封育年限对高寒草甸群落组分和物种多样性的影响
2016-10-25刘晓琴张立锋李英年徐世晓李红琴马荣荣高玉葆
刘晓琴,张 翔,张立锋,李英年,赵 亮,徐世晓,李红琴,马荣荣,牛 犇,高玉葆,古 松,*
1 南开大学生命科学学院, 天津 300071 2 中国科学院西北高原生物研究所, 西宁 810008
封育年限对高寒草甸群落组分和物种多样性的影响
刘晓琴1,张翔1,张立锋1,李英年2,赵亮2,徐世晓2,李红琴2,马荣荣1,牛犇1,高玉葆1,古松1,*
1 南开大学生命科学学院, 天津300071 2 中国科学院西北高原生物研究所, 西宁810008
高寒草甸在青藏高原畜牧业发展中占有重要地位,然而气候和人为活动的干扰对原本脆弱的生态系统产生了深刻影响。虽然封育是被广泛采用的草地恢复有效措施之一,但封育年限对群落组分和多样性的影响一直是受关注的核心问题,探明这一问题对高寒草甸的可持续发展具有重要意义。研究了封育2、7、17a的高寒矮嵩草(Kobresiahumilis)草甸的群落组分和物种多样性变化特征,并探讨了不同封育年限影响物种多样性的可能原因。结果表明:1)随封育年限延长,群落上层禾本科植物的总体重要值明显增加,其中异针茅(Stipaaliena)最为显著,其重要值封育2a(13.87%)< 封育7a(21.76%)< 封育17a(23.95%),而豆科植物总体重要值显著降低,莎草科和杂类草植物变化不明显;2)α多样性:随封育年限延长,高寒草甸群落物种丰富度指数(Patrick和Margalef指数)没有发生显著变化,然而物种多样性指数(Simpson、Shannon-Wiener和种间相遇机率指数)及均匀度指数(Alatalo和Pielou指数)均表现为封育2a > 封育7a > 封育17a,且封育2a与封育7、17a呈显著性差异(P<0.05);3)β多样性:Cody指数和Morisita-Horn相似指数的变化表明,封育2a与封育7、17a之间的物种差异较大,而封育7a和17a之间的物种差异较小;4)封育后凋落物的覆盖时间及其分解引起的土壤养分变化是导致物种多样性降低的重要原因之一,土壤碳氮比(C/N)与物种均匀度指数(Alatalo指数)和多样性指数(Simpson指数和种间相遇机率指数)均呈显著性负相关关系(P<0.05),说明土壤养分元素之间的耦合效应对物种多样性变化具有重要影响。本研究说明,虽然物种丰富度指数随封育年限的延长没有变化,但物种均匀度和多样性指数均降低,且封育前期对物种多样性的影响大于后期,就物种多样性而言,该高寒草甸不宜进行长期封育。该研究以期为高寒草甸生物多样性的保护和可持续利用及管理提供科学依据。
青藏高原;丰富度指数;均匀度指数;多样性指数;β多样性
青藏高原的植被类型多样、植物种类丰富,是我国生物多样性最为丰富的地区之一,也是我国生物多样性的重要保护基地。其中高寒草甸是最主要的植被类型,其生态系统对气候变化的响应异常敏感[1]。高寒草甸不仅在畜牧业可持续发展中占有十分重要的地位,而且在区域气候调节、水源涵养、生物多样性保护和碳源/汇等方面均发挥着不可替代的作用[2]。然而,由于高原生态环境的脆弱性,在气候变化、超载放牧和生物资源的不合理开发利用等自然和人为因素影响下,导致高寒草甸的退化,并引起群落组分和物种多样性的改变以及生产力下降等一系列生态问题[3]。据统计,高原上约有0.45×108hm2草地出现了不同程度的退化,占青藏高原草地总面积的1/3,对高寒草甸生态系统的可持续发展造成了严重的威胁[4]。因此,保护高寒草甸的生物多样性、生态系统的稳定性及其服务功能已引起科学界和政府的高度重视。
围栏封育作为恢复草地生态系统最主要的措施之一而被广泛采用[5],封育不仅可以改变草地群落的覆盖度、生物量等数量特征,同时也改变了其土壤理化性质和外界环境特征[6]。物种多样性是评价草场封育效果的重要指标,通常封育可以避免外界干扰,使退化草场植物在一定时间内得以繁衍,使物种多样性增加,植物群落逐渐恢复[7]。但是,封育在草地恢复过程中也有不利的一面,因为封育本身也是一种人为干扰,消除了大型食草动物对草地的采食和践踏,切断了封育草地与外界生态系统能量与物质交换的通道[8]。因此,探明封育对草地生态系统群落结构和物种多样性的影响已成为近年的研究热点,这对于准确评估封育年限效果与草地的可持续利用具有重要意义。
封育年限是影响草地群落组分和物种多样性变化的重要因子,国内外学者对此做了大量的研究和报道[9- 13],但由于草地类型、气候、土壤等差异,其结果也不尽相同。对我国温性草原的研究表明,随围封年限的延长物种多样性呈逐渐增加趋势[9];但也有研究指出,物种多样性随封育年限的延长呈“增加→降低→增加”或“降低→增加→降低”的变化[10, 11]。对温带沙化草地的研究结果表明,随着围封年限的增加,呼伦贝尔沙化草地的群落结构逐渐趋于合理,物种丰富度、群落均匀性和多样性均呈增加趋势[12];而科尔沁沙化草地的物种多样性指数则随封育时间延长呈现下降趋势,即封育不利于物种多样性的维持[13]。因此,封育年限对草地生态系统群落结构和物种多样性的影响非常复杂,不仅表现在不同类型草地之间的差异,即使对同一类型草地也有不同的结果。
