13C辅助的超高效液相色谱-三重四极杆质谱联用方法精确分析毕赤酵母胞内代谢物浓度
2016-11-01郭孟磊刘晓云黄明志李敏超庄英萍张嗣良
郭孟磊 刘晓云 黄明志李敏超 储 炬 庄英萍 张嗣良
(华东理工大学国家生化工程技术研究中心(上海),上海 200237)
13C辅助的超高效液相色谱-三重四极杆质谱联用方法精确分析毕赤酵母胞内代谢物浓度
郭孟磊 刘晓云 黄明志*李敏超 储 炬 庄英萍 张嗣良
(华东理工大学国家生化工程技术研究中心(上海),上海 200237)
毕赤酵母作为一种高效的外源蛋白表达平台,其蛋白表达水平与胞内代谢物浓度紧密相关。但胞内代谢物种类多、物化性质差异大、浓度低、周转快,对其绝对浓度的精确检测一直难于实现。本研究将超高效液相色谱-三重四极杆质谱联用分析方法与13C同位素标记技术相结合,探索解决该难题的方法。首先,优化了超高效液相色谱的操作条件,利用3种色谱柱实现了64种常见中间代谢物的分离;对三重四极杆质谱仪的检测离子对和碰撞电压等操作条件进行优化,找到了对各种物质具有专一性的检测离子对。然后,利用全标记13C标记底物培养细胞,收集胞内的全标记代谢物用作定量内标物,建立了53种中间代谢物的标准曲线。实验结果表明,本方法不但精确性高,标准曲线相关系数达到0.99以上,而且重现性好,受实验条件和仪器操作条件的影响很小。将本方法应用于毕赤酵母胞内代谢物浓度的绝对定量分析,成功获得了胞内各种代谢物的浓度水平,为后续深入研究毕赤酵母代谢调控机理,实现外源蛋白的高效生产奠定了基础。
超高效液相色谱-三重四极杆质谱;13C同位素标记;代谢物浓度;毕赤酵母
1 引言
毕赤酵母是一种高效的蛋白表达平台,已用于1000多种外源蛋白的表达,其表达水平与胞内代谢物浓度紧密相关。掌握毕赤酵母胞内中间代谢物的浓度信息对理解其代谢调控机理,实现外源蛋白的高效生产具有重要意义[1~4]。
中间代谢物具有种类多、物化性质差异大、浓度低和周转快等特点,色谱-质谱联用技术已大量应用于这些物质的定性与定量分析。常用的质谱定量方法包括气相色谱-质谱法(GC-MS)和液相色谱-质谱联用法(LC-MS)。气相色谱不适于分离强极性代谢物,液相色谱-质谱更适合于中间代谢物的定量分析[5,6],但其定量性能受到离子抑制效应[7]和样品前处理过程的影响。为弥补这些不足,Wu和Bennett[7,8]等提出同位素稀释-质谱定量法(简称IDMS),通过测量样品的未标记代谢物和作为内标的全标记代谢物的比值实现定量分析[9]。
IDMS已应用于大肠杆菌和面包酵母等多个细胞体系[6,10]。已有研究者尝试将其引入到毕赤酵母体系中,如Carnicer等[11]对比了5种淬灭方法的影响,发现温度-27℃、甲醇含量60%时效果最好;Tredwell等[12]分析了不同提取方法的影响,指出沸乙醇提取方法具有优越性;Jordà等[13]将其与非稳态13C代谢通量分析技术相结合,探讨了外源蛋白表达对毕赤酵母胞内代谢的影响。尽管IDMS理论上是一种准确度和精度较高的方法[7],仍有多种因素可能会影响其定量性能。毕赤酵母的代谢活性很强,在定量分析其胞内代谢物浓度时,与传统定量方法相比,有关IDMS方法的优势和不足尚缺乏深入研究。本研究优化了LC-MS/MS检测毕赤酵母胞内代谢物的操作条件,建立各种代谢物的标准曲线,评估IDMS方法在定量分析不同代谢物时具有的优势和不足,探讨实现毕赤酵母胞内代谢物浓度可靠检测的方法。
