酸枣叶表皮微形态对不同生态环境的适应特征
2016-10-25朱广龙邓荣华魏学智
朱广龙,邓荣华,魏学智
山西师范大学生命科学学院, 临汾 041000
酸枣叶表皮微形态对不同生态环境的适应特征
朱广龙,邓荣华,魏学智*
山西师范大学生命科学学院, 临汾041000
以生长于烟台—石家庄—宁夏—新疆不同生境形成的自然梯度干旱环境中的酸枣(Ziziphusjujubavar.spinosas)为材料,应用扫描电镜和能谱分析研究酸枣叶表皮微形态对不同自然梯度干旱环境的适应特征。结果表明:酸枣叶片的上、下表皮都有表皮毛着生且在叶脉处分布浓密;叶片表层覆盖有较厚的角质层和蜡质并形成突起,其主要成分为C、O、Ca三种元素;叶片表面气孔数量平均40个/视野且按一定规律分布。不同生境中酸枣叶片表面都分布有大量的晶体,且晶体的形态和组成成分表现出一定的生态适应特征。晶体的形状多样,有正六棱体、长六棱体、不规则片状、正方体、棒状晶体或针状,晶体的分布方式有单晶和簇晶。从烟台到新疆随干旱梯度的加剧,酸枣叶片表皮毛逐渐增多,气孔数量减少,气孔腔下陷明显,有助于植株减少蒸腾保水抗旱;晶体的体积逐渐变小,数量增多,密度增大,提升了叶片硬度及抗旱性;酸枣叶片晶体的主要成分为C、O、Ca三种元素,且随生境干旱加剧,C元素的含量逐渐增加,O和Ca元素含量依次减少,此外宁夏的晶体中还含有Si元素,进一步提升了叶片的机械性能。酸枣叶表皮微形态中表皮毛、角质层与蜡质、气孔、晶体等在形态结构、生物功能和组成成分上特异性的变化是其长期生存于不同梯度干旱生境的生态适应特征。
叶表皮微形态;能谱分析;晶体;气孔;生境
干旱是限制植物生长发育及生产的主要环境因子[1],植物长期生长于干旱的生境中,其生理特性和结构特征必然发生适应性的变化,并形成特异的抗旱特征。植物适应干旱胁迫的策略有两种途径:一是改变细胞代谢途径,启动各种防御机制[2- 3],如抗氧化保护酶活性的提高、渗透调节物质的积累[4];二是形态结构特征的改变,诸如根系和植株地上部分缩短节间、降低株高,叶片变小、着生腺毛、角质层加厚等[4- 9]。随着全球气候变化,区域性干旱频发,干旱直接影响植物的生态环境和分布格局[10- 11]。植物对不同生态环境条件的适应机制存在差异,研究同种植物对不同生境的适应特征及机理对评估气候变迁对植物的影响及指导生态建设恢复有重要的作用。
酸枣(ZizyphusjujubaMill.)为鼠李科(Rhamnaceae)枣属(ZizyphusMill.)植物,原产于我国,广布于华北、西北、东北和华东的向阳山坡、荒芜丘陵和平原,从湿润的海滨到干旱的荒漠均有分布[5]。酸枣根系深广,抗旱、耐寒、耐贫瘠能力强,具有重要的水土保持作用,是绿化荒山的先锋树种[12]。目前对该植物的研究主要分布在生物活性物质提取[13- 14]、药理作用探索[15]、组培嫁接[16]、抗旱生理及结构研究等方面[17- 19]。魏学智等[18]首次对酸枣叶片的结构进行了初步研究,发现其具有典型的旱生结构。贺少轩[19]、梁静[20]和朱广龙[5]等分别用野外覆膜、盆栽控水等干旱胁迫的方法研究了酸枣生理特性对干旱的响应,结果发现酸枣在生理特性上有较强的抗旱机能。康东东等[21]采用石蜡切片技术从形态结构上探究了不同地理环境中酸枣叶片的适应特征,但关于叶表皮微形态及其晶体的生态学意义,迄今尚不清楚。因此,本研究以不同生境条件下酸枣叶片为研究材料,利用扫描电镜和能谱分析技术研究不同生境条件下酸枣叶表皮形态和晶体的适应特征,首次对酸枣叶表皮晶体的组成成分和生态功能进行分析阐释,以期了解在全球气候变迁的背景下植物对不同生态环境的适应机理。
