北疆地区同域分布的沙拐枣属植物是否存在杂交初探
2016-11-10康晓珊索菲娅段士民潘伯荣张永智
康晓珊 索菲娅 段士民 潘伯荣 张永智 田 聪
(1.中国科学院新疆生态与地理研究所,中国科学院干旱区生物地理与生物资源重点实验室,乌鲁木齐 830011; 2.中国科学院新疆生态与地理研究所吐鲁番沙漠植物园,新疆吐鲁番 830083; 3.新疆大学生命科学与技术学院,乌鲁木齐 830046; 4.新疆畜牧科学院草业研究所,乌鲁木齐 830000)
北疆地区同域分布的沙拐枣属植物是否存在杂交初探
康晓珊1,2索菲娅3段士民1,2潘伯荣1,2张永智2田 聪4
(1.中国科学院新疆生态与地理研究所,中国科学院干旱区生物地理与生物资源重点实验室,乌鲁木齐 830011;2.中国科学院新疆生态与地理研究所吐鲁番沙漠植物园,新疆吐鲁番 830083;3.新疆大学生命科学与技术学院,乌鲁木齐 830046;4.新疆畜牧科学院草业研究所,乌鲁木齐 830000)
杂交问题一直是沙拐枣属植物分类的核心问题。本文通过对自然生境和植物园的开花物候观测,人工授粉杂交实验以及花粉管荧光显色实验来探讨同域分布的沙拐枣属植物是否存在杂交,结果显示:同域分布种间均存在花期重叠;人工杂交授粉坐果率极低,且无法正常萌发,花粉管荧光显微观察实验表明杂交花粉能在柱头上萌发,但是花粉管不能伸长到子房。综上实验结果表明,种间配子亲和性很低,既存在合子前隔离,又存在合子后生殖隔离,认为该属不存在杂交。开展此项研究对于了解同域分布沙拐枣属植物间的隔离分化具有重要意义;为该属不存在杂交提供有力证据,也为该属的分类研究开拓了新的思路。
沙拐枣属(CalligonumL.);开花物候;杂交;花粉管荧光显微
沙拐枣(CalligonumL.)属为蓼科(Polygonaceae)灌木或半灌木,广泛分布于亚洲,欧洲南部和非洲北部,以沙拐枣属植物为建群种或共建种组成的植物群落,是亚非荒漠区中一类很独特的植被,中国是该属分布的东部区域。在我国该属主要分布于新疆,内蒙古,甘肃,青海,宁夏等地区。全球共35种,中国有23种,其中新疆有22种[1~2]。
该属的分类主要依据果实特征,但是其果实形态即使在同一居群也展示出丰富的多态性,甚至在同株间也有差异[3~6],有些果实的形态被称为过渡形态,中国学者称之为果实形态的平行显现[7~8],认为是存在天然杂交[9~11],苏联学者将这类具有过渡形态果实的种,归入杂交种或异名种。在《苏联维管束植物》[12]一书中就记录了一些通过果实形态特征比较后被认为是杂交的种,但是究竟是否存在杂交,还没有进行相应的研究。因此,杂交问题一直是该属的核心问题,也是解决分类问题的关键所在。本文将重点放在分布区域重叠的种间进行研究。