在高原极端环境影响下形成的高寒草甸生态系统极其脆弱,对气候变化和人为干扰非常敏感[1]。然而,就封育年限对高寒草甸群落组分和物种多样性影响的研究相对匮乏,虽然近年有一些关于围封对高寒草地群落影响的报道[14- 17],但其中大多以短期封育为主。青藏高原幅员辽阔,草地和气候类型迥异,结果也不尽相同,更缺少长期封育对高寒草甸生物多样性影响的研究。本研究在位于青藏高原东北隅的中国科学院高寒草甸生态系统定位站,对封育2、7、17a的高寒草甸群落结构、物种多样性、凋落物和土壤养分等进行了调查与测定,其主要目的是:1)揭示不同封育年限下的高寒草甸群落组分及物种多样性状况;2)探讨封育年限对高寒草甸物种多样性的影响及其主要原因,以期为保护高寒草甸在围栏封育过程中的生物多样性以及畜牧业的可持续发展提供科学依据。
1 材料与方法
1.1研究区概况
实验地位于中国科学院海北高寒草甸生态系统定位站(简称海北站),地理位置为37°29′—37°45′N,101°12′—101°23′E,海拔3200—3600 m,地处青藏高原东北隅祁连山北支冷龙岭东段南麓坡地的大通河河谷。该区位于亚洲大陆腹地,属明显的高原大陆性气候,东南季风及西南季风微弱。受高海拔条件影响,该区辐射强,气温低,无明显四季之分,仅有冷暖季之别,干湿季分明。年平均气温 -1.7 ℃,降水量约560 mm,其中降水量的80%以上集中在植物生长季的5—9月,年均实际日照时数为2467.7 h,可照日照时数为4434.7 h[18]。
实验区地势开阔平坦,植被类型为高寒矮嵩草草甸,物种分布均匀,在垂直结构上一般分上下两层[19],上层包括禾草类和较高类杂草,优势种有异针茅(Stipaaliena)、山地早熟禾(Poaorinosa)、羊茅(Festucaovina)等;下层包括莎草类和较低类杂草,优势种为矮嵩草(Kobresiahumilis)、麻花艽(Gentianastraminea)等,土壤为草毡寒冻雏形土(Mat-Cryic Cambisols)。
1.2研究方法
在海北站高寒矮嵩草草甸选取3块试验样地进行封育,至2013年底,封育年限分别为2、7、17a,面积分别为650、6000 m2和900 m2,样地之间相隔约300 m。于2013年8月底,在不同封育年限样地的观测区内随机选取5个50 cm × 50 cm的样方,将样方分为100个5 cm × 5 cm小方格,采用针刺法测量样方内各植物种的高度和盖度,再将样方内植物分种齐地面剪掉,并收集各样方内凋落物,分别放入纸袋,带回实验室烘干称重,统计各物种的生物量。然后清除样方内表层土,使用直径1.5 cm土钻分别对不同封育年限样地0—10 cm土壤进行取样(每个样地5个重复),放入自封袋保存,带回实验室风干,分别采用重铬酸钾容量法、半微量凯氏定氮法和硫酸-高氯酸消煮法测定土壤有机碳(SOC)、全氮(STN)和全磷(STP)含量。
1.3计算方法
本文计算物种重要值、α和β多样性,其中α多样性包括丰富度指数、多样性指数和均匀度指数,β多样性包括Whittaker 指数、Cody指数和Morisita-Horn相似指数,具体计算公式如下:
(1) 物种重要值[20]
IV=(RB+RC+RH)×100/3
式中,RB为相对地上生物量,即单种植物地上生物量占所有物种地上生物量和的比例;RC为相对盖度,即单种植物盖度占所有物种盖度和的比例;RH为相对高度,即单种植物自然高度占所有物种自然高度和的比例。
(2) α多样性指数[21-22]
1) 丰富度指数
Patrick 指数(R)
R = S′
Margalef 指数(Ma)
Ma =(S′-1)/ln(B)
2) 多样性指数
Simpson 指数(D)
Shannon-Wiener 指数(H′)
种间相遇机率指数(PIE)
3) 均匀度指数
Pielou 指数(Jsw)
Jsw= H′/ln(S′)
Alatalo 指数(Ea)
式中,S′为样地样方的平均物种数;B为群落的总生物量;pi为第i种物种的相对重要值;Ni为物种i的重要值;N为物种i所在样方的各个种的重要值之和。
(3)β多样性指数[23]
1)Whittaker指数(βws)
βws= S/ma - 1
2)Cody指数(βc)
βc= [ g(H) + I(H)]/2
3) 经Wolda改进的Morisita-Horn指数(CMH)
式中,S为各封育样地出现的物种总数,ma为各样方的平均物种数;g(H)是沿生境梯度H而增加的物种数,I(H) 是沿生境梯度H而失去的物种数;pia和pib为a和b样地中第i种植物的相对重要值,Sa和Sb为a和b样地出现的物种总数。
1.4数据分析
采用SPSS17.0统计软件对数据进行统计分析。在P= 0.05显著水平上,One-way ANOVA和LSD分析方法进行差异性检验,Pearson法进行相关性检验。
2 结果与分析
2.1不同封育年限的群落组分变化