2 实验部分
2.1 仪器与试剂
超高效液相色谱-三重四级杆串联质谱(Thermo Fisher公司),ACQUITY UPLC®RBEH Amide色谱柱(100 mm×2.1 mm,1.7μm,Waters公司);XselectTMHSS T3色谱柱(150 mm ×2.1 mm,3.5μm,Waters公司);XselectTMHSS T3色谱柱(100 mm ×2.1 mm,1.8μm);振荡器(Thermo Fisher公司);移液器(GILSON公司);SHZ-D(Ⅲ)循环水式真空泵(上海予华仪器设备有限公司);纤维滤膜(Millipore公司);pH计(Mettler Toledo公司)。
标准品和甲酸、乙酸、二甲基己二胺、三丁胺、甲酸铵等试剂(Sigma公司);流动相甲醇、异丙醇、乙腈、超纯水(Fisher Chemical公司)。U13C葡萄糖(Cambridge Isotope Laboratories公司)。
2.2 菌种和培养条件
毕赤酵母β-半乳糖苷酶生产菌株GHL。培养基组成为葡萄糖20 g/L,MgSO4·7H2O 14.9 g/L,K2SO418.2 g/L,85%H3PO426.8 mL/L,无水Ca2SO40.93 g/L,KOH 4.13 g/L,消泡剂(1∶1000,V/V)。微量元素溶液(12∶1000,V/V)其中微量元素:KI 0.08 g/L,CuSO4·5H2O 6.0 g/L,MnSO4·H2O 3.0 g/L,FeSO4·7H2O 65.0 g/L,H2SO4(1∶200,V/V),NaMoO4·2H2O 0.2 g/L,CoCl20.5 g/L,ZnCl220.0 g/L,H3BO40.02 g/L,生物素0.2 g/L。培养条件:接种量10%,温度30℃,pH 5.5,每升发酵液的通气量1 L/min。
13C全标记菌体的培养采用U13C葡萄糖为唯一碳源,接种量1%,空气中CO2用NaOH溶液除去,其它条件与上述相同。提取菌体胞内代谢物检测U13C代谢物所占比例,发现其中仅含有极微量的12C代谢物,不影响化合物浓度的定量分析。
2.313C全标菌体样品的处理
发酵结束后,将100 mL发酵液加入-20℃预冷的含有500 mL 60%甲醇的补料瓶中,搅拌混匀后分装到30个试管中,每个试管20 mL,4℃,12000 r/min离心10 min,弃上清液。在试管加入70℃预热的20 mL 75%乙醇,95℃保温10 min。冷却至室温后,4℃、以12000 r/min离心10 min,取上清液,将30支试管中上清液混合后再分装。