1 材料与方法
1.1采样地概况
实验材料分别采自烟台(莱山区大庄村)、石家庄(赞皇县北沟村)、宁夏(银川市贺兰山附近)、新疆(吐鲁番市红星农场三队),同时采集酸枣各个原生境土样,测定土壤含水量。烟台的莱山区大庄村(37°24′N,121°26′E)属温带海洋性季风气候,年降雨量651.9 mm,空气湿润系数为1.65,样地土壤为壤土,含水量为14%左右。石家庄赞皇县北沟村(37°42′N,114°18′E)属暖温带半湿润季风型大陆性气候,年降雨量568 mm,空气湿润系数为1.07,样地土壤为壤土,含水量为9%左右。宁夏银川市贺兰山(38°31′N,106°12′E)属中温带大陆性气候,年降雨量200 mm,空气湿润系数为0.50,样地土壤为沙土,含水量为2%左右。新疆吐鲁番市红星农场三队(42°55′N,89°11′E)属大陆性暖温带干旱荒漠气候,年降雨量15.6 mm,空气湿润系数为0.04,样地土壤为壤土,含水量为4%。样地从东到西(烟台—石家庄—宁夏—新疆)构成一个自然的梯度干旱环境(表1)。

表1 采样地区地理环境及气候特征
湿润系数是指一地降水量与蒸发量的对比关系,湿润系数越小,则该地干燥程度越明显; 各个样地的湿润系数根据, 当地气象部门提供的气象资料计算而得
1.2采样方法
2010年7月,按自然气候干旱梯度,分别从烟台(莱山区大庄村)、石家庄(赞皇北沟村)、宁夏(银川贺兰山附近)、新疆(吐鲁番红星农场三队)等地依次采样。分别选取上述样地树龄5年(依据芽鳞痕判断树龄)的酸枣植株5株,选取植物顶部往下数第3个枝条、且都靠东部方向上枝条的成熟叶片10片,共选取50张叶片摘下叶片后立即分装于(每个植株的叶片样品单独保存)FAA固定液(配方:95%酒精90 mL,福尔马林5 mL,冰醋酸5 mL)中固定保存。
1.3电镜扫描
选取固定液固定24 h以上、生长成熟、无病斑的酸枣叶片,乙醇(50%、70%、80%、90%、100%)逐级脱水,纯乙醇洗涤2—3次, 转入乙酸异戊酯中,CO2临界点干燥仪(E- 3100,英国)干燥;JFC (JFC- 1600,日本) 型离子溅射及真空镀膜仪中喷金镀膜;JSM- 7500F(日本)扫描电镜观察拍照,并对叶表面晶体及蜡质成分用能谱仪(INCA mics 04BCC,日本)进行能谱分析[8]。
1.4统计分析
每个样地随机取 30 张切片拍照(从每个样地每个植株的10个叶片中随机取 6个叶片进行拍照,共计30张切片),FSX100 OLYMPS 观察分析图像, Motic Images Plus 2.0 图像分析软件测定,然后用SPSS13.0统计软件对数据进行统计分析,SigmaPlot 10.0 软件作图, Photoshop 软件图像处理,排版。
2 结果
2.1酸枣叶片扫描电镜
电镜扫描显示,酸枣叶片上、下表皮均有表皮毛着生,上表皮毛稀少且短(图1 B),下表皮毛数量多,尤其是叶脉处表皮毛浓密较长(图1A);上表皮细胞椭圆形或近圆球形,排列紧密(图1C);下表皮气孔数量较多,且按一定的规律分布:即保卫细胞较大的气孔位于中央,周围较小的气孔围绕其呈辐射状圆环形分布(图1D);叶片上表皮覆有厚厚的角质及蜡质,蜡质在表面形成突起,成块状分布(图1E);叶片上、下表皮均分布有大量的晶体,尤其在下表皮气孔周围晶体数量较多(图1E、F),晶体的形状多样,有正六棱体(图1G)、长六棱体(图1H)、不规则片状(图1I)、正方体(图1J)、棒状晶体或针状(图1J、K),晶体的分布方式有单晶(图1K)和簇晶(图1L),单晶是晶体单个分布,簇晶是晶体聚集成簇。在扫描电镜下发现叶片下表皮气孔腔中含有大量的晶体颗粒,有的晶体排列整齐,有的摇摇欲坠,即将排出(图2)。