不同封育年限高寒草甸的植物群落结构和组成发生了明显变化(表1和图1)。封育2a样地共有44种植物,分属于39个属、17个科;封育7a样地有44种植物,分属于39个属、15个科;封育17a样地则41种植物,分属于35个属、15个科。从功能群组成来看,在封育2、7、17a样地的群落上层,禾本科植物占绝对优势,总体重要值分别为35.02%、34.23%和40.99%(图1)。封育2a与7a之间的群落物种数和禾本科植物的重要值均未发生明显变化;但与封育2a和7a相比,封育17a群落物种数降低了6.81%,禾本科植物重要值分别增加了17.05%和19.75%。值得注意的是,在封育2a的样地中,异针茅(Stipaaliena)与山地早熟禾(Poaorinosa)的重要值分别为13.87% 和10.53%,而在封育7a和17a样地中,异针茅的重要值分别增加到21.76%(P<0.05)和23.95%(P<0.05),山地早熟禾则分别降低到3.92%(P<0.05)和5.70%(P<0.05),即随封育年限延长,群落上层逐渐由共优势种的异针茅和山地早熟禾变为单优势种的异针茅。
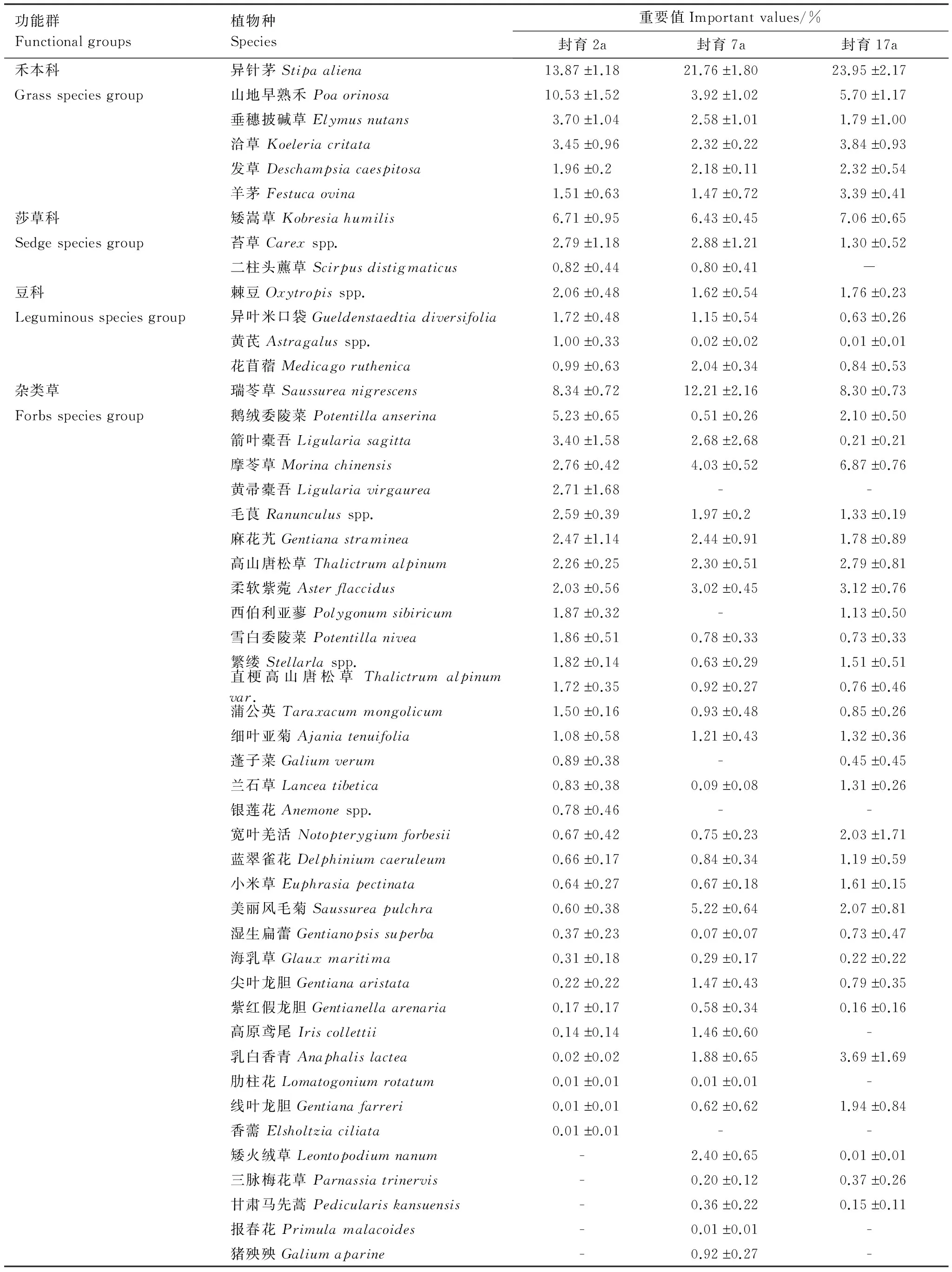
表1 不同封育年限高寒草甸群落物种组成和重要值
表中“-”表示该种植物不存在
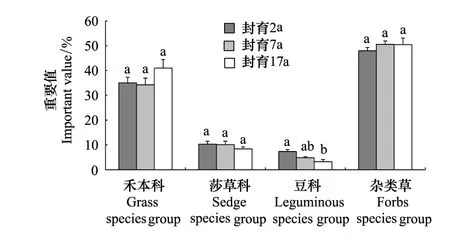
图1 不同封育年限高寒草甸群落功能群 Fig.1 Community functional group for different exclosure duration (year) in alpine meadow图中字母不同表示差异显著(P<0.05)
莎草科植物的总体重要值随封育年限延长而降低,封育2、7、17a样地分别为10.32%、10.11%和8.36%(图1),没有显著变化。下层优势种矮嵩草(Kobresiahumilis)的重要值封育2、7、17a分别是6.71%、6.43%和7.06%,未发生显著变化,即不受封育年限的影响。
豆科植物的总体重要值在封育2、7、17a样地分别是5.77%、4.82%和3.23%,封育7a和17a分别比封育2a样地降低了16.46%和44.02%(P<0.05)(图1)。
杂类草植物的总体重要值封育2、7、17a分别是47.96%、51.49%和49.53%(图1),没有发生显著变化。但其中的几种植物重要值却发生了较大变化。例如,鹅绒委陵菜(Potentillaanserina)的重要值为封育2a(5.23%)> 封育17a(2.10%)> 封育7a(0.51%);摩苓草(Morinachinensis)重要值为封育17a(6.87%)> 封育7a(4.03%)> 封育2a(2.76%);黄帚橐吾(Ligulariavirgaurea)在封育2a样地中的重要值为2.71%,但没有出现在封育7a和17a样地中;箭叶橐吾(Ligulariasagitta)的重要值为封育2a(3.40%)> 封育7a(2.68%)> 封育17a(0.21%);西伯利亚蓼(Polygonumsibiricum)的重要值在封育2a和17a样地中分别为1.87%和1.13%,而没有出现在封育7a样地;美丽风毛菊(Saussureapulchra)重要值为封育7a(5.22%)> 封育17a(2.07%)> 封育2a(0.60%);线叶龙胆(Gentianafarreri)的重要值是封育17a(1.94%)> 封育7a(0.62%)> 封育2a(0.01%)。该结果表明,虽然杂类草植物总体重要值随封育年限延长未出现显著性变化,但封育对其群落内部物种的组成及重要值产生了较大影响。
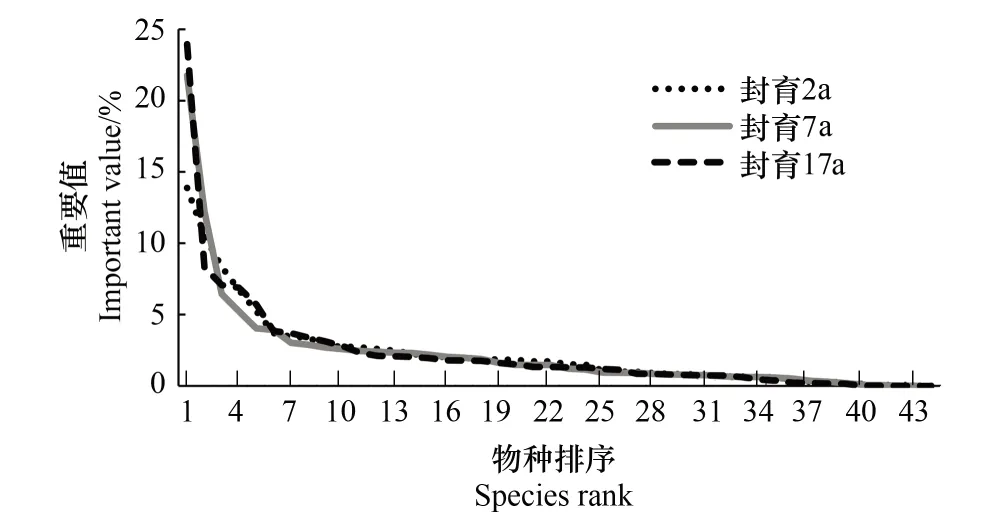
图2 不同封育年限高寒草甸群落物种重要值排序 Fig.2 Ranking of species important values for different exclosure duration (year) in alpine meadow
由物种重要值的大小排序可以发现(图2),不同封育年限的物种重要值变化大致从第10个物种开始趋于重合,而前9个物种占整体重要值的比例较大,封育2、7、17a样地分别为58.02%、62.15%和65.92%。此外,不同封育年限之间的变化趋势有所不同:封育2a样地降低速率最为缓慢,且趋势线最平滑;封育7a样地降低速率加快,但波动很小;而封育17a样地不仅降低速率加快且趋势线波动最大,其主要原因是随封育年限增加异针茅重要值迅速增加,而山地早熟禾则呈相反变化。
随封育年限的延长,凋落物量呈下降趋势,但差异不显著(图3);群落上层重要值最大的异针茅相对高度变化由封育2a的6.11%增加到封育7a和17a的8.13%和7.46%(P<0.05)(7a与17a之间不显著)(图3)。
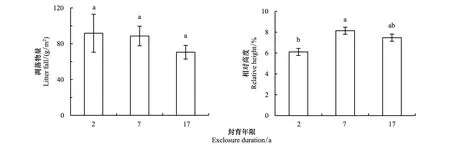
图3 不同封育年限高寒草甸凋落物量和异针茅相对高度Fig.3 Litter fall and relative height of Stipa aliena for exclosure duration (year)图中字母不同表示差异显著(P<0.05)
2.2不同封育年限的物种多样性2.2.1不同封育年限的α多样性
