Pichia kluyveri strain 28FR菌的26s rRNA(KJ095611)有100%的相似度。构建的系统发育树 (图1)表明,1号菌株在Pichia属进化树一个分支上。因此,该菌株为克鲁维毕赤酵母,命名为XYN086(P.kluyveri XYN086)。

表3 1号菌株的26s rRNA基因测序BLAST结果Table 3 BLAST results of 26s r RNA gene sequencing of strain 1

图1 1号菌株(XYN086)基于26s r RNA基因发育树Fig.1 Development tree diagram based on 26s r RNAgenes of strain 1(XYN086)
2.4 酶的最适pH及其酸碱稳定性
测定不同pH对β-葡萄糖苷酶活性的影响,如图2所示,随着pH的增加,该菌株所产β-葡萄糖苷酶酶活力逐渐增大,在pH 6.0时达到最高,pH继续增加,酶活力降低较大幅度,故确定该菌株的产酶最适pH为6.0。
如图3所示,该菌株所产β-葡萄糖苷酶在pH为6.0时,酶活力保持最佳,在pH 6.0~8.0,可保持较好酶活力 (相对酶活大于60%),在强酸pH为3.0、强碱pH为10.0时,酶活损失较严重,损失率约为80%。说明该菌株所产β-葡萄糖苷酶在pH 6.0~8.0较稳定,利于β-葡萄糖苷酶的贮藏和生产。

图2 pH对β-葡萄糖苷酶活性的影响Fig.2 Effect of pH on β-glucosidase activity

图3 pH稳定性Fig.3 pH stability
2.5 酶的最适温度及其热稳定性
在不同温度下,测定β-葡萄糖苷酶的活性,由图4可知,随着温度的增加,β-葡萄糖苷酶活力增大,在60℃时,该菌株表现了最佳酶活力,在50~60℃各菌株均具有较高活性,当温度低于30℃或高于70℃时,酶活力显著降低。
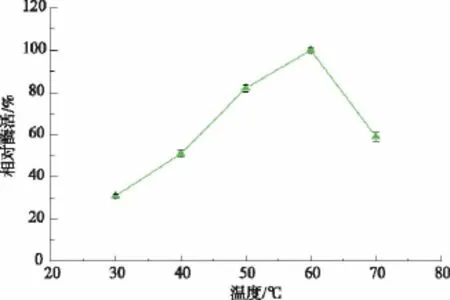
图4 温度对β-葡萄糖苷酶活性的影响Fig.4 Effects of temperature on β-glucosidase activity
如图5所示,随着温度的升高,经一段时间后,酶的活性逐渐降低;在60℃下,在不同时间内,测定的酶的活性相对稳定,保温60 min后,仍保留80%的活性;70℃下保温20 min后,酶活力损失30%,保温60 min后,剩余酶活力仅为21%。由此可知,该β-葡萄糖苷酶在60℃即最适温度下拥有良好的稳定性。
2.6 金属离子对蛋白酶活性的影响
如表4所示,Fe2+和Cu2+对该菌株所产β-葡萄糖苷酶的酶活力均有激活作用,Fe2+的激活作用不明显,Cu2+的激活作用最强,为 (136.92±2.27)%;Mg2+、Ca2+、K+和Na+对该菌株所产β-葡萄糖苷酶的酶活力均有抑制作用,K+的抑制作用最显著,为(43.51±1.21)%,说明该菌株所产的β-葡萄糖苷酶需要依赖金属离子维持其活性。