图1 扫描电镜下酸枣叶表皮共同特征Fig.1 The common characteristic of leaf epidermis of Ziziphus jujuba var. spinosas under scanning electron microscopeA:酸枣叶片背面主脉上着生的腺毛,×100;B:酸枣叶片正面主脉处的腺毛,×300;C:酸枣叶片正面的表皮细胞,×1000;D:酸枣叶片正面气孔的分布,×500;E:酸枣叶片表面的蜡质,×10000;F:酸枣叶片表面的气孔及晶体,×1500;G、H、I、J、K、L:酸枣叶片不同类型的晶体;G、H、×20000;L:×50000. I:K:×10000
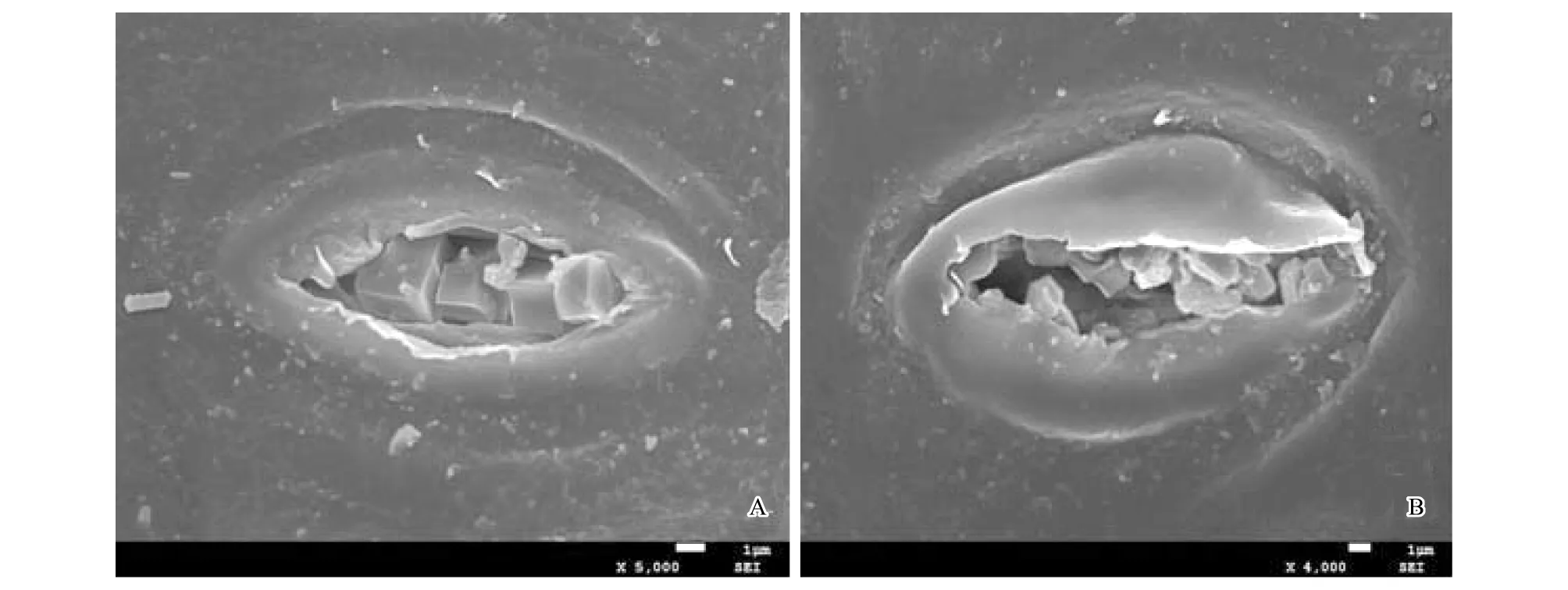
图2 酸枣叶片气孔排泄晶体,示叶表皮晶体的来源Fig.2 The source of crystals on the leaf epidermis of Ziziphus jujuba var. spinosas It showed the source of crystalsA:×5000;B:×4000
从烟台、石家庄、宁夏到新疆,随着自然干旱梯度的加剧,酸枣叶表皮毛逐渐增多(图3Y1、S1、N1、X1),气孔数量总体呈减少的趋势,其中烟台平均38.88个/视野(×300),石家庄和新疆的气孔数量与烟台相比分别减少了0.54%、8.26%,新疆的气孔数量最少,平均为35.67个/视野,宁夏的气孔数量最多,比烟台增加了15.1%(图3Y2、S2、N2、X2,图4),然而4个样地间酸枣叶表皮气孔数差异不显著(P>0.05)。气孔长度以宁夏的最大,为 514.83 um,和宁夏的相比,石家庄和新疆酸枣叶表皮的气孔长度分别减小了24.05%、26.18%(P<0.05);气孔宽度总体呈变小的趋势,烟台的气孔宽度最大,宁夏次之,新疆的最小,与烟台相比,石家庄和新疆的气孔宽度分别减小了47.13%和50.46%(P<0.05),而宁夏的减小了3.56%,差异不显著(P>0.05)(图3Y3、S3、N3、X3,图5)。保卫细胞的厚度呈先增后减的趋势,宁夏的保卫细胞厚度最大,平均为180.8μm,而新疆的最小,平均为 93.6μm,与烟台相比,石家庄和宁夏分别增加了33.57%、46.05%,新疆的减小了24.39%(P<0.05)(图3Y3、S3、N3、X3,图6)。烟台叶表皮晶体的形状主要是多面体,无针晶或棒状晶体,石家庄叶表皮中有针晶或棒状晶体的分布,但数量较少,宁夏和新疆叶片中针晶或棒状晶体分布较多,尤其新疆的叶片中,针晶聚集成簇(图1L,图3Y4、S4、N4、X4)。