2.4 标记实验样品的处理
样品取出后直接放入27℃预冷的含有3 mL 60%甲醇的试管灭活[10],取样量为1 mL。用0.8μm纤维滤膜过滤,去离子水清洗菌体;滤膜与菌体均加入70℃预热的含有30 mL 75%乙醇的试管中,95℃保温10 min,用于定量分析的样品同时还加入100 μL内标。提取代谢物后将滤膜取出,4℃,以12000 r/min离心10 min,取上清液。上清液用快速旋转蒸发仪蒸干后-80℃保存待测。
2.5 色谱条件
2.5.1 有机酸与磷酸糖类 采用 Acquity T3色谱柱(150 mm×2.1 mm,1.8μm)。流动相 A: 5 mmol/L三丁胺,15 mmol/L乙酸,5% 甲醇;流动相B:异丙醇。柱温:40℃。梯度洗脱:0~5 min,100%A;5~16 min,100%~91%A;16~19 min,91%~50%A;19~25 min,50%A;25~26 min,50%~100%A。流速0.2 mL/min。
2.5.2 核苷酸类 采用XselectTMHSS T3色谱柱(100 mm×2.1 mm,1.8μm)。流动相A:5 mmol/L三丁胺,以甲酸调至pH 4.95;流动相B:5 mmol/L三丁胺,100%ACN,0.5%甲酸。柱温:40℃。梯度洗脱:0~30 min,100%~50%A。流速0.2 mL/min。
2.5.3 氨基酸类 采用ACQUITY UPLCR BEH Amide色谱柱(100 mm×2.1 mm,1.7μm)。流动相A:5 mmol/L三丁胺,以甲酸调至pH 3.0;流动相B:5 mmol/L三丁胺,85%ACN,0.5%甲酸。柱温:40℃。梯度洗脱:0~30 min,100%~75%B。流速0.2 mL/min。
2.6 质谱条件
氨基酸的检测采用正离子模式:喷雾电压3.5 kV,汽化温度300℃,鞘气压15 psi,离子扫描气压0 psi,辅助气压10 psi,毛细管温度300℃。
有机酸、磷酸糖和核苷类物质的检测采用负离子模式:喷雾电压 -3.0 kV,汽化温度200℃,鞘气压15 psi,离子扫描气压0 psi,辅助气压10 psi,毛细管温度200℃。
2.7 标准曲线的建立
分别采用传统方法和IDMS方法进行胞内代谢物浓度的绝对定量分析。传统方法只利用标准品浓度与峰面积之间的对应关系建立标准曲线。采用IDMS方法时,将不同浓度梯度的化合物标准品(未标记)与U13C内标按4∶1比例混合,根据未标记代谢物与全标记代谢物的峰面积比值与标准品浓度建立标准曲线。
2.8 加标回收实验
回收率(R)定义为:

其中,ΔCm是添加已知浓度12C标准品后与添加前测定值之差,ΔC是理论上添加前后的差值,

其中C0为未添加前代谢物浓度;V0为未添加前样品体积;Ca和Va分别是添加标准品的浓度和体积。
3 结果与讨论
3.1 色谱操作条件优化