在不同封育年限样地中,表示物种丰富度的Patrick指数(R)和Margalef指数(Ma)(图4)均未发生显著性变化,3个封育年限样地的R和Ma分别在34和7.7左右,说明封育年限对物种丰富度并未造成明显影响。然而,表示物种均匀度的Pielou指数(Jsw)和Alatalo 指数(Ea)(图4)均为封育2a > 封育7a > 封育17a。其中,封育2a与封育17a样地之间的Jsw差异显著(P<0.05),而封育7a与另外两样地的差异不显著;Ea则随封育年限延长呈显著性降低(P<0.05)。
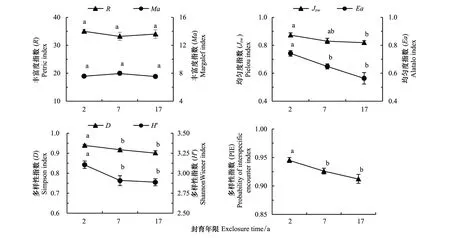
图4 不同封育年限高寒草甸群落α多样性Fig.4 α diversity of alpine meadow for different exclosure duration (year)图中小写字母不同表示差异显著(P<0.05)
表示物种多样性的Simpson指数(D)、Shannon-Wiener 指数(H′)(图4)和种间相遇机率指数(PIE)(图4)均为封育2a> 封育7a> 封育17a,而封育2a样地显著高于封育7a和17a样地(P<0.05),但封育7a和17a样地之间的差异不显著。
2.2.2不同封育年限的β多样性
不同封育年限的Morisita-Horn(CMH)指数和Cody(βc)指数表明(表2),封育2a和7a样地的CMH值居中(0.85),βc值最大(5.0),封育7a较封育2a增加和减少的物种数均为5;封育2a和17a的CMH值最小(0.83),βc值居中(4.5),封育17a较封育2a增加和减少的物种数分别为3和6;封育7和17aCMH值最大(0.94),βc值最小(3.5),封育17a较封育7a增加和减少的物种数分别为2和5,说明封育2a与7、17a样地之间的物种差异较大,而封育7a和17a样地之间的物种差异相对较小。
封育2、7、17a样地的Whittaker(βws)指数分别为0.26、0.33和0.21(图5),说明封育2a样地群落内物种更替程度居中,封育7a样地物种更替程度最大,样方间种类组成相似性最小;封育17a则物种更替程度最小,样方间种类组成相似性最大。β多样性指数综合结果说明,物种多样性对封育年限的响应程度在封育前期(封育2和7a之间)大于封育后期(封育7和17a之间),这与α多样性的结果相一致。

表2 不同封育年限之间高寒草甸Cody指数(βc)和Morisita-Horn(CMH)相似指数
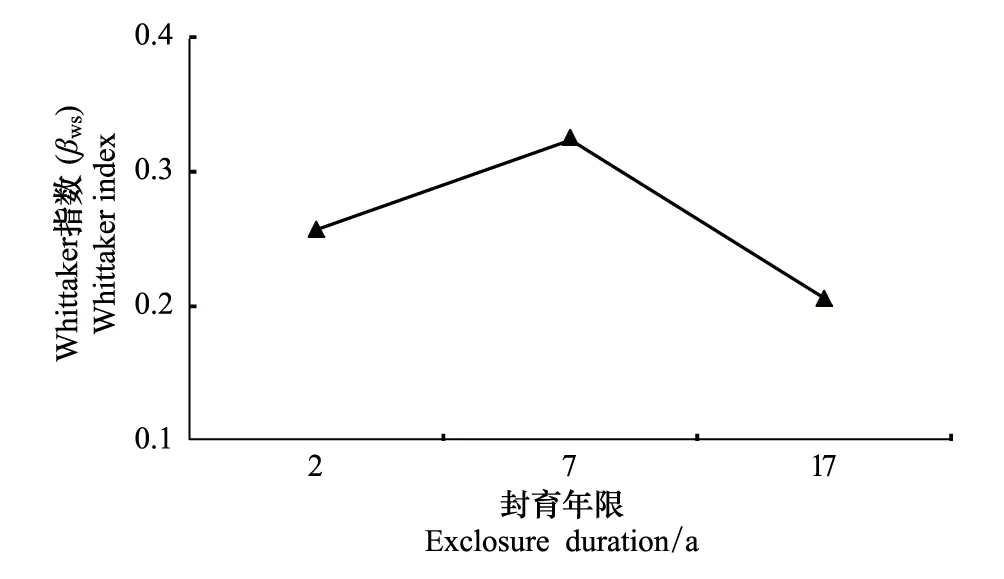
图5 不同封育年限高寒草甸Whittaker指数(βws) Fig.5 Whittaker index for different exclosure duration in alpine meadow
2.2.3不同封育年限的物种多样性相关性
丰富度指数(Ma)与Whittaker指数(βws)呈显著正相关关系(P<0.05)(图6),多样性指数(D、H′和PIE)随均匀度指数(Jsw和Ea)的增加呈显著性上升趋势(P<0.05)(图6)。
2.3不同封育年限的土壤养分与物种多样性相关性
封育2、7、17a的SOC、STN、STP含量以及土壤氮磷比(N/P)均表现为封育17a > 封育2a > 封育7a,土壤碳氮比(C/N)没有发生显著变化(表3)。SOC、STN、STP含量与α和β多样性指数的相关性均不显著,土壤C/N与多样性指数和均匀度指数呈负相关关系,其中均匀度指数(Ea)和多样性指数(D和PIE)与C/N均达到显著关系(P<0.05)(图7),物种均匀度和多样性指数随土壤N/P的增加呈降低趋势(图7)。
3 讨论与结论
3.1凋落物对群落组分和物种多样性的影响
凋落物的积累与分解是影响植被群落组分和生态系统物种多样性的重要因子,包括凋落物的量和覆盖时间两方面[24-25]。凋落物量增加是草地封育初期最显著的特征之一,但随封育年限延长,凋落物量呈下降趋势[26-27],然而也有相反结果的报道[28]。本研究与左万庆等[26]和康博文等[27]研究结果一致,即随封育年限的增加凋落物量呈下降趋势(图3),但由于3个不同封育年限样地之间的凋落物量差异并不显著,故认为不同封育年限群落组分和物种多样性的改变与凋落物量无明显关系。Xiong等[25]也指出凋落物覆盖的时间越长,对物种多样性的负效应越大,而凋落物量的影响并不显著。本研究中,凋落物覆盖对不同功能群植物的影响不同,莎草科植物受影响最小,豆科植物则最大,这与Wang等[29]的研究结果一致。本研究结果亦表明,均匀度指数和多样性指数随封育年限延长而下降,其主要原因可能与凋落物覆盖时间有关。通常,覆盖的时间越长,凋落物阻止种子到达土壤表面的可能性越大,从而抑制种子萌发和幼苗建植[30],降低物种多样性。另外,凋落物覆盖极大地消弱了群落底层的光照强度,导致一些喜光植物随封育年限延长,其生存空间逐渐缩小而被淘汰,而光竞争能力强或耐荫的物种得以存留。因此,由于凋落物覆盖时间的负效应影响,最终导致群落组分发生变化、物种均匀度指数和多样性指数随封育年限延长而降低。
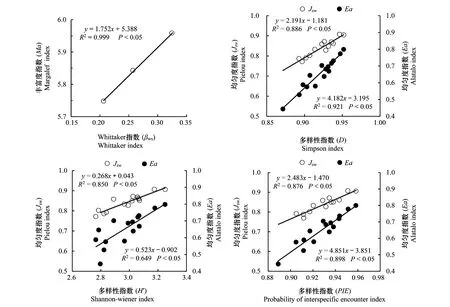
图6 不同封育年限高寒草甸群落物种多样性相关性Fig.6 Correlations of species diversity for different exclosure duration (year) in alpine meadow