图3 扫描电镜下不同地区酸枣叶表皮微形态特征Fig.3 The characteristic of leaf epidermal micromorphology of Ziziphus jujuba var. spinosas in different ecotypes under scanning electron microscopeY:烟台,S:石家庄,N:宁夏,X:新疆;Y1:烟台酸枣叶片表面腺毛,×1500;S1:石家庄酸枣叶片气孔分布,×300; N1:宁夏酸枣叶片气孔分布,×300、X1:新疆酸枣叶片气孔分布,×300;Y2:烟台酸枣叶片气孔分布,×500;S2:石家庄酸枣叶片气孔分布,×500;N2:宁夏酸枣叶片气孔分布,×500;X2:新疆酸枣叶片气孔分布,×500;Y3:烟台酸枣叶片气孔腔,×3000;S3:石家庄酸枣叶片气孔腔,×3000;N3:宁夏酸枣叶片气孔腔,×3000;X3:新疆酸枣叶片气孔腔,×3000;Y4:烟台酸枣叶片晶体形态,×20000;S4:石家庄酸枣叶片晶体形态,×20000;N4:宁夏酸枣叶片晶体形态,×10000;X4:新疆酸枣叶片晶体形态,×20000
2.2酸枣叶片能谱分析
能谱分析结果表明:酸枣叶表皮蜡质的主要成分为C、O、Ca三种元素,其中C占62.09%,是蜡质的主要组成成分,O占19.69%,Ca占18.22%(图7,表2)。烟台、石家庄、新疆3个地域中,酸枣叶片晶体的成分都为C、O、Ca三种元素(图8,图9,图10),宁夏的叶片晶体除含有C、O、Ca三种元素外,还含有少量的Si元素(图11),据此推断烟台、石家庄、新疆的酸枣叶片晶体为碳酸钙晶体或草酸钙晶体,宁夏的还含有硅质体。
烟台、石家庄、宁夏、新疆各地酸枣叶表皮晶体的成分能谱分析中,C元素的含量逐渐增多,新疆酸枣叶表皮晶体的C含量最高达到59.46%;O元素的含量逐渐减少,以烟台的含量最高为42.91%,新疆的最少为24.03%;Ca元素的含量先增后减,石家庄的Ca含量最高为47.33%,新疆的Ca含量最少为16.51%(表3)。晶体形状和组成成分的变化,是其长期适应梯度干旱的结果。

表 2 叶表皮蜡质能谱分析

表3 不同地区酸枣叶表皮晶体能谱分析

图4 不同地区酸枣叶表皮气孔数量变化Fig.4 The quantity of stomas of leaf epidermis of Ziziphus jujuba var. spinosas in different ecotypes
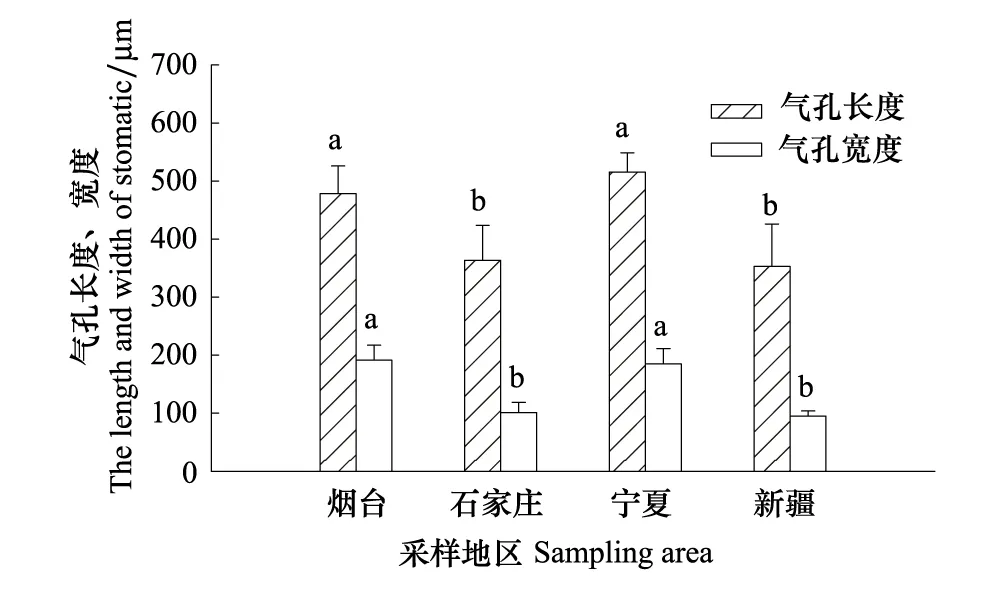
图5 不同地区酸枣叶表皮气孔长度、宽度变化Fig.5 The length and width of stomas of leaf pidermis of Ziziphus jujuba var. spinosas in different ecotypes图中误差异为SE,n=30图中误差异为SE,n=30
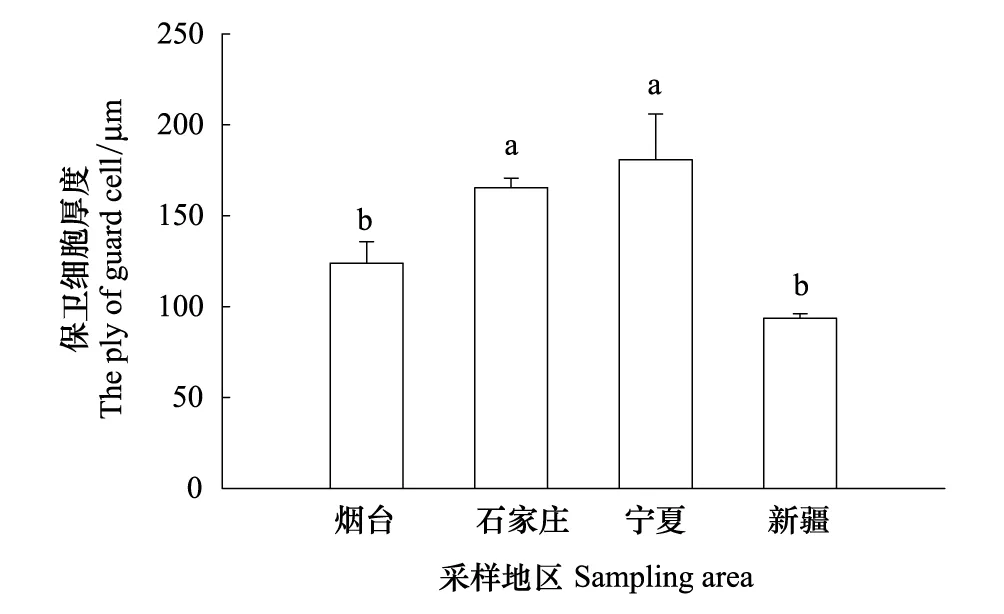
图6 不同地区酸枣叶表皮气孔保卫细胞厚度变化 Fig.6 The thickness of stomas guard cells of leaves of Ziziphus jujuba var. spinosas in different ecotypes图中误差异为SE,n=30

图7 叶表皮蜡质能谱分析Fig.7 The energy spectrum analysis for waxiness of leaf epidermis

图8 烟台叶表皮晶体能谱分析Fig.8 The energy spectrum analysis for crystal of leaf epidermis in Yantai