沙拐枣属植物新疆分布有22个种,南疆分布的仅有5个种(英吉沙沙拐枣,库尔勒沙拐枣,塔里木沙拐枣,甘肃沙拐枣和若羌沙拐枣),其中17个种都分布在北疆地区,存在2~3个种同域分布的现象,因此北疆地区是研究沙拐枣属植物是否存在杂交的理想研究区域。
在查阅标本信息以及考察相关文献的基础上,课题组从2005~2014年多次进行野外考察,较为充分的了解了西北地区沙拐枣属植物的分类与分布,为本研究的样地选择和观测做好了充分的准备。
吐鲁番沙漠植物园(TEBG)从原产地引进13种沙拐枣的种子进行实生栽培已有近40年的历史了[13],沙拐枣属植物生长良好,植株均能正常发育并开花结实。在植物园内形成了一定规模的群落景观,也是进行实验研究的良好的场所。
沙拐枣属植物在花的颜色、形态方面都差异不大,部分种同域分布,花期相近或有重叠[14~15],这都为杂交提供了便利和可能。笔者曾针对吐鲁番沙漠植物园的沙拐枣属4种植物的繁殖生物学进行过前期研究,本文在前期研究基础上,将植物园和野外生境同域分布的研究相结合,通过对开花物候观测,人工授粉杂交实验以及花粉管荧光显色实验来探讨同域分布的沙拐枣属植物是否存在杂交或者生殖隔离,同时,开展此项研究对于了解同域分布沙拐枣属植物间的隔离分化具有重要意义,对该属的分类、果实多型[4]以及是否存在杂交提供新的思路和见解。
2 研究方法和内容
2.1 开花物候
在课题组2005至2014年多次野外考察的基础上,确定自然生境下同域分布的沙拐枣的观测样点(表1)。开花时期多次在植物园和野外生境定点观测,记录开花时间,考察同域分布种间是否存在花期重叠。
2.2 人工授粉杂交试验
在盛花期,每个种选取生长势基本一致的植株5~10株,在即将开花前标记,在对花期重叠的种进行杂交授粉前,先进行套袋,在花瓣闭合,花药未开裂,柱头表面干净无花粉时,用羊皮纸袋套住花朵(保证每个袋中只有一朵花)。授粉时,去掉羊皮纸袋,戴头戴式放大镜,用尖头镊子去掉全部雄蕊后,将预先采集好的父本花粉抖落在母本柱头上,授粉完成再次套上羊皮纸袋,每个杂交组合授粉花数目大于30朵(表2)。48 h后用剪刀剪开纸袋上端,每日观察记录果实生长情况,直至果实脱落后收集、统计。坐果率的统计方法为:
成熟结实数/授粉套袋数×100%