图5 热稳定性Fig.5 Thermal stability

表4 金属离子对酶活力的影响Table 4 Effects of metal ions on β-glucosidase activity
3 结语
在新疆赤霞珠葡萄酒发酵过程中,通过对非酿酒酵母菌的采集与筛选,得到1株产β-葡萄糖苷酶能力较强的菌株;紫外-微波复合诱变后,酶活力增加1.81倍;经26s rRNA基因序列分析,鉴定为克鲁维毕赤酵母,并命名为 XYN086(P.kluyveri XYN086);酶学性质研究表明:该菌株所产β-葡萄糖苷酶最适pH为6.0,在pH为6.0~8.0时酶活力保持在60%以上;酶的最适作用温度为60℃,在60℃酶活力保持较好;金属离子依赖性发现,Fe2+和Cu2+对该菌株所产β-葡萄糖苷酶的酶活力均有激活作用,Cu2+的激活作用最强,为 (136.92±2.27)%;Mg2+、Ca2+、K+和Na+均有抑制作用,K+的抑制作用最显著,为(43.51±1.21)%,说明此菌株所产的β-葡萄糖苷酶对金属离子具有依赖性。据以上数据确认,该菌株为产β-葡萄糖苷酶克鲁维毕赤酵母。
参考文献:
[1]SEGURA-García L E,TAILLANDIER P,BRANDAM C,et al.Fermentative capacity of Saccharomyces and non-Saccharomyces in agave juice and semi-synthetic medium[J].LWT-Food Science and Technology,2015,60(1):284-291.
[2]van Rensburg S J,BERMAN P A,POTOCNIK F C V,et al.Glycosylation of transferrin in Alzheimers disease and alcohol-induced dementia[J].Metabolic Brain Disease,2010,15(4):243-247.
[3]MARAI L,KUKSIS A.Simultaneous quantitation of krebs cycle and related acids by mass fragmentography[J].Journal of Chromatography(A),2013,268:447-460.
[4]RAPP A,MANDERY H.Wine aroma[J].Cellular and Molecular Life Sciences,2000(8):873-884.
[5]LAFFORT P,ETCHETO M,PATTE F.Implications of power law exponent in synergy and inhibition of olfactory mixtures[J]. Chemical Senses,2011,14(1):11-23.
[6]MULLER C J,FUGELSANG K C.Red wine but not white:the importance of fully characterizing wines used in health studies[J]. The American Journal of Clinical Nutrition,2000,66(2):447.
[7]FERNANDEZ R M,LAMY-FREUND M T.Correlation between the effects of a cationic peptide on the hydration and fluidity of anionic lipid bilayers:a comparative study with sodium ions and cholesterol[J].Biophysical Chemistry,2000,87(2):87-102.
[8]MANZANARES P,ROJAS V.A preliminary search for anthocyanin-beta-D-glucosidase activity in non-Saccharomyces wine yeasts[J].International Journal of Food Science and Technology,2000,35(1):95.
科技信息
韩国修订糖、碳水化合物、脂肪、维生素D、铬等的每日营养成分标准值
2016年6月16日,据韩媒报道,韩国食品和药品安全部于15日发布了《食品等的标示标准》部分修改告示(案)行政预告。
糖类的每日营养成分标准值规定为100 g,并对碳水化合物、脂肪、维生素D、铬等的标准值进行了调整。碳水化合物由330 g下调至324 g,脂肪由51 g上调至54 g,维生素D由5 μg上调至10 μg,铬由50 μg减至30 μg。
另外,为使以婴幼儿、孕妇、哺乳期妇女等特定人群为对象的食品可以该类人群的推荐摄入量或充分摄入量为标准进行标示,对 “韩国人营养素摄取标准表”进行修改。
根据糖类标准值的制定,对于糖类,要标示营养成分标准值相关比例,营养标示表格图案中追加糖类的“1日营养成分标准值相关比例(%)”。
[信息来源]WTO检验检疫信息网.韩国修订糖、碳水化合物、脂肪、维生素D、铬等的每日营养成分标准值 [EB/OL].(2016-6-17).http://www.wtociq.gov.cn
Isolation,Identification and Enzymatic Characteristics of Yeasts Producing β-Glucosidase
XU Yanan1,SHI Xuewei1,AN Yangyang2,TAN Siwei1,XIAO Jing1,TONG Junmao1
(1.College of Food Science,Shihezi University,Shihezi 832000,China;2.Administered by the Department,Baotou Vocational and Technical College,Baotou 010043,China)
During the fermentation of wine under natural conditions,non-Saccharomyces yeasts predominate in the initial stage of fermentation,and they can produce the unique hydrolytic activity β-glucosidases which give the wine a special aroma.In this study,a strain with good capacity of producing β-glucosidase was isolated from mustby the collection and screening of non-Saccharomyces yeasts during the fermentation of Xinjiang Cabernet.The strain was indentified as Pichia kluyver and named P.kluyveri XYN086 by morphology and ITS sequence analysis.The studies on enzymatic properties showed that the optimum pH for the β-glucosidase was 6.0 and its activity remained at more than 60%in pH 6.0-8.0.Furthermore,the optimum temperature for this enzyme was 60℃ and its activity was kept better at this temperature.Metal ions has a great influence on the activity of β-glucosidase,the enzyme was activated by Fe2+and Cu2+,whereas it was inhibited by Mg2+,Ca2+,K+,and Na+.These results showed that XYN086 had the potentialdevelopment values of producing β-glucosidase.
β-glucosidase,non-Saccharomyces,enzymatic characteristics
Q 939.9
A
1673—1689(2016)010—1100—06
2015-01-07
新疆生产建设兵团科技攻关项目(P20136500002-1146;2015AB016,2016AB009);新疆生产建设兵团青年科技创新资金项目(DEC_PGUEIK_893341);新疆生产建设兵团科技攻关与成果转化项目(2016AD024)。
*通信作者:童军茂(1963—),男,山东招远人,工学博士,教授,主要从事果蔬贮藏与加工研究。E-mail:1730233122@qq.com