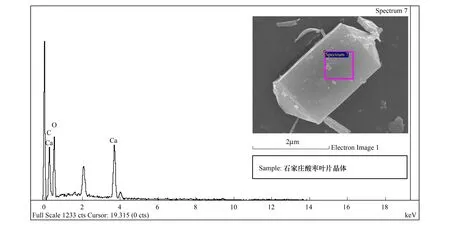
图9 石家庄叶表皮晶体能谱分析Fig.9 The energy spectrum analysis for crystal of leaf epidermis in Shijiazhuang
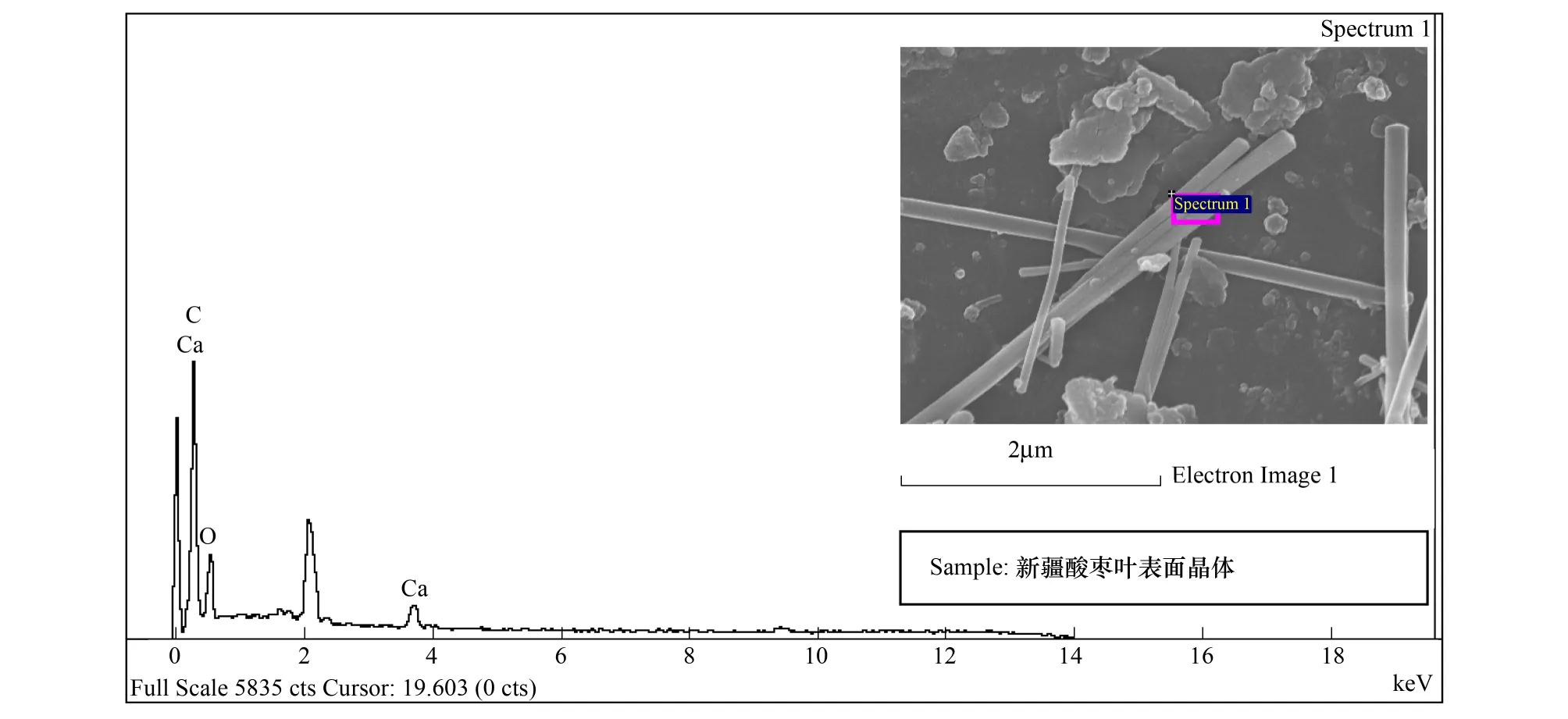
图10 新疆叶表皮晶体能谱分析Fig.10 The energy spectrum analysis for crystal of leaf epidermis in Xinjiang
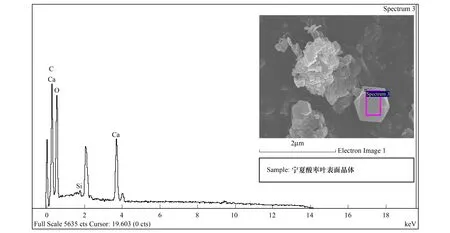
图11 宁夏叶表皮晶体能谱分析Fig.11 The energy spectrum analysis for crystal of leaf epidermis in Ningxia
3 讨论
水分是限制植物生长发育和作物生产的主要环境因子[23],干旱是植物生长过程中经常面临的逆境之一,经过长期的演化,植物可对干旱环境采取多种适应对策[24]。从烟台—石家庄—宁夏—新疆4个采样地的土壤含水量、年降雨量及湿润系数来看,其形成一个大尺度的自然梯度干旱环境的(表1),为酸枣的研究提供了一个得天独厚的条件。酸枣在长期适应梯度干旱的过程中,演化出了一些高度耐旱的特性。从烟台到新疆,四个样地的酸枣叶表皮都有表皮毛分布,叶表皮覆盖有较厚的角质层和蜡质并形成突起(图3),且随干旱程度的加剧,叶表皮表皮毛逐渐增多,角质层逐渐增厚(数据未发表,将在另一篇文章中呈现)。叶片是植物体暴露于环境中面积最大的器官,最易感知和适应环境变化而发生形态和结构上的改变,叶表皮表皮毛和厚的角质层可以有效防止水分蒸腾,是反映植物抗旱性的一个重要指标[23- 24]。赵小仙等[25]研究表明,随干旱程度的加剧,细胞角质化外壁增厚,气室变大,能有效减少水分蒸发,增加沙拐枣的抗旱性。这与本文对酸枣叶表皮的研究结果相似,角质层和蜡质加厚形成突起都是植物对不同干旱生境长期适应的结构特征。此外,坚硬的角质层除具有保水作用外,还具有机械支撑作用,使植株在水分缺乏时不会立即萎蔫[26]。酸枣叶表皮表皮毛、角质层与蜡质的变化,是其长期对不同生境的适应特征之一。
气孔是蒸腾作用的主要通道,同时也是光合作用和呼吸作用交换气体的开关,此外气孔还具有排泄功能。本研究发现,从烟台到新疆随着生境干旱程度的加剧,酸枣叶表皮气孔的纹饰变密,气孔密度增加(研究发现随干旱梯度增加,4个样地中酸枣叶片叶面积显著减小,而气孔数量减少的差异不显著,因此气孔密度增加),气孔腔由外凸逐渐下陷,气孔开度变小,保卫细胞厚度增加。
一般认为旱生植物的气孔密度增加是一种适应特征、气孔密度增加是植物对旱生环境的适应,气孔密度低是植物对湿生环境的适应,在环境干旱程度增加时植物提高气孔分化的程度,增加气孔密度,待水分供应充足时,可增加气体交换,以提高光合作用效率[27]。气孔腔下陷、凹陷的气孔可能有利于滞留CO2和H2O,在旱生条件下可为光合作用提供充足的原料,同时还可以使内在空气的饱和差值减小,阻止水分散失,形成较湿润的小环境,这可能是植物长期在逆境中产生的抗御蒸腾的机制[28];此外下陷的气孔在减少光线辐射和风沙损害方面有积极的作用[29]。气孔长宽比减小可能是为在长径一定的情况下增加气孔开口度,加大呼吸面积,从而补偿呼吸效率,增加通气量,使酸枣的气孔结构具有较强的抗逆特性[30],这与康东东等[21]对不同地理环境下酸枣叶的形态解剖特征研究结论一致。酸枣叶表皮气孔形态特征的变化,是其长期适应于不同生境的又一重要策略。