(1)
2.3 花粉管荧光显色实验
不同的杂交组合,每个组合处理10朵,分别于授粉套袋后36 h取回(36 h是经过对柱头可受性检测后得出的最佳时间),用FAA固定液固定,将其花柱取下在8 mol·L-1的NaOH溶液中软化15 min,用蒸馏水清洗后转入0.1%的苯胺蓝溶液染色30 min以上,压片,置Olympus BH-2型荧光显微镜下观察其自花粉在柱头上的萌发及花粉管萌发生长情况。
2.4 数据分析
利用SPSS18.0统计分析软件对所测数据进行处理分析。
3 结果
3.1 花期物候及分布区域
不同种间的同域分布的主要有如下六个地点(表1),同域分布的种间均存在或多或少的花期重叠,同时由于气候差异,植物园的物候和野外生境差异较大大。
3.2 套袋实验
7个同域分布种组间的杂交坐果率见表2。种间杂交坐果率极低,有极个别坐果的果实,也不能正常萌发。种间杂交坐果率低,说明种间配子亲和性很低。
3.3 花粉管荧光显色反应
从图1B看出,大量杂交花粉在柱头上置落,仅有部分花粉能够在柱头上萌发,生长出一段花粉管,但是所有照片中均未发现花粉管到达子房。种间的花粉能在柱头上萌发,但是花粉管不能到达子房,使得花粉与胚珠无法受精形成种子,因此认为存在合子前生殖隔离。结合套袋实验结果,认为种间隔离机制在合子后也存在。
表1自然生境和吐鲁番沙漠植物园沙拐枣属植物开花物候比较
Table1FloweringphenologicalofCalligonumL.speciesbetweennaturalhabitatsandTEBG
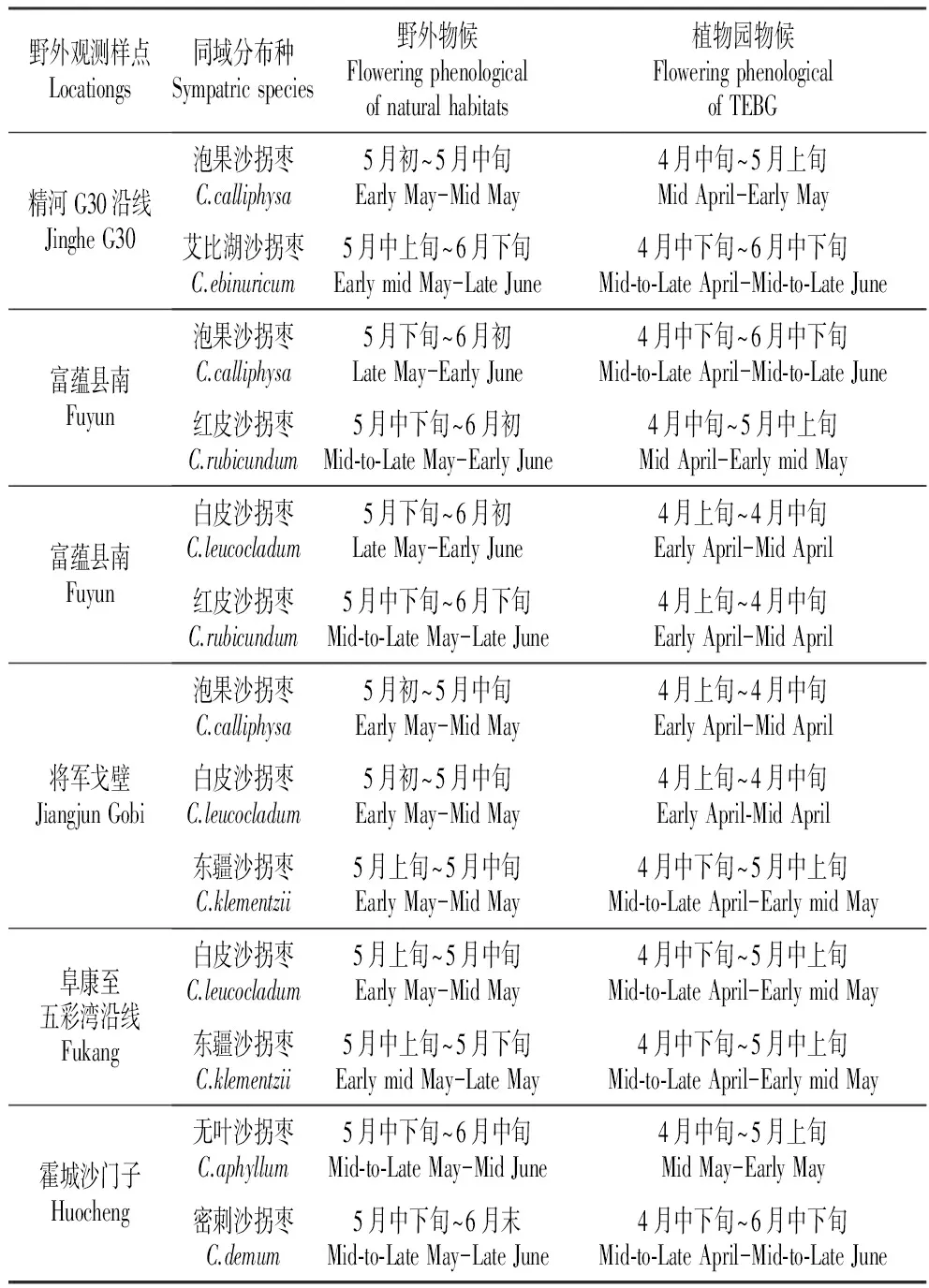
野外观测样点Locationgs同域分布种Sympatricspecies野外物候Floweringphenologicalofnaturalhabitats植物园物候FloweringphenologicalofTEBG精河G30沿线JingheG30泡果沙拐枣C.calliphysa5月初~5月中旬EarlyMay-MidMay4月中旬~5月上旬MidApril-EarlyMay艾比湖沙拐枣C.ebinuricum5月中上旬~6月下旬EarlymidMay-LateJune4月中下旬~6月中下旬Mid⁃to⁃LateApril-Mid⁃to⁃LateJune富蕴县南Fuyun泡果沙拐枣C.calliphysa5月下旬~6月初LateMay-EarlyJune4月中下旬~6月中下旬Mid