胞内有机酸、磷酸糖和核苷类物质由于极性大,pKa值低,存在同分异构体,使得对该类化合物的分离成为难题,而氨基酸类物质极性差异大,pKa值范围广,也成为分离氨基酸的最大障碍。根据这3类物质性质的不同,首先利用选择离子扫描模式[14]对色谱分离方法进行了优化,利用3根色谱柱有效分离了包括有机酸、磷酸糖、核苷类物质和氨基酸在内的64种常见胞内代谢物(图1~图3)。质谱通过质荷比来鉴别化合物,但是不能区分同分异构体,因此色谱分离至关重要,本研究有效分离了上述几种化合物,尤其是实现了G6P、F6P、M6P等同分异构体的分离,为更有效鉴别代谢物奠定了基础。
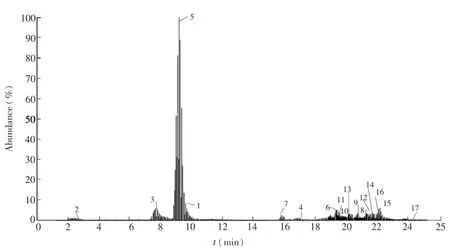
图1 有机酸、磷酸糖选择离子扫描总离子流图Fig.1 Total ion chromatograms of sugar phosphates and carboxylic acids
3.2 质谱仪操作条件优化
用标准品进行质谱操作条件优化,获得了64种代谢物的专一检测离子对及最佳操作条件,如表1所示。化合物在碰撞池中发生电离,虽然不同的化合物电离方式不同,但相似化合物电离方式具有相似性,例如磷酸糖类化合物特征离子多为,有机酸类化合物以羧基为主,表1给出了所有化合物子离子分子式,为后续代谢物的检测和合理选择标记化合物离子对提供了依据。
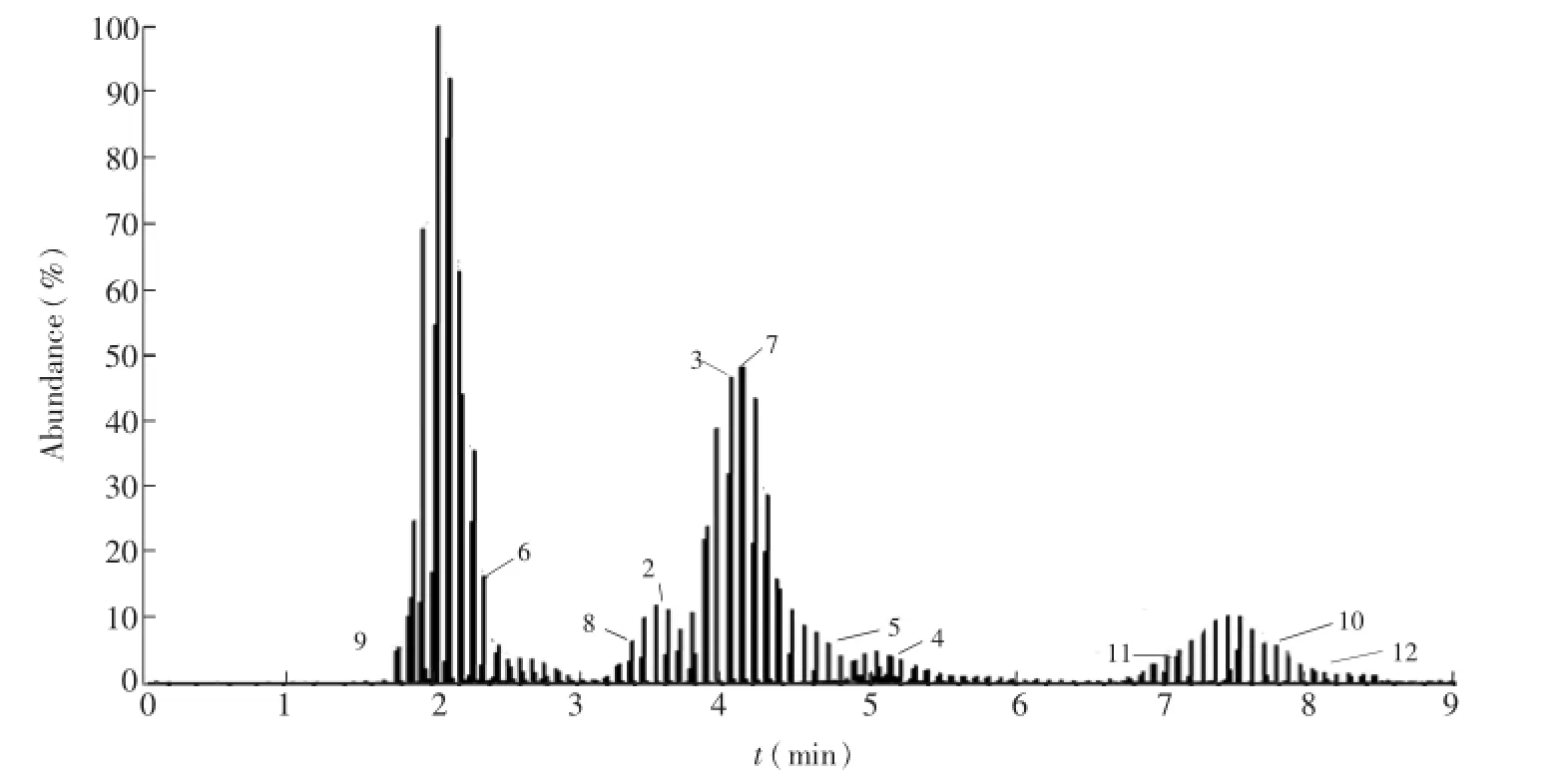
图2 核苷类物质选择离子扫描总离子流图Fig.2 Total ion chromatograms of nucleosides
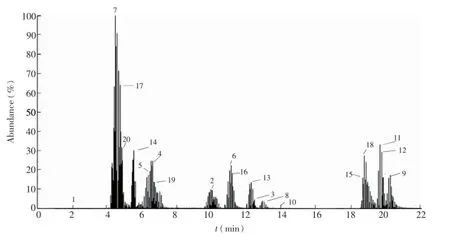
图3 20种氨基酸物质选择离子扫描总离子流图Fig.3 Total ion chromatograms of 20 amino acids
3.3 标准曲线
从表2可见,对于大多数物质而言,基于13C内标的标准曲线的线性系数要好于传统的12C方法。但是部分物质,如CDP,SUC,OXA和部分氨基酸,13C方法所得的线性系数反而小于12C方法。究其原因,可能是这些物质提取时在胞内降解太快,可保留用作13C内标物的量较少,导致内标峰面积的测定误差变大,使得其线性系数反而变小,有些化合物的IDMS标准曲线甚至无法建立。
理论上,标记代谢物与未标记代谢物具有色谱同一性,因此基于13C内标的标准曲线方程的斜率只与该U13C代谢物在标记细胞中的含量和稀释比例有关。在进样量确定时,两者保持不变,标准曲线斜率就不会发生变化,因此该斜率不会随仪器状态和不同的分析样品批次的变化而变化,而12C方法因缺少内标物,峰面积会发生一些变化,从而影响其斜率。为了验证,依照上述方法做出标准曲线,连续测量4次,得到不同进样批次之间标准曲线斜率的变化情况(表3)。从表3可知,13C法标准曲线比12C法具有更好的稳定性。
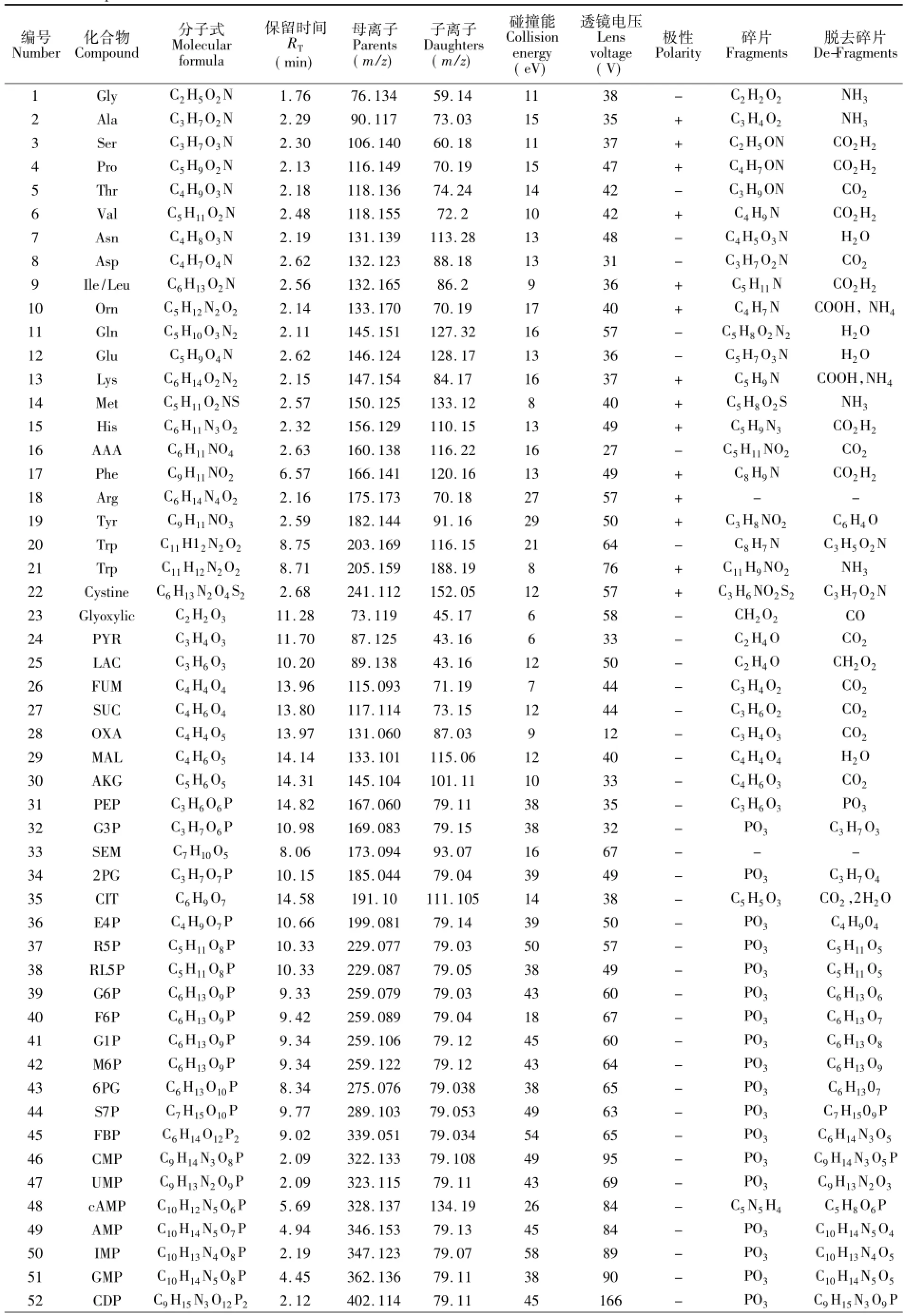
表1 SIM检测离子对Table 1 Ion pair of SIM