晶体是植物产生的一种具有特殊形态结构与生理功能的代谢物,晶体多集中在沙生、旱生植物的叶表皮、内部和轴器官中,被认为是荒漠植物抗旱结构特征之一, 晶体的形成可能与荒漠植物适应干旱和盐碱环境有关[22,31-32]。本研究发现酸枣的叶表皮分布有较多的晶体,且从烟台到新疆随生境干旱程度加剧,酸枣叶表皮晶体的形状由多面体向针晶过渡(图3),干旱越严重的生境晶体的体积越小,数量和密度越大。晶体可增加叶片的硬度,防止风沙损伤[33],因此宁夏和新疆生境中酸枣叶表皮晶体数量和形态特征的变化是长期适应其多风沙环境的结果。有研究表明,晶体具有较高的渗透势, 吸水能力强, 在外界环境条件适宜、植物水分输导良好时, 晶体可以吸收并贮存水分呈溶解状态;当外界环境干旱、导管中水分输导受阻而不能正常满足植物需水时,晶体呈析出状态而释放水分,为其周边细胞提供一个较为湿润的小环境,从而提高植物的抗旱性[22,33- 34]。此外还有研究表明晶体的存在一方面可加强叶片的机械性能,另一方面可避免重金属离子对细胞的毒害作用、改变细胞的渗透势、提高吸水和保水能力[33- 34]。
能谱分析表明,晶体的组成元素主要为C、O、Ca三种元素,说明该晶体主要为碳酸钙晶体或/和草酸钙晶体,且各元素的含量及比例随干旱梯度环境呈规律性的分布,C含量逐渐增加,O和Ca的含量依次减少(表3)。从化学结构上分析推测可知,C含量的增加可增大碳链的长度和不饱和程度,有利于增加晶体对水的吸水和束水能力,在宁夏和新疆极度干旱的野外生境中,酸枣叶表皮晶体在雨季可以涵养更多的水分,在旱季或逆境时将析出释放水分贡献于植株维持生命活动,从而提高植株的耐旱性。Ca含量随生境干旱加剧而依次减少的变化特征可能与各地生境的气候特征和各生境中植株各自的代谢水平有关,我国中、东南部地区工业发达环境污染较重,NO2、SO2等大量排放造成酸雨危害趋势加重[35- 38]。受酸雨等环境变化的影响,植被代谢过程中产生较多的草酸等酸性代谢物[39],此类物质对植物体会造成伤害,植株通过吸收或从晶体中析出更多的钙离子与草酸等代谢物相结合,形成中性的螯合物减轻对植物的伤害[40];而生存于宁夏和新疆等西北地区植株体内酸性代谢物少,所以酸枣植株对钙离子等碱性离子的吸收较少,则储藏于晶体中的钙离子含量就少[40]。此外,宁夏的酸枣叶表皮的晶体还有少量的Si元素,反映出宁夏酸枣叶表皮的晶体还有硅质体,硅质体的主要成份是SiO2,硅质体多为圆簇状小团块, 除氢氟酸外不溶于其它任何酸,其多存在于表皮细胞或薄壁组织中, 少数分布在纤维束周围的细胞中而类似晶鞘纤维,可增强叶片的机械性能、减少水分散失[22]。由此可见,酸枣叶表皮晶体的形态特征、类型及组成元素含量的变化是其长期适应于不同生境的又一重要适应机制。
[1]Xu S C, Ding H D, Su F X, Zhang A Y, Jiang M Y. Involvement of protein phosphorylation in water stress-induced antioxidant defense in maize leaves. Journal of Integrative Plant Biology, 2009, 51(7): 654-662.