⁃to⁃LateApril-Mid⁃to⁃LateJune红皮沙拐枣C.rubicundum5月中下旬~6月初Mid⁃to⁃LateMay-EarlyJune4月中旬~5月中上旬MidApril-EarlymidMay富蕴县南Fuyun白皮沙拐枣C.leucocladum5月下旬~6月初LateMay-EarlyJune4月上旬~4月中旬EarlyApril-MidApril红皮沙拐枣C.rubicundum5月中下旬~6月下旬Mid⁃to⁃LateMay-LateJune4月上旬~4月中旬EarlyApril-MidApril将军戈壁JiangjunGobi泡果沙拐枣C.calliphysa5月初~5月中旬EarlyMay-MidMay4月上旬~4月中旬EarlyApril-MidApril白皮沙拐枣C.leucocladum5月初~5月中旬EarlyMay-MidMay4月上旬~4月中旬EarlyApril⁃MidApril东疆沙拐枣C.klementzii5月上旬~5月中旬EarlyMay-MidMay4月中下旬~5月中上旬Mid⁃to⁃LateApril-EarlymidMay阜康至五彩湾沿线Fukang白皮沙拐枣C.leucocladum5月上旬~5月中旬EarlyMay-MidMay4月中下旬~5月中上旬Mid⁃to⁃LateApril-EarlymidMay东疆沙拐枣C.klementzii5月中上旬~5月下旬EarlymidMay-LateMay4月中下旬~5月中上旬Mid⁃to⁃LateApril-EarlymidMay霍城沙门子Huocheng无叶沙拐枣C.aphyllum5月中下旬~6月中旬Mid⁃to⁃LateMay-MidJune4月中旬~5月上旬MidMay-EarlyMay密刺沙拐枣C.demum5月中下旬~6月末Mid⁃to⁃LateMay-LateJune4月中下旬~6月中下旬Mid⁃to⁃LateApril-Mid⁃to⁃LateJune
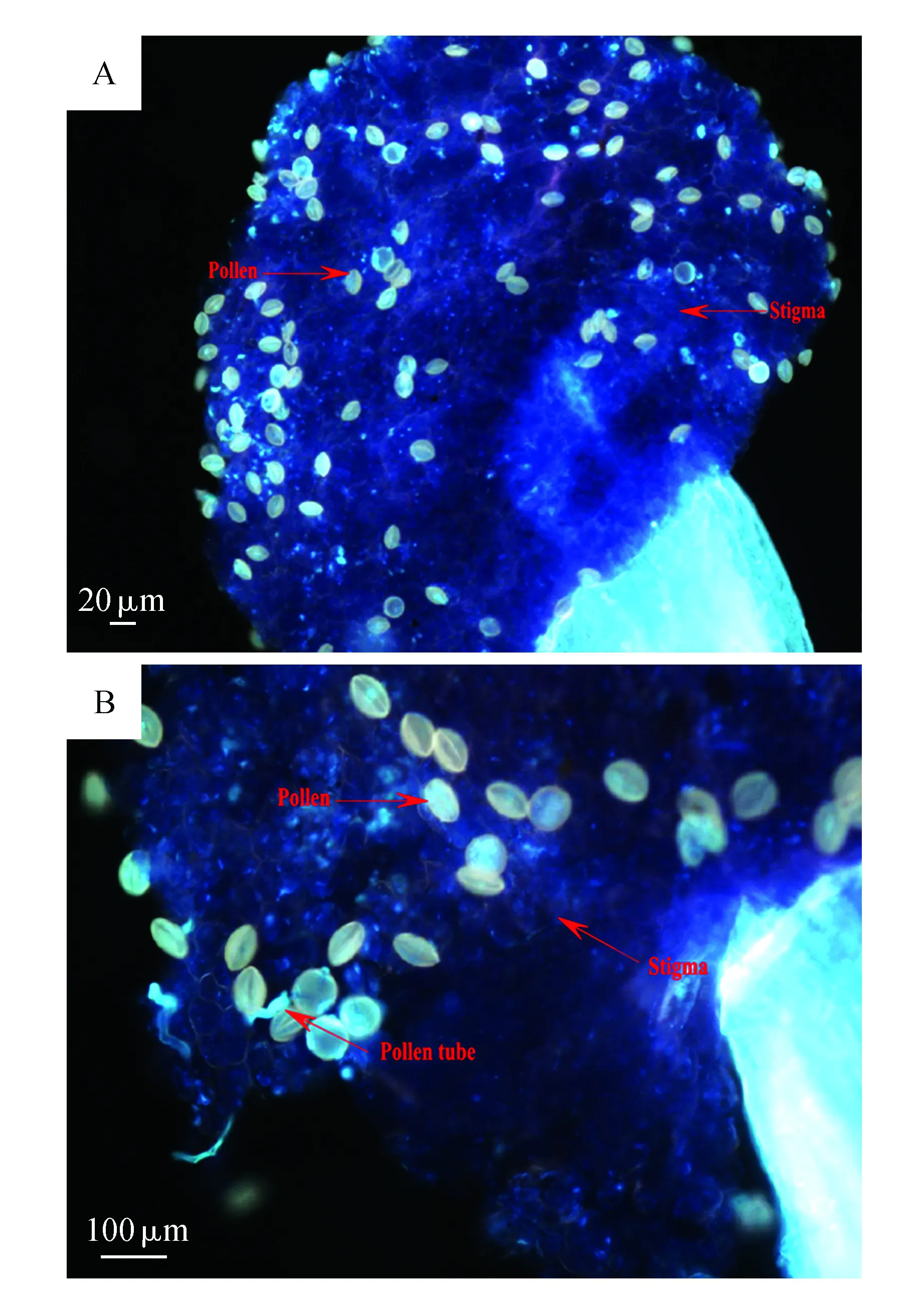
图1 杂交花粉在柱头上的生长情况 A.柱头表面的花粉粒;B:花粉在柱头上萌发的花粉管(荧光显微镜观测)Fig.1 The growth of hybrid pollen on stigmaA.The pollen on the surface of stigma; B.The growth of the pollen tube on the stigma(as viewed by using a fluorescence microscope)