封育年限/aExclosureduration土壤养分含量Soilnutrientcontent/(g/kg)土壤有机碳(SOC)Soilorganiccarbon土壤全氮(STN)Soiltotalnitrogen土壤全磷(STP)Soiltotalphosphorus土壤碳氮比(C/N)SoilC/Nratio土壤氮磷比(N/P)SoilN/Pratio254.50±0.90b4.77±0.13a0.79±0.03a11.45±0.25a6.60±0.19b750.46±1.25b4.29±0.20b0.69±0.02b11.78±0.32a6.48±0.23b1771.34±1.25a5.95±0.18a0.80±0.02a12.01±0.29a7.89±0.24a
表中字母不同表示差异显著(P<0.05)

图7 不同封育年限高寒草甸群落土壤养分比与多样性的关系Fig.7 Relationship between soil nutrient ratio and species diversity for different exclosure duration in alpine meadow图中没有标明P值表示相关性不显著(P>0.05)
然而,物种丰富度指数并没有随封育年限增加发生显著变化,这可能与该高寒草甸存在一定的物种冗余有关,因为该群落存在许多重要值特别低(表1)且对初级生产力贡献很小的物种[31],并且研究样地在0.25 m2样方内平均物种数目是内蒙古贝加尔针茅群落、大针茅群落、克氏针茅群落和小针茅群落物种数[32]的1.88—4.67倍。根据“冗余种假说”,该生态系统可能存在一些物种能够适应因封育而改变的微环境。例如,群落下层优势种矮嵩草,不同封育年限之间并没有显著变化;另外,美丽风毛菊较耐荫,单位面积叶片的叶绿素含量和PSⅡ有效光化学量子产量要高于其它植物[33],叶片在地表伸张使其在与其它植物的竞争中占据更多的地表空间,其重要值封育7a和17a均显著大于封育2a(P<0.05)。
3.2光竞争对群落组分和物种多样性的影响
光照条件对草地植物群落的生长有重要作用[34],物种对光资源的竞争是影响植被群落结构和多样性变化的因素之一[25]。根据Tilman[35]的资源比例假说,草地植物群落结构的改变与透光率有关,低透光率导致低的物种存活率。虽然本研究没有对群落内的光照强度进行观测,然而植株高度是反映植物对光资源竞争能力重要的代表性指标之一[31]。在本研究中,随封育年限的延长,群落上层重要值最大的异针茅相对高度变化由封育2a的6.11%增加到封育7a和17a的8.13%和7.46%(P<0.05)(7a与17a之间不显著)(图3B)。由于异针茅的光竞争能力较强[36],其重要值(表1)和相对高度随封育年限延长而增加,减弱其群落下层的光照,遮蔽了其他物种,其最终结果导致即长不高又不耐荫的物种被排除掉,使物种多样性下降,这与Chapin等[37]报道的结果一致。
3.3土壤养分对群落组分和物种多样性的影响
土壤养分是维系地上和地下生物活动最基本的营养元素,也是影响群落组分和生物多样性的重要因子[38],而封育后改变了地上植物与土壤之间的相互作用过程。多数研究表明,土壤养分随封育年限的延长而增加[39,40],然而文海燕等[41]则指出,不同封育年限之间的土壤有机质和全氮的差异均不显著。本研究中,封育2、7、17a SOC、STN和STP含量均表现为封育17a > 封育2a > 封育7a(表3),这与邱莉萍等[42]报道的半干旱地区的结果相似,其原因可能是草地封育后随外界干扰的消失,植被加速了对原有土壤营养的利用,导致土壤养分降低[42],但随封育时间的延长,由于凋落物的累积与分解导致土壤养分含量逐渐增加,因为土壤养分供应和养分循环的正反馈作用是通过凋落物的累积而加强[43]。
然而,土壤养分与群落组分和物种多样性之间的关系较为复杂,因为不同功能群植物对土壤养分变化的响应不同[44]。杨晓霞等[36]和沈振西等[45]的研究表明,N和P元素均是高寒草甸植物生长重要的限制性营养元素,因此植物对土壤养分资源的竞争强度大,不同封育年限间土壤养分的变化必定引起群落组分的改变。本研究不同功能群总体重要值的变化表明,禾本科植物变化较大,如异针茅、羊茅对土壤养分变化的响应较为敏感(表1),这可能是由于它们对养分利用率较高的原因[45-46];莎草科和杂类草植物变化不明显;而豆科植物却显著降低。土壤养分对物种多样性的影响亦存在争议,有研究表明随土壤养分的增加,物种多样性降低,但也有先增加后降低,或二者之间没有相关性的报道[44]。本研究表明, SOC、STN、STP含量与α和β多样性指数的相关性均不显著,与左小安[47]的研究结果相同。然而,土壤C/N与多样性指数和均匀度指数呈负相关关系,其中均匀度指数(Ea)和多样性指数(D和PIE)与C/N均达到显著关系(P<0.05)(图7),说明物种多样性主要是通过养分因子之间的耦合效应起作用,可能与土壤矿化作用有关,这与Small[48]的研究结果一致。另外,本研究表明物种均匀度和多样性指数随土壤N/P的增加呈降低趋势(图7),可能原因是N元素的增加导致优势植物快速生长,进而对其它物种竞争排斥,导致物种多样性降低,这与Roem[49]的研究结果一致。