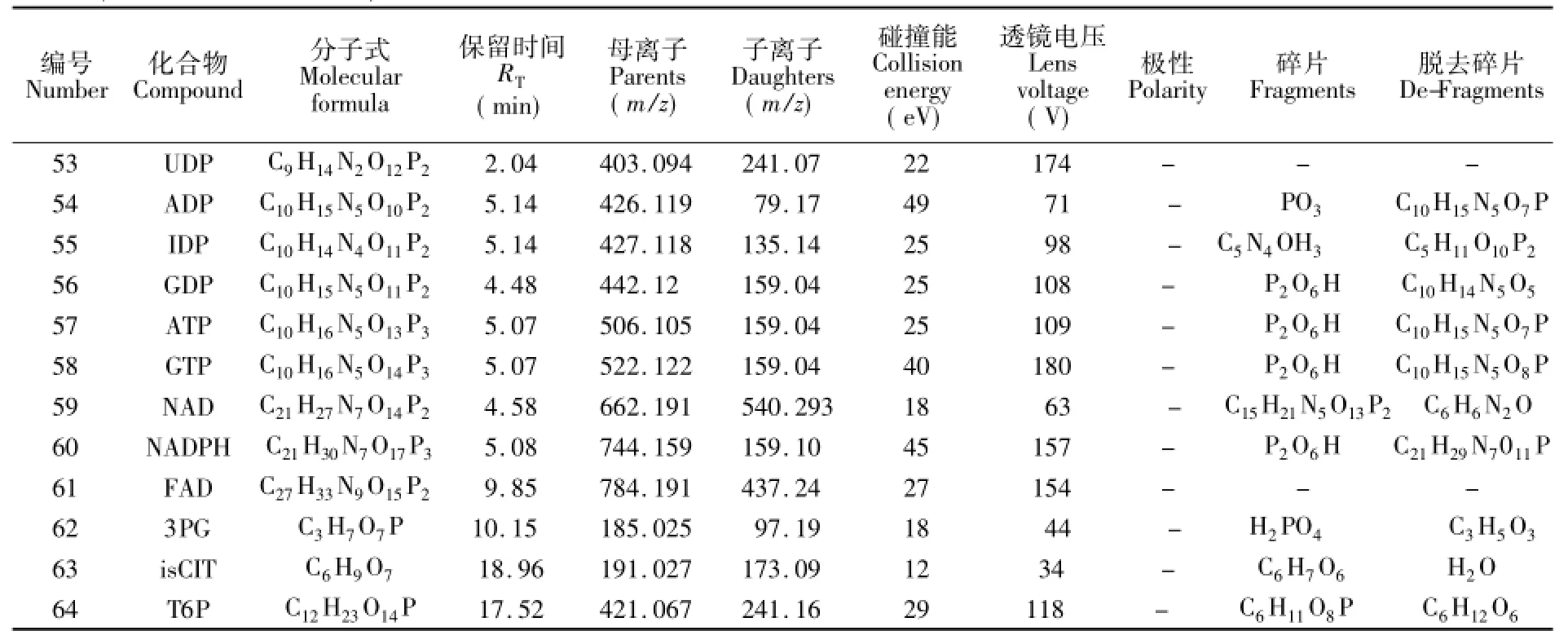
续表1(Continued to Table 1)

表2 标准曲线相关系数(R2值)Table 2 Correlation coefficients(R2values)of calibration lines
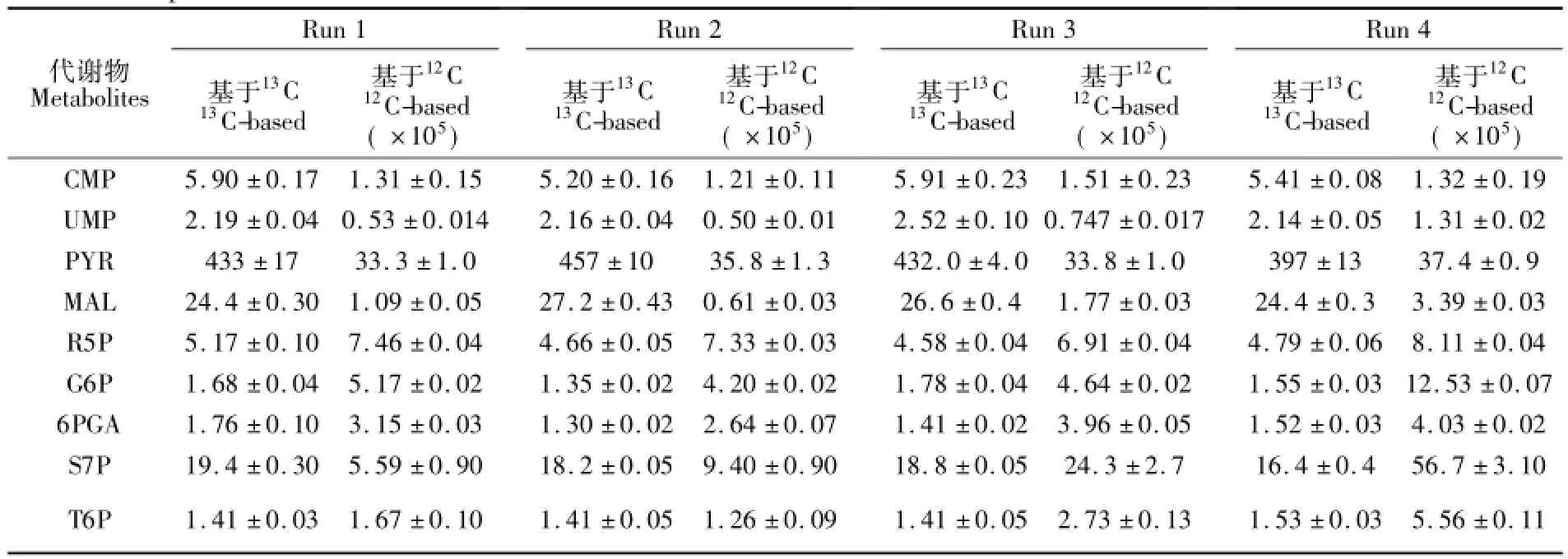
表3 4次重复实验中标准曲线的斜率及其95%的置信区间Table 3 Slopes and their 95%confidences intervals in four LC-ESI-MS/MS runs
3.4 回收率
表4给出了部分化合物的回收率偏差,从回收率偏差上可以看出,IDMS方法回收率偏差小于传统12C方法,证明本方法的优越性和重要性。
3.5 毕赤酵母胞内代谢物定量分析结果
不同菌株的细胞在不同生长状态下其胞内代谢物浓度存在一定的差异,但是对于特定种类生物而言,其胞内代谢物浓度水平和细胞的容量具有相似性。将IDMS方法与传统12C(Lac,Asp等)方法相结合,能更完整地测定毕赤酵母胞内代谢物。图4给出了毕赤酵母胞内代谢物的定量结果,无论是胞内各种代谢物的浓度水平,例如有机酸、磷酸糖(0.01~50μmol/g细胞干重(DCW))、核苷类物质(0.01~10μmol/g DCW)、游离氨基酸(0.1~100μmol/g DCW),还是胞内游离氨基酸的总量(378.02μmol/g DCW),或者不同氨基酸在毕赤酵母胞内的百分含量,都与文献[12,15,16]报道一致,说明了本定量方法的可靠性。
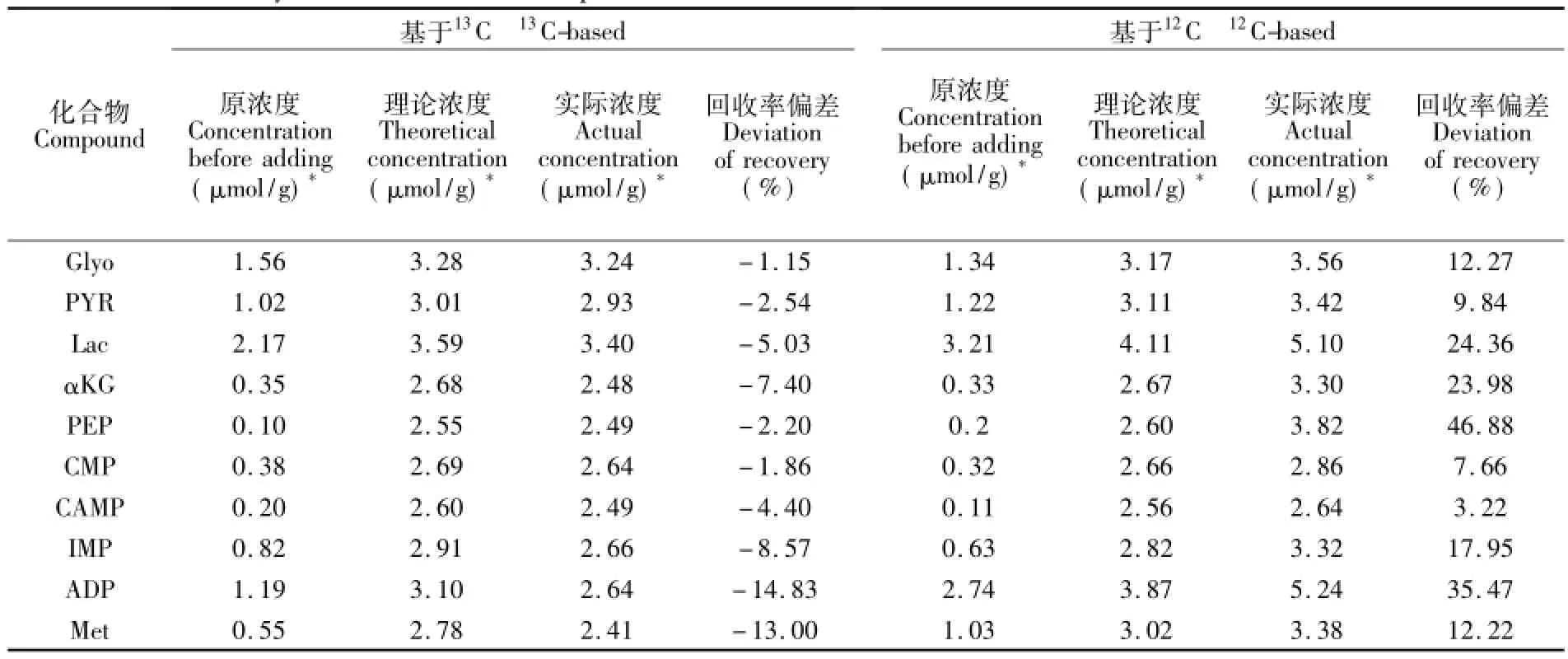
表4 部分化合物的回收率偏差Table 4 The recovery deviation for some compounds
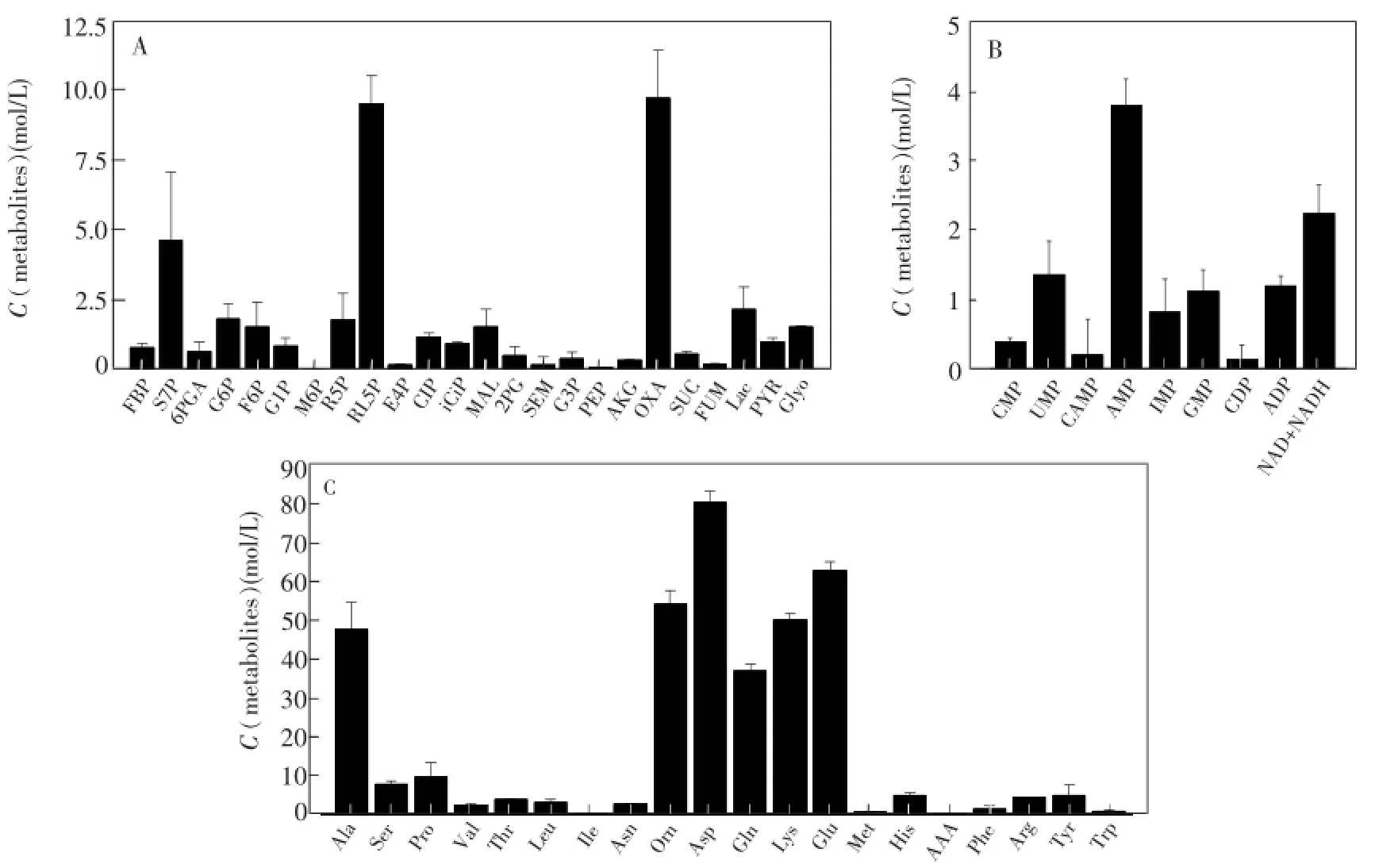
图4 毕赤酵母GHL胞内代谢物浓度(μmol/g细胞干重)Fig.4 Intracellular metabolites concentration in Pichia pastoris GHL.Intracellular concentration of metabolites pools are given inμmol/g DCW
4 结论
比较了IDMS方法与传统定量方法在毕赤酵母胞内代谢物定量分析方面的优劣,说明了IDMS方法的优越性,但是也不能忽略传统方法在测量可快速降解代谢物方面的作用,将两者结合,将使得毕赤酵母胞内代谢物的定量分析更加精确完整,为后续深入研究毕赤酵母代谢调控机理,实现外源蛋白的高效生产奠定了基础。
1 Fan H,Li S P,Xiang J J,Lai C M,Yang F Q,Gaoa J L,Wang Y T.Anal.Chim.Acta,2006,567(2):218-228