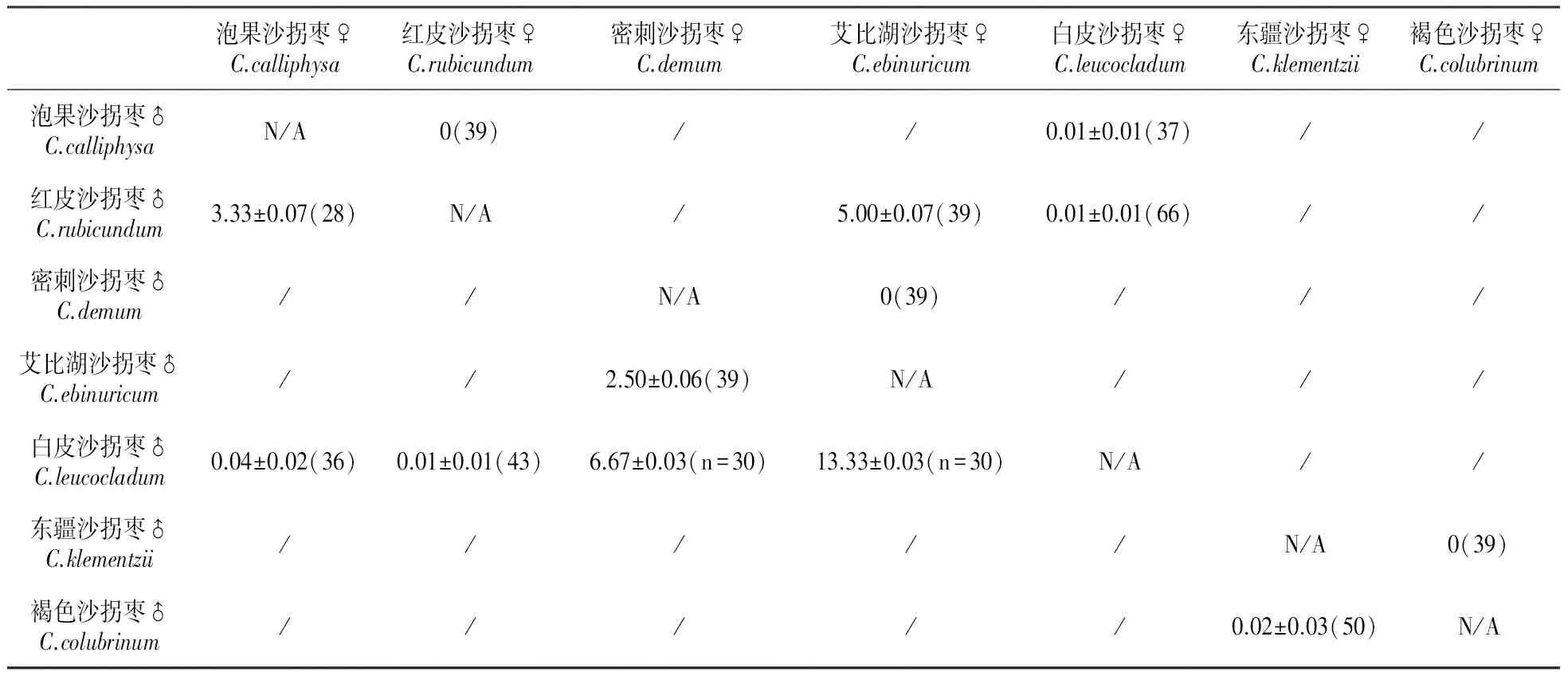
泡果沙拐枣♀C.calliphysa红皮沙拐枣♀C.rubicundum密刺沙拐枣♀C.demum艾比湖沙拐枣♀C.ebinuricum白皮沙拐枣♀C.leucocladum东疆沙拐枣♀C.klementzii褐色沙拐枣♀C.colubrinum泡果沙拐枣♂C.calliphysaN/A0(39)//0.01±0.01(37)//红皮沙拐枣♂C.rubicundum3.33±0.07(28)N/A/5.00±0.07(39)0.01±0.01(66)//密刺沙拐枣♂C.demum//N/A0(39)///艾比湖沙拐枣♂C.ebinuricum//2.50±0.06(39)N/A///白皮沙拐枣♂C.leucocladum0.04±0.02(36)0.01±0.01(43)6.67±0.03(n=30)13.33±0.03(n=30)N/A//东疆沙拐枣♂C.klementzii/////N/A0(39)褐色沙拐枣♂C.colubrinum/////0.02±0.03(50)N/A
4 讨论
物种的形成与维持需要种间隔离机制作为保证,是物种得以保持完整性和独立性的基础[16]。隔离机制根据在植物生活史中起作用的不同阶段一般划分为合子前隔离和合子后隔离。合子前隔离包括生态地理隔离、开花时间隔离、传粉者隔离,另外还包括花粉竞争、配子不亲和;合子后隔离包括杂种不活,不育和杂种衰退[17~20]。
开花物候结果显示,同域分布的沙拐枣属植物的开花物候或多或少的存在重叠,虽然有些只是末花期和始花期相遇,但花期仍不能完全阻断种间的花粉交流。
沙拐枣属植物的花部特征相似,仅在颜色和大小上略有差异,属于泛化传粉植物,依赖昆虫授粉以实现繁殖成功[21],因此,传粉者的访花行为可能也不足以隔离同域分布花期重叠的沙拐枣属植物间的基因交流。
通过荧光显微实验,我们发现种间花粉在柱头上能萌发,并伸长,但不能生长到子房与胚珠受精,表明种间存在合子前隔离机制。人工控制授粉实验也能够较好的说明这7个种杂交的不亲和性,虽然有个别结实,但不具有可育性,表明种间存在合子后隔离机制。相对于外部隔离机制,内部隔离机制更可靠一些,外部隔离机制易受到环境条件的影响,当环境剧烈变化时,甚至会彻底瓦解[22]。在自然界,合子前生殖隔离往往和合子后隔离一起作用[23~28]。在Macior[29~32]的研究中,他就认为在马先蒿属(Pedicularis)外部和内部隔离机制共同作用维持了该属的隔离屏障。在本研究中,沙拐枣属种间的隔离也是外部和内部隔离机制共同起作用的。