[2]Zhu J K. Salt and drought stress signal transduction in plants. Annual Review of Plant Biology, 2002, 53: 247- 273.
[3]Boudsocq M, Lauriere C. Osmotic signaling in plants: Multiple pathways mediated by emerging kinase families. Plant Physiology, 2005, 138(3): 1185- 1194.
[4]汪攀, 陈奶莲, 邹显花, 马祥庆, 吴鹏飞. 植物根系解剖结构对逆境胁迫响应的研究进展. 生态学杂志, 2015, 34(2): 550- 556.
[5]朱广龙. 酸枣生理生化特性及结构特征对梯度干旱的响应[D]. 临汾: 山西师范大学, 2012.
[6]Brodribb T J, Holbrook N M. Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer. Plant Physiology, 2005, 137(3): 1139- 1146.
[7]Atkin O K, Macherel D. The crucial role of plant mitochondria in orchestrating drought tolerance. Annals of Botany, 2009, 103(4): 581- 597.
[8]朱广龙, 赵挺, 康冬冬, 魏学智. 木香薷腺毛形态结构发生发育规律的研究. 植物研究, 2011, 31(5): 518- 523.
[9]王顺才, 邹养军, 马锋. 干旱胁迫对3种苹果属植物叶片解剖结构、微形态特征及叶绿体超微结构的影响. 干旱地区农业研究, 2014, 32(3): 15- 23.
[10]黎祜琛, 邱治军. 树木抗旱性及抗旱造林技术研究综述. 世界林业研究, 2003, 16(4): 17- 22.
[11]赵成章, 高福元, 石福习, 任珩, 盛亚萍. 高寒退化草地甘肃臭草种群分布格局及其对土壤水分的响应. 生态学报, 2011, 32(22): 6688- 6695.
[12]王慧莉, 田涛, 王建永, Batool A, 赵旭喆, 莫非, Akram N A, 熊友才. 旱区农业雨水资源利用与生态系统可持续性: 2013干旱农业和生态系统可持续性国际会议综述. 生态学杂志, 2014, 33(11): 3127- 3136.
[13]王旭峰, 何计国, 陈阳, 李欣, 李智欣. 酸枣仁皂苷的提取及改善睡眠功效的研究. 食品科学, 2006, 27(4): 226- 229.
[14]丁轲, 张彤楠, 韩涛. 快速溶剂萃取联合高效液相色谱法测定不同产地酸枣仁中皂苷A、B的含量. 食品工业科技, 2014, 35(23): 265- 270.
[15]Barrieu F, Chaumont F, Chrispeels M J. High expression of the tonoplast aquaporinZmT1P1 in epidermal and conducting tissues of maize. Plant Physiology, 1998, 117(4): 1153- 1163.
[16]Yu S W. Plant Physiology and Molecular Biology. Beijing: Science Press, 1992.
[17]曹正其. 加强技术机制创新 提高退耕还林科技含量. 四川林业科技, 2001, 22(2): 1- 3.
[18]魏学智, 毕润成. 旱生植物酸枣的叶结构. 山西师大学报: 自然科学版, 1997, 11(3): 44- 47.
[19]贺少轩, 梁宗锁, 蔚丽珍, 周自云. 土壤干旱对2个种源野生酸枣幼苗生长和生理特性的影响. 西北植物学报, 2009, 29(7): 1387- 1393.
[20]梁静, 魏学智. 酸枣叶对土壤水分的生理生化响应. 植物研究, 2010, 30(5): 549- 555.
[21]康东东, 韩利慧, 马鹏飞, 魏学智, 毕润成. 不同地理环境下酸枣叶的形态解剖特征. 林业科学, 2008, 44(12): 135- 140.
[22]朱广龙, 马茵, 韩蕾, 霍张丽, 魏学智. 植物晶体的形态结构、生物功能及形成机制研究进展. 生态学报, 2014, 34(22): 6429- 6439.
[23]Jaleel C A, Manivannan P, Wahid A, Farooq M, Al-Juburi H J, Somasundaram R, Panneerselvam R. Drought stress in plants: a review on morphological characteristics and pigments composition. International Journal of Agriculture and Biology, 2009, 11(1): 100- 105.
[24]王勋陵, 马骥. 从旱生植物叶结构探讨其生态适应的多样性. 生态学报, 1999, 19(6): 787- 792.
[25]赵小仙, 李毅, 苏世平, 白潇. 3个地理种群蒙古沙拐枣同化枝解剖结构及抗旱性比较. 中国沙漠, 2014, 34(5): 1293- 1300.
[26]林植芳, 林桂珠. 鼎湖山植物叶片的一些与光合作用有关的结构特征 // 中国科学院华南植物研究所集刊. 北京: 科学出版社, 1989: 101- 107.