3.4其他因素的可能影响
影响群落物种多样性变化的因子非常复杂,是生物多样性研究的主要内容。这些影响因子主要分为生物因子和环境因子,如光照、温度、水分、养分、干扰以及物种之间的相互关系, 而群落生境差异可能是影响多样性的主要原因,当前研究最多的环境因子是土壤因子和光照因子[50]。围封年限的不同对其生境的改变亦不相同,对于天然草地,围封后凋落物量、覆盖时间及其分解对土壤因子和光照因子的改变是影响群落生境最显著的因素,前文对此行了探讨。然而,围封引起的其他因子变化也可能对生物多样性产生影响,如土壤水分、温度以及物种间的相互作用等。
水分是影响植物生长发育及生物多样性的重要因子,尤其是在干旱地区[51]。本研究3个围封样地相邻,降水量相同,但封育样地中的凋落物覆盖可能通过截留部分降水、或减少土壤蒸发而改变水分收支和土壤水分[30]。然而,该研究样地年均降水量达560 mm,特别是生长季中水分相对丰富,有研究指出水分不是该生态系统的限制因素[52]。另外,3个样地间凋落物量无显著差异(图3),故可认为凋落物对降水的截留不会成为影响生物多样性的直接原因。
温度是控制植物生理活动和生化反应最重要的生态因子之一,能限制植物的分布,从而影响生物多样性。气候变暖对全球的生态系统和生物多样性产生着显著影响[53],特别是受低温条件控制的高山生态系统对气候变暖的响应尤为敏感[54],相关研究主要集中在全球尺度或较大的区域尺度上。围封后由于凋落物覆盖而改变群落底部温度等微环境,进而影响生物多样性,但目前尚未见到相关研究报道。本研究的3个样地温度背景相同,且样地间凋落物量无显著差异(图3),但对于不同封育年限,凋落物覆盖是否可导致样地间的温度差异及其对生物多样性产生的影响仍有待于进一步探讨。
极限相似性理论认为物种间相互作用可引起群落性状趋异的构建模式[55],从而影响群落物种多样性。封育后,凋落物的增加,牲畜采食、践踏等干扰的排除,都是引起群落结构和土壤理化性质发生改变的重要因素,很多研究也报道了围封对群落结构和土壤理化性质的影响[6,9- 15]。总之,围封引起群落结构和土壤性质的改变使原有种间关系发生变化,也改变了群落中共存物种对资源的利用状况,最终可能导致物种多样性的改变。然而,物种间的相互作用关系非常复杂,欲探明其种间关系对生物多样性的影响还需长期的、更深入的实验数据提供依据。
3.5结论
本文探讨了2、7、17a不同封育年限对青藏高原东北部高寒草甸群落组分及物种多样性的影响,结果表明,随封育年限的延长,禾本科植物的总体重要值增加最明显,特别是异针茅最为显著,成为17a封育样地中群落上层的单优势种;而豆科植物总体重要值显著降低;莎草科和杂类草植物变化不明显,说明封育对群落的物种组成及重要值具有较大影响。物种多样性指数和均匀度指数均随封育年限的延长呈下降趋势,且封育前期(2a与7a之间)变化大于后期(7a与17a之间)。凋落物的存在影响植物的种子萌发、幼苗定居以及物种间的相互关系,但3个样地间凋落物量的差异并不显著,说明不同封育年限样地间物种多样性的差异与凋落物量无关,而与凋落物覆盖时间有关。随封育年限的延长,凋落物分解参与调节的土壤养分含量及其比例发生改变,其中土壤C/N和N/P与群落物种多样性呈负相关关系,说明土壤养分之间的耦合作用影响高寒草甸群落组分及其物种多样性。本研究说明,封育虽然是草地恢复的有效措施之一,但就封育年限对物种多样性的影响而言,该地区高寒草甸生态系统不宜进行长期封育,可采取适度的放牧或轮牧管理措施,保护其物种多样性。由于影响物种多样性变化的因子非常复杂,在以后的研究中还需进一步加强研究封育年限对群落结构及其生物多样性影响的生态学机制,同时加强封育年限及其生物多样性变化对生态系统功能影响的研究。
[1]徐玲玲, 张宪洲, 石培礼, 于贵瑞. 青藏高原高寒草甸生态系统表观量子产额和表观最大光合速率的确定. 中国科学(D辑: 地球科学), 2004, 34(S2): 125- 130.
[2]孙鸿烈, 郑度, 姚檀栋, 张镱锂. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3- 12.
[3]王长庭, 龙瑞军, 王启兰, 曹广民, 施建军, 杜岩功. 放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应. 生态学报, 2008, 28(9): 4144- 4152.
[4]马玉寿, 郎百宁, 王启基.“黑土型”退化草地研究工作的回顾与展望. 草业科学, 1999, 16(2): 5- 9.
[5]韩龙, 郭彦军, 韩建国, 郭芸江, 唐华. 不同刈割强度下羊草草甸草原生物量与植物群落多样性研究. 草业学报, 2010, 19(3): 70- 75.
[6]乔春连, 王基恒, 葛世栋, 陈懂懂, 赵亮, 李英年, 徐世晓. 围封和放牧条件下高寒矮嵩草草甸土壤性质的比较. 草业科学, 2012, 29(3): 341- 345.
[7]姜世成, 周道玮. 过牧、深翻及封育三种方式对退化羊草草地的影响. 中国草地, 2002, 24(5): 5- 9.