通过本研究,我们认为这些同域分布的沙拐枣属植物之间不存在杂交,并且合子前和合子后均存在生殖隔离,隔离机制还需要进一步研究。
不存在杂交这一结论的提出,否定了“存在天然杂交”的假设,为该属的分类争议提供了新的想法,也为果实多型提供了新的见解。
沙拐枣属种间不存在杂交,也是对吐鲁番沙漠植物园中沙拐枣属植物迁地保护近四十年多年工作的肯定,为今后建立世界沙拐枣种质基因圃提供了理论支持。
同域重叠分布的沙拐枣属植物不存在自然种间杂交或基因渐渗,那么同域分布物种各自居群遗传多样性水平和遗传结构是怎样的呢?目前尚不得而知。进一步地,我们将利用微卫星等分子标记对该属的遗传多样性和遗传结构进行研究。
致谢沈观冕研究员推荐、翻译俄文文献,特致谢忱!
1.Bao B J,Grabovskaya-borodina A E.Flora of China,Vol 5[M].Beijing:Beijing Science Press,2003:325-329.
2.毛祖美.新疆植物志:第1卷[M].乌鲁木齐:新疆科技卫生出版社,1992:265-275.
Mao Z M.CalligonumL.[M].//Yang C Y.Flora Xinjiangensis,vol.1.Urumqi:Xinjiang Science & Technology & Hygiene Publishing House,1992:118-133.
3.康晓珊,张永智,潘伯荣,等.不同地区精河沙拐枣居群果实形态差异性分析[J].西北植物学报,2008,28(6):1213-1221.
Kang X S,Zhang Y Z,Pan B R,et al.The fruits morphological characteristics of different population inCalligonumebi-nuricum[J].Acta Bot Borea1-Occident Sin,2008,28(6):1213-1221.
4.康晓珊,潘伯荣,张永智,等.中国特有植物艾比湖沙拐枣(Calligonumebi-nuricum)居群内果实性状的变异[J].植物研究,2009,29(6):747-752.