[27]Cutter E G. 植物解剖学. 李正理译. 北京: 科学出版社, 1973.
[28]胡适宜. 植物学(形态学及解剖学部分). 北京: 高等教育出版社, 1959.
[29]张道远, 张娟, 谭敦炎, 潘伯荣. 国产柽柳科3属6种植物营养枝的解剖观察. 西北植物学报, 2003, 23(3): 382- 388.
[30]李正理, 李荣敖. 我国甘肃九种旱生植物同化枝的解剖观察. 植物学报, 1981, 23(3): 181- 185.
[31]刘家琼. 我国荒漠不同生态类型植物的旱生结构. 植物生态学与地植物学丛刊, 1982, 6(4):314- 319.
[32]苏培玺, 安黎哲, 马瑞君, 刘新民. 荒漠植物梭梭和沙拐枣的花环结构及C4光合特征. 植物生态 学报, 2005, 29(1): 1- 7.
[33]李芳兰, 包维楷. 植物叶片解剖形态结构对环境变化的响应与适应. 植物学通报, 2005, 22(增刊): 118- 127.
[34]韦存虚, 张军, 王建军, 孙国荣. 星星草营养器官适应盐胁迫的结构特征. 植物资源与环境学报, 2006, 15(1): 51- 56.
[35]邵学新, 黄标, 顾志权, 钱卫飞, 金洋, 毕葵森, 严连香. 长三角经济高速发展地区土壤pH时空变化及其影响因素. 矿物岩石地球化学通报, 2006, 25(2): 143- 149.
[36]郝吉明, 段雷, 谢绍东. 中国土壤对酸沉降的相对敏感性区划. 环境科学, 1999, 20(4): 1- 5.
[37]贾曼. 石家庄污灌区土壤重金属污染现状调查及其风险评价[D]. 石家庄: 河北医科大学, 2012.
[38]赵艳霞, 侯青. 1993- 2006 年中国区域酸雨变化特征及成因分析. 气象学报, 2008, 66(6): 1032- 1042.
[39]黄晓华, 陆天虹, 周青, 曾庆玲, 梁婵娟. 酸雨伤害植物机理与稀土调控研究. 中国生态农业学报, 2004, 12(3): 116- 118.
[40]徐静静, 慈华聪, 何兴东, 薛苹苹, 赵雪莱, 郭健潭, 高玉葆. 天津盐渍化生境54种植物钙晶体与钙组分特征. 应用生态学报, 2012, 23(5): 1247- 1253.
Leaf epidermal micromorphology ofZiziphusjujubavar.spinosain response to a gradient of drought stress
ZHU Guanglong, DENG Ronghua, WEI Xuezhi*
CollegeofLifeSciences,ShanxiNormalUniversity,Linfen041000,China
Global climate change is predicted to increase average temperatures, alter geographical patterns of rainfall, and increase the frequency of extreme climatic events. Such changes are likely to alter the timing and magnitude of drought stresses experienced by plants. Drought and heat events are occurring more frequently with climate change; therefore, plants must develop morphological and physiological characteristics that confer tolerance to different drought conditions (ecotypes). The objective of this study was to explore leaf epidermal micromorphology ofZiziphusjujubavar.spinosain response to different natural drought conditions. The plant samples were collected from Yantai, Shijiazhuang, Ningxia, and Xinjiang provinces, China, which form a gradient of environmental drought according to soil moisture, annual precipitation, and humidity. We found that hairs developed in both the upper and lower epidermis, which was thicker in leaf veins and functioned to reduce evaporation from the leaf surface. Cuticles and wax composed of carbon (C), oxygen (O), and calcium (Ca) were distributed in the leaf epidermis and formed protuberances. Large numbers of stomata (approximately 40 actual unit area were arranged in regular patterns on the leaf epidermis. Numerous crystals were also observed on the epidermis in each ecotype, and the shape and composition of the crystals differed among the ecotypes. The crystals were categorized according to shape and included six-surfaced, irregularly schistose, cubic, rhabdolith, and raphide forms. Single crystals or crystal clusters were the common crystal types. As the natural drought gradient increased from Yantai to Xinjiang, the density of leaf epidermal hairs inZ.jujubavar.spinosaincreased, the number of stomata decreased, and stoma were more shrunken, which could help plants to reduce transpiration and tolerate drought. The numbers of crystals increased and crystal volume diminished, which would enhance leaf rigidity and drought resistance. Crystals mainly included C, O, and Ca; however, silica (Si) was detected in the Ningxia ecotype, in which it enhanced leaf mechanical properties. With increasing drought severity, carbon content increased but oxygen and calcium decreased. In summary, changes in morphological structure, biological function, and leaf epidermal micromorphology (including epidermal hairs, cuticles, wax, and crystals) represented adaptive ecological characteristics for long-term drought tolerance.
leaf epidermal micromorphology; energy spectrum analysis; crystal; stoma; ecotype
国家自然科学基金资助项目(30972396);山西省自然科学基金项目资助(2009011041- 1)
2015- 01- 24; 网络出版日期:2015- 11- 30
Corresponding author.E-mail: wxz3288@163.com
10.5846/stxb201501240191
朱广龙,邓荣华,魏学智.酸枣叶表皮微形态对不同生态环境的适应特征.生态学报,2016,36(16):5193- 5203.
Zhu G L, Deng R H, Wei X Z.Leaf epidermal micromorphology ofZiziphusjujubavar.spinosain response to a gradient of drought stress.Acta Ecologica Sinica,2016,36(16):5193- 5203.