[8]江小蕾, 张卫国, 杨振宇, 王刚. 不同干扰类型对高寒草甸群落结构和植物多样性的影响. 西北植物学报, 2003, 23(9): 1479- 1485.
[9]吴建波, 包晓影, 李洁, 赵念席, 高玉葆. 不同围封年限对典型草原群落及大针茅种群特征的影响. 草地学报, 2010, 18(4): 490- 495.
[10]单桂莲, 徐柱, 宁发, 马玉宝, 李临杭. 围封年限对典型草原群落结构及物种多样性的影响. 草业学报, 2008, 17(6): 1- 8.
[11]闫玉春, 唐海萍, 常瑞英, 刘亮. 典型草原群落不同围封时间下植被、土壤差异研究. 干旱区资源与环境, 2008, 22(2): 145- 151.
[12]吕世海, 冯长松, 高吉喜, 卢欣石. 呼伦贝尔沙化草地围封效应及生物多样性变化研究. 草地学报, 2008, 16(5): 442- 447.
[13]苗仁辉, 蒋德明, 王永翠. 科尔沁沙质草地封育过程中的植被变化及其机制. 干旱区研究, 2013, 30(2): 264- 270.
[14]晁增国, 汪诗平, 徐广平, 胡宜刚, 张振华. 围封对退化矮嵩草草甸群落结构和主要种群空间分布格局的影响. 西北植物学报, 2009, 28(11): 2320- 2326.
[15]苏淑兰, 李洋, 王立亚, 郭丁, 康海军, 李旭东, 傅华. 围封与放牧对青藏高原草地生物量与功能群结构的影响. 西北植物学报, 2014, 34(8): 1652- 1657.
[16]赵景学, 曲广鹏, 多吉顿珠, 尚占环. 藏北高寒植被群落物种多样性与土壤环境因子的关系. 干旱区资源与环境, 2011, 25(6): 105- 108.
[17]苗福泓, 郭雅婧, 缪鹏飞, 郭正刚, 沈禹颖. 青藏高原东北边缘地区高寒草甸群落特征对封育的响应. 草业学报, 2012, 21(3): 11- 16.
[18]李英年, 赵新全, 曹广民, 赵亮, 王勤学. 海北高寒草甸生态系统定位站气候、植被生产力背景的分析. 高原气象, 2004, 23(4): 558- 567.
[19]周华坤, 周立, 刘伟, 王启基, 赵伟, 周彦艳. 封育措施对退化与未退化矮嵩草草甸的影响. 中国草地, 2003, 25(5): 17- 22.
[20]周杰, 赵洪, 袁晓霞, 杨玉婷, 罗开嘉, 郭正刚. 青藏高原边缘区高寒草甸植物群落的特征. 草业科学, 2013, 30(7): 1066- 1070.
[21]陈芙蓉, 程积民, 刘伟, 朱仁斌, 杨晓梅, 赵新宇, 苏纪帅. 不同干扰对黄土区典型草原物种多样性和生物量的影响. 生态学报, 2013, 33(9): 2856- 2866.
[22]刘维暐, 王杰, 王勇, 杨帆. 三峡水库消落区不同海拔高度的植物群落多样性差异. 生态学报, 2012, 32(17): 5454- 5466.
[23]白永飞, 邢雪荣, 许志信, 李德新. 内蒙古高原针茅草原群落β多样性研究. 应用生态学报, 2000, 11(3): 408- 412.
[24]Wardle D A, Bonner K I, Nicholson K S. Biodiversity and plant litter: experimental evidence which does not support the view that enhanced species richness improves ecosystem function. Oikos, 1997, 79(2): 247- 258.
[25]Xiong S J, Nilsson C. The effects of plant litter on vegetation: a meta-analysis. Journal of Ecology, 1999, 87(6): 984- 994.
[26]左万庆, 王玉辉, 王风玉, 师广旭. 围栏封育措施对退化羊草草原植物群落特征影响研究. 草业学报, 2009, 18(3): 12- 19.
[27]康博文, 刘建军, 侯琳, 李胜功. 蒙古克氏针茅草原生物量围栏封育效应研究. 西北植物学报, 2006, 26(12): 2540- 2546.
[28]阚雨晨, 王堃, 王宇通, 黄欣颖, 邵新庆. 典型草原封育过程中植物凋落物的变化动态. 草原与草坪, 2011, 31(4): 25- 29.
[29]Wang C T, Long R J, Wang Q L, Liu W, Jing Z C, Zhang L. Fertilization and litter effects on the functional group biomass, species diversity of plants, microbial biomass, and enzyme activity of two alpine meadow communities. Plant and Soil, 2010, 331(1/2): 377- 389.
[30]李强, 周道玮, 陈笑莹. 地上枯落物的累积、分解及其在陆地生态系统中的作用. 生态学报, 2014, 34(14): 3807- 3819.
[31]李晓刚. 模拟放牧和土壤资源获得性对高寒草甸物种多样性、功能多样性与初级生产力关系的影响[D]. 西安: 陕西师范大学, 2012.
[32]白永飞, 许志信, 李德新. 内蒙古高原针茅草原群落α多样性研究. 生物多样性, 2000, 8(4): 353- 360.
[33]师生波, 李惠梅, 王学英, 岳向国, 徐文华, 陈桂琛. 青藏高原几种典型高山植物的光合特性比较. 植物生态学报, 2006, 30(1): 40- 46.
[34]Tilman D. Species richness of experimental productivity gradients: how important is colonization limitation? Ecology, 1993, 74(8): 2179- 2191.
[35]Tilman D. The resource-ratio hypothesis of plant succession. The American Naturalist, 1985, 125(6): 827- 852.
[36]杨晓霞, 任飞, 周华坤, 贺金生. 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应. 植物生态学报, 2014, 38(2): 159- 166.
[37]Chapin III F S, Shaver G R, Giblin A E, Nadelhoffer K J, Laundre J A. Responses of arctic tundra to experimental and observed changes in climate. Ecology, 1995, 76(3): 694- 711.
[38]Huang P M, Wang S L, Tzou Y M, Huang Y B, Weng B Q, Zhuang S Y, Wang M K. Physicochemical and biological interfacial interactions: impacts on soil ecosystem and biodiversity. Environmental Earth Sciences, 2013, 68(8): 2199- 2209.
[39]白于. 黄土区不同封育时期天然草地根系与土壤特性[D]. 杨凌: 西北农林科技大学, 2014.
[40]何玉惠, 赵哈林, 刘新平, 赵学勇, 李玉强. 封育对沙质草甸土壤理化性状的影响. 水土保持学报, 2008, 22(2): 159- 161, 181- 181.
[41]文海燕, 赵哈林, 傅华. 开垦和封育年限对退化沙质草地土壤性状的影响. 草业学报, 2005, 14(1): 31- 37.