Kang X S,Pan B R,Zhang Y Z,et al.Variation in Fruit Characteristics of the Chinese Endemic SpeciesCalligonumebi-nuricumin Population[J].Bulletin of Botanical Research,2009,29(6):747-752.
5.李得禄,王继和,马全林.沙拐枣属植物果实性状的数值分类研究[J].中国沙漠,2009,29(4):718-722.Li D L,Wang J H,Ma QL.Numerical Taxonomy of Fruit Characteristic ofCalligonumL.[J].Journal of desert research,2009,29(4):718-722.
6.师玮,潘伯荣,段士民,等.蒙古沙拐枣(Calligonummongolicum)与其相关种的果实形态差异性分析[J].中国沙漠,2011,31(1):121-128.
Shi W,Pan B R,Duan S M,et al.Difference of Fruit Morphological Characters ofCalligonummongolicumand Related Species[J].Journal of desert research,2011,31(1):121-128.
7.МУСАЕВ ИФ,СОСКВ ЮД.О географии и Филогении представителей родаCalligonumL.[J].Бот.ж,1977,62(10):1415-1432.
8.СОСКВ ЮД.Использование закона гомологический рядов Н.И.Вавилова в систематиге на примере изучения родаCalligonumL.[J].Бот.ж,1968,53(4):470-479.
9.СОСКВ ЮД.СекцияCalligonumРодаCalligonumL.Новасти сист[J].Bысших раст,1975,12:147-159.
10.СОСКВ ЮД.Новые серии подвиды и гибриды в родеCalligonumL.[J].Бот.ж,1975,60(8):1162-1163.
11.毛祖美,潘伯荣.我国沙拐枣属的分类与分布[J].中国科学院大学学报,1986,24(2):98-107.
Mao Z M,Pan B R.Thee classication and distribution of the genusCalligonumL.in China[J].Acta Phytotaxonomica Sinica,1986,24:98-107.
12.ЧЕРЕПАНΟВ С К.Сосудистые растения Ссср.[M].Лeнинград:Ленинардское отделение Наука,1981:397-398.
13.尹林克.中国科学院吐鲁番沙漠植物园[J].干旱区研究,2004,24(21):1-4.
Yin L K.Turpan Eremophytes Botanic Garden of the Chinese Academy of Sciences-a developing conservation and research base of plant resources diversity in desert area[J].Arid Zone Research,2004,21:1-4.
14.康晓珊,潘伯荣,段士民,等.沙拐枣属4种植物同地栽培开花物候与生殖特性比较[J].中国沙漠,2012,32(5):1315-1327.
Kang X S,Pan B R,Duan S M,et al.Comparison of flowering phenology and reproductive traits of four species ofCalligonunL.(Polygonaceae) under ex-situ conservation at the Turpan Eremophytes Botanical Garden in Xinjaing,China[J].Journal of desert research,2012,32(5):1315-1327.
15.赵艳芬,师玮,潘伯荣,等.沙拐枣属(CalligonumL.)植物物候对长期气温变化的响应[J].中国沙漠,2014,34(3):732-739.
Zhao Y G,Shi W,PAN B R,et al.The phenophase response ofCalligonumL.in Turpan Eremophytes Botanical Garden to temperature change in recent 29 years[J].Journal of desert research,2014,34(3):732-739.
16.Crawford D J.Electrophoretic data and plant speciation[J].Systematic Botany,1985,10(4):405-416.
17.Grant V.Plant speciation[M].2nd ed.New York:Columbia University Press,1981:71-76.
18.Schemske D W.Understanding the origin of species[J].Evolution,2000,54(3):1069-1073.
19.Rieseberg L H,Willis J H.Plant speciation[J].Science,2007,317(5840):910-914.
20.张敬丽,张长芹,关之坤,等.探讨种间传粉在杜鹃花属自然杂交物种形成中的作用[J].生物多样性,2007,15(6):658-665.
Zhang J L,Zhang C Q,Guan ZK,et al.The potential roles of interspecific pollination in natural hybridization of Rhododendron species in Yunnan,China[J].Biodiversity Science,2007,15(6):658-665.
21.康晓珊,索菲亚,段士民,等.迁地保护条件下4种沙拐枣(Calligonum)的花部特征和传粉特性[J].中国沙漠,2015,35(5):1239-1247.