2 Ogawa T,Washio J,Takahashi T,Echigo S,Takahashi N.Oral Surg.Oral.Med.Oral.Pathol.Oral.Radiol.,2014,118(2):218-225
3 Xia M L,Huang D,Li S S,Wen J P,Jia X Q,Chen Y L.Biotechnol.Bioeng.,2013,110(10):2717-2730
4 Wasylenko T M,Stephanopoulos G.Biotechnol.Bioeng,2014,112(3):470-483
5 Buchholz A,Takors R,Wandrey C.Anal.Biochem.,2001,295(2):129-137
6 van Dam J C,Eman M,Frank J,Lange H C,van Dedem G W K,Heijnen S J.Anal.Chim.Acta,2002,460(2): 209-218
7 Wu L,Mashego M R,van Dam J C,Proell A M,Vinke J L,Ras C,van Winden W A,van Gulik W M,Heijnen J J. Anal.Biochem.,2005,336(2):164-171
8 Bennett B D,Yuan J,Kimball E H,Rabinowitz J D.Nature Protocols,2008,3(8):1299-1311
9 Hellerstein M K,Neese R A.Am.J.Physiol,1999,276(6):1146-1170
10 McCloskey D,Gangoiti G A,King Z A,Naviaux R K,Barshop B A,Palsson B O,Feist A M.Biotechnol.Bioeng.,2014,111(4):803-815
11 Carnicer M,Canelas A.B,Pierick A,Zeng Z,Dam J,Albiol J,Ferrer P,Heijnen J J,Gulik W.Metabolomics,2012,8(2):284-298
12 Tredwell G D,Edwards-Jones B,Leak D J,Bundy J G.PLoS One,2011,6(1):e16286
13 Jordà J,Rojas H C,Carnicer M,Wahl A,Ferrer P,Albiol J.Metabolites,2014,4(2):281-299
14 LI Min-Chao,HUANG Ming-Zhi,LIU Yu-Wei,CHU Ju,ZHUANG Ying-Ping,ZHANG Si-Liang.Chinese J.Anal. Chem.,2014,42(10):1408-1403