[42]邱莉萍, 张兴昌, 程积民. 不同封育年限草地土壤有机质组分及其碳库管理指数. 植物营养与肥料学报, 2011, 17(5): 1166- 1171.
[43]Aerts R, de Caluwe H. Nutritional and plant-mediated controls on leaf litter decomposition ofCarexspecies. Ecology, 1997, 78(1): 244- 260.
[44]王长庭, 龙瑞军, 刘伟, 王启兰, 张莉, 吴鹏飞. 高寒草甸不同群落类型土壤碳分布与物种多样性、生物量关系. 资源科学, 2010, 32(10): 2022- 2029.
[45]沈振西, 周兴民, 陈佐忠, 周华坤. 高寒矮嵩草草甸植物类群对模拟降水和施氮的响应. 植物生态学报, 2002, 26(3): 288- 294.
[46]曹广民, 杜岩功, 梁东营, 王启兰, 王长庭. 高寒嵩草草甸的被动与主动退化分异特征及其发生机理. 山地学报, 2008, 25(6): 641- 648.
[47]左小安, 赵学勇, 赵哈林, 李玉强, 郭轶瑞, 赵玉萍. 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系. 环境科学, 2007, 28(5): 945- 951.
[48]Small C J, McCarthy B C. Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. Forest Ecology and Management, 2005, 217(2/3): 229- 243.
[49]Roem W J, Berendse F. Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities. Biological Conservation, 2000, 92(2): 151- 161.
[50]汪殿蓓, 暨淑仪, 陈飞鹏. 植物群落物种多样性研究综述. 生态学杂志, 2001, 20(4): 55- 60.
[51]李新荣, 张景光, 刘立超, 陈怀顺, 石庆辉. 我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究. 植物生态学报, 2000, 24(3): 257- 261.
[52]李红琴, 李英年, 张法伟, 刘晓琴, 吴启华, 毛绍娟. 高寒草甸植被生产量年际变化及水分利用率状况. 冰川冻土, 2013, 35(2): 475- 482.
[53]Walther G R, Post E, Convey P, Menzel A, Parmesan C, Menzel A, Parmesan C, Beebee T J, Fromentin J M, Guldberg O H, Bairlein F. Ecological responses to recent climate change. Nature, 2002, 416(6879): 389- 395.
[54]Körner C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlín & Heidelberg: Springer Verlag, 1999: 297- 298.
[55]Pillar V D, da S Duarte L, Sosinski E E, Joner F. Discriminating trait-convergence and trait-divergence assembly patterns in ecological community gradients. Journal of Vegetation Science, 2009, 20(2): 334- 348.
Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet Plateau
LIU Xiaoqin1, ZHANG Xiang1, ZHANG Lifeng1, LI Yingnian2, ZHAO Liang2, XU Shixiao2, LI Hongqin2, MA Rongrong1, NIU Ben1, GAO Yubao1, GU Song1,*
1CollegeofLifeSciences,NankaiUniversity,Tianjin300071,China2NorthwestPlateauInstituteofBiology,ChineseAcademyofSciences,Xining810008,China
The Qinghai-Tibet Plateau is one of the richest areas in terms of biodiversity in China. The alpine meadow is the most widespread vegetation type on this plateau and plays an important role in the carbon cycle, conservation of biodiversity, and livestock husbandry. However, climate change and human activities have greatly affected this already fragile ecosystem. Although exclosure, one of the effective measures, has been widely applied in grassland restoration, the impact of exclosure duration on community structure and species diversity is of particular interest. Therefore, determining the influence of exclosure duration on the alpine meadow is important for the development of sustainable livestock husbandry. We examined the community structure and species diversity in patches ofKobresiahumilisalpine meadow, which had been enclosed for 2, 7, or 17 years. We have discussed possible reasons for the changes in community structure and species diversity. The results indicated the following: 1) With increase in exclosure duration, the percentage of grass species in the upper community increased significantly, especially ofStipaaliena. The percent change was 13.87% for 2-year, 21.76% for 7-year, and 23.95% for 17-year exclosure. The percent change of leguminous species significantly decreased, whereas sedge and forbs species exhibited no obvious change in exclosure duration. 2) The α-biodiversity, Patrick index, and Margalef index of the plant communities indicated no significant changes in exclosure duration. In contrast, the Simpson index, Shannon-Wiener index, probability of interspecific encounter index, Alatalo index, and Pielou index decreased in exclosure duration (2-year > 7-year > 17-year enclosures); there were significant differences between the 2-year and 7-year exclosures and between the 2-year and 17-year exclosures (P<0.05). 3) Theβ-biodiversity, Cody index, and Morisita-Horn similarity index indicated that differences inspecies were greater between 2-year and 7-year exclosures, as well as 2-year and 17-year exclosures, than that between 7-year and 17-year exclosures. 4) The amount of plant litter decreased in exclosure duration, but differences were not significant. Litter coverage time appeared to be one of the reasons for the decrease in species diversity because the litter layer could prevent the seeds from falling to the ground. 5) Soil organic carbon (SOC), soil total nitrogen (STN), and soil total phosphorus (STP) increased in exclosure duration due to litter decomposition, but we observed no effect on species diversity. We found that the ratio of soil carbon to nitrogen (C/N) showed significantly negative correlation with the Alatalo index, Simpson index, and probability of interspecific encounter index (P<0.05), indicating that the coupling relationship between soil carbon and nitrogen was possibly the most important factor influencing species diversity. Our results suggested that species diversity decreased in exclosure duration because of the effects of litter accumulation, litter decomposition, and plant competition for light. The influence of exclosure on species diversity in the early stage was greater than that in the later stage, and thus, long-term exclosure was unnecessary for species diversity in this alpine meadow. We hope that our study will provide a scientific basis for biodiversity protection, sustainable utilization, and management of alpine meadow ecosystems.
Qinghai-Tibet Plateau; richness index; evenness index; diversity index;βdiversity
国家自然科学基金项目(31070433, 31570478, 31270523, 31300385)
2015- 01- 30; 网络出版日期:2015- 12- 03
Corresponding author.E-mail: songgu@nankai.edu.cn
10.5846/stxb201501300251
刘晓琴,张翔,张立锋,李英年,赵亮,徐世晓,李红琴,马荣荣,牛犇,高玉葆,古松.封育年限对高寒草甸群落组分和物种多样性的影响.生态学报,2016,36(16):5150- 5162.
Liu X Q, Zhang X, Zhang L F, Li Y N, Zhao L, Xu S X, Li H Q, Ma R R, Niu B, Gao Y B, Gu S.Effects of exclosure duration on the community structure and species diversity of an alpine meadow in the Qinghai-Tibet Plateau.Acta Ecologica Sinica,2016,36(16):5150- 5162.