Kang X S,Sophia,Duan S M.Floral syndrome and pollination characteristics of fourCalligonumspecies(Polygonaceae) under ex-situ conservation[J].Journal of desert research,2015,35(5):1239-1247.
22.Grant V.Floral isolation between ornithophilous and sphingophilous species ofIpomopsisandAquilegia[J].Proceedings of the National Academy of Sciences of the United States of America,1992,89(24):11828-11831.
23.Sprague E F.Pollination and evolution inPedicularis(Scrophulariaceae)[J].Aliso,1962,5:181-209.
24.Grant V.Mechanical and ethological isolation betweenPedicularisgroenlandicaandP.attllens(Scrophulariaceae)[J].Biologisches Zentralblatt,1994,113:43-51.
25.Grant V.Modes and origins of mechanical and ethological isolation in angiosperms[J].Proceedings of the National Academy of Sciences of the United States of America,1994,91(1):3-10.
26.Gardner M,Macnair M.Factors affecting the co-existence of the serpentine endemicMimulusnudatusCurran and its presumed progenitor,MimulusguttatusFischer ex DC[J].Biological Journal of the Linnean Society,2000,69(4):443-459.
27.Ando T,Nomura M,Tsukahara J,et al.Reproductive isolation in a native population ofPetuniasensuJussieu(Solanaceae)[J].Annals of Botany,2001,88(3):403-413.
28.Yang C F,Gituru R W,Guo Y H.Reproductive isolation of two sympatric louseworts,PedicularisrhinanthoidesandPedicularislongiflora(Orobanchaceae):how does the same pollinator type avoid interspecific pollen transfer?[J].Biological Journal of the Linnean Society,2007,90(1):37-48.
29.Macior L W.The pollination ecology of Pedicularis on Mount Rainier[J].American Journal of Botany,1973,60(9):863-871.
30.Macior L W.The pollination ecology ofPedicularis(Scrophulariaceae) in the Sierra Nevada of California[J].Bulletin of the Torrey Botanical Club,1977,104(2):148-154.
31.Macior L W.Plant community and pollinator dynamics in the evolution of pollination mechanisms inPedicularis(Scrophulariaceae)[M].//Armstrong J A,Powell J M,Richards A J.Pollination and evolution.Sydney:Royal Botanic Gardens,1982:29-45.
32.Macior L W.The pollination dynamics of sympatric species ofPedicularis(Scrophulariaceae)[J].American Journal of Botany,1983,70(6):844-853.
This work was supported by National Natural Science Foundation of China NSFC(No.31200237 and 41271069)
introduction:KANG Xiao-Shan(1982—),female,Assistant researcher,plantreproductive biology.
date:2016-04-26
PreliminaryStudiesonHybridizationamongSympatricDistributedCalligonumSpeciesinNorthernXinjiang
KANG Xiao-Shan1,2Sophia3DUAN Shi-Min1,2PAN Bo-Rong1,2ZHANG Yong-Zhi2TIAN Cong4
(1.Xinjiang Institute of Ecology and Geography,Chinese Academy of Sciences,Key Laboratory of Biogeography and Bioresource in Arid Land,Urumqi 830011;2.Turpan Eremophytes Botanical Garden,Chinese Academy of Sciences,Turpan 838008;3.College of Life Science and Technology,Xinjiang University,Urumqi 830046;4.Grassland Institute,Xinjiang Academy of Animal Science,Urumqi 830000)
Hybridization is one key issue in taxonomy of the genusCalligonum. To certificate whether or not hybridization are existing among sympatric distributedCalligonumspecies, observations of flowering phenology both in natural habitats and botanical garden, artificial pollination hybridization experiments and fluorescence microscope observation on cross-pollination of pollen tubes were carried on. Florescence among the sympatric species was overlapped. Fruit rate by artificial pollination was very low, and could not germinate. Pollen tube fluorescent experiments showed that hybrid pollen could germinate on the stigma, but the pollen tube could not stretch to the ovary. Above all, affinity of interspecific gametes was low, reproductive isolation was existing, and hybridization was absent. This study has important implications for understanding the isolation and differentiation of sympatricCalligonumspecies.
CalligonumL.;flowering phenology;hybridization;fluorescence microscopy
国家自然科学青年基金资助项目(31200237);国家自然科学基金面上项目(41271069)
康晓珊(1982—),女,博士,助理研究员,主要从事繁殖生物学研究。
2016-04-26
S793.1
A
10.7525/j.issn.1673-5102.2016.05.022