李敏超,黄明志,刘玉伟,储炬,庄英萍,张嗣良.分析化学,2014,42(10):1408-1403
15 Jordà J,Suarez C,Carnicer M,ten Pierick A,Heijnen J J,Gulik W,Ferrer P,Albiol J,Wahl A.BMC Syst.Biol.,2013,7(1):1-16
16 Jordà J,Jouhten P,Cámara E,Maaheimo H,Albiol J,Ferrer P.Microb.Cell Fact.,2012,11:57
13C-assisted Ultra-high Performance Liquid Chromatography Triple Quadrupole Mass Spectrometry Method for Precise Determination of Intracellular Metabolites in Pichia pastoris
GUO Meng-Lei,LIU Xiao-Yun,HUANG Ming-Zhi* ,LI Min-Chao,CHU Ju,ZHUANG Ying-Ping,ZHANG Si-Liang
( National Engineering Research Center for Biotechnology ( Shanghai) ,East China University of Science and Technology,Shanghai 200237,China)
The yeast Pichia pastoris is an effective host for recombinant protein production and the recombinant protein production level is tightly related to the concentrations of intracellular metabolites.The intracellular metabolites have the features of wide range of types,distinct variation in physical and chemical properties,rapid turning-over and low concentration,so it is difficult to quantify their concentrations precisely.In this experiment,we tried to make it possible by combining ultra-high performance liquidchromatography(UPLC)-triple quadrupole mass spectrometry and13C isotope labeling techniques.64 metabolites including organic acids,sugar phosphate,nucleoside substance,amino acid were successfully separated by UPLC with three kinds of chromatography column.The appropriate and unique ion pairs and collision voltages were found after the mass spectrometry condition optimization.By using the U13C metabolites as internal standards collected from the cells growing on U13C-glucose as sole carbon source,the standard curves of 53 metabolites were established.The results showed that the method presented here had a high accuracy.The correlation coefficients were above 0.99.The method also had a good reproducibility,and the influence of experimental and equipment operating condition was very small.Reliable and accurate determination of the concentrations of intracellular metabolites in Pichia pastoris was obtained.The works lays the foundation for comprehensive regulation mechanism research and efficient recombinant protein production in Pichia pastoris.
Ultra-high performance liquid chromatography-triple quadrupole mass spectrometry;Carbon-13 isotope labeling;Absolute quantification of intermediate metabolites concentration;Pichia pastoris
10 August 2015;accepted 29 October 2015)
10.11895/j.issn.0253-3820.150635
2015-08-10收稿;2015-10-29接受
*E-mail:249533969@qq.com
