不同鸟类生境网络复合与优化——以苏锡常地区白鹭、鸳鸯、雉鸡为例
2016-10-24胡余挺范诗薇欧名豪
吴 未, 胡余挺, 范诗薇, 欧名豪,2,*
1 南京农业大学土地管理学院,南京 210095 2 农村土地资源利用与整治国家地方联合工程研究中心,南京 210095
不同鸟类生境网络复合与优化
——以苏锡常地区白鹭、鸳鸯、雉鸡为例
吴未1, 胡余挺1, 范诗薇1, 欧名豪1,2,*
1 南京农业大学土地管理学院,南京210095 2 农村土地资源利用与整治国家地方联合工程研究中心,南京210095
通过复合优化不同焦点物种的生境网络表征多数物种生境网络实现物种多样性保护,具有重要理论价值和实践意义。以苏锡常地区为研究区域,以白鹭、鸳鸯和雉鸡3种鸟类为焦点物种,通过采用生境斑块约束条件模型与最小成本路径法相结合的方法识别出生境网络;从满足集合覆盖问题视角出发,通过采用网络构成要素复合的方法进行网络复合,其中廊道与廊道的复合通过采用评价生态系统服务价值的方法实现;通过采用观测数据为生境斑块和迁移廊道增补依据的方法进行复合生境网络优化。结果表明:生境网络经过复合,廊道总面积、总长度及生态系统服务价值在分别减少了16%、68%和10%的情况下,保持了斑块间的连接度、兼顾到经济效益和生态效益最大化、覆盖了占总数86%以上的75处观测点;复合生境网络经过优化,覆盖了2010年及以后所有84处观测点,廊道总面积、总长度及生态系统服务价值分别增加了19%、21%和27%;说明生境网络复合优化定量分析方法和思路具有可行性,为网络复合研究提供了借鉴。
网络复合;网络优化;生境网络;方法论;白鹭;鸳鸯;雉鸡;苏锡常地区
生境斑块为物种生存和生物多样性保护提供了重要空间保障[1- 2]。与保护生境斑块相比,构建和保护连接生境斑块的生境网络更具有意义和价值[3- 4]。研究时可以选择单一[5- 7]或多个[8- 10]焦点物种,对识别的生境网络进行叠加,但结果不一定适用于其它物种。受诸多限制目前尚无法实现对所有甚至大多数物种的生境网络进行叠加,因此有必要通过采用对多个焦点物种生境网络复合优化的方法来表征,这在物种多样性保护研究中具有重要理论价值和实践意义。
网络复合目前还停留在理论探索与创新阶段[11]。生态网络复合可以采取网络叠加、无标度网络构建与检测相结合的方法实现[11],而生境网络复合研究还很少。网络优化可以采用扩大斑块面积、设置缓冲区、提升景观异质性[12],或者依据图论修复或增补生态断裂点和重要生境斑块[1]、暂息地[1,11,13]、生态节点[14- 15]等网络构成要素的方法实现,而采用物种分布数据增补网络构成要素的方法还较少。
本文以苏锡常地区为例,在3个焦点物种生境网络叠加的基础上,尝试通过将网络构成要素复合与观测数据检测相结合的方法,回答如何实现生境网络复合?复合效果如何等问题,以期提供一套生境网络复合优化定量分析的方法和思路,丰富网络复合研究。
1 研究区概况
苏锡常地区位于江苏省南部太湖之滨,属长江冲积平原。区内地势平坦,河湖众多,属北亚热带季风气候,年均降水1092.4 mm,年均气温15.3℃。境内物种丰富,鸟类、兽类、两栖爬行类200多种,鸟类170余种。其中,国家和省级重点保护鸟类50余种,如东方白鹳(Ciconiaboyciana)、鸳鸯(Aixgalericulata)、中华秋沙鸭(Mergussquamatus)等;涵盖了留鸟、冬候鸟、夏候鸟、旅鸟等居留类型,以及涉禽、游禽、猛禽、陆禽、攀禽、鸣禽等生态类群。白鹭在戗港、尚湖风景区(苏州)、斗山茶场、鼋头渚(无锡)、太湖湾度假区、长荡湖湿地公园(常州)等均有分布,尚湖风景区、斗山茶场、太湖湾度假区等地均出现过上百只聚集的现象;鸳鸯在大阳山森林公园、贡山岛、尚湖风景区(苏州)、鼋头渚、石塘山(无锡)、太湖湾度假区、丫髻山(常州)等均有分布;雉鸡在太湖湿地公园、沙湖生态公园、荷塘月色湿地公园、虎丘湿地公园(苏州)、斗山茶场(无锡)、丫髻山(常州)等地均有分布。
2 数据来源与研究方法
2.1数据来源
主要包括2010年苏锡常地区TM遥感数据、1∶5万数字高程模型图(DEM)、2010年苏锡常三市行政区划图以及中国观鸟记录中心(http://www.birdtalker.net/)2003—2014年观测数据。
2.2研究方法
2.2.1网络复合
生境网络复合包括生境斑块之间、生境斑块与迁徙廊道之间以及迁徙廊道之间的复合3种情形:(1)生境斑块之间的复合:当物种差异性较大、斑块间替代性较差时,可采用对所有生境斑块空间叠加的方法实现;当斑块间替代性较强时,可采用斑块优先性排序的方法实现。从集合覆盖问题(Set Covering Problem,SCP)[16]出发,(2)生境斑块与迁徙廊道之间的复合:当斑块与廊道重叠时,可将之转化为廊道优先性排序问题;当斑块与廊道不重叠时,可从斑块重要程度及网络连接度视角将之转化为网络优化问题。(3)迁徙廊道之间的复合情况最复杂。通常认为生境斑块间廊道的数量在可能范围内越多越好[17],但本文将着重探讨如何构建一条最优的潜在迁移廊道问题。
2.2.2焦点物种选择
通过焦点物种途径实现生物多样性保护是目前国际通用做法[18- 20]。焦点物种选择的原则主要包括:栖息地丧失或破碎化威胁、对其他物种及各类型栖息地的指示作用、栖息地类型是否覆盖研究区内主要栖息地类型、生物学上的代表性与典型性、是否引起公众关注[21- 23]等。鸟类在苏锡常地区种类多,栖息地类型丰富,能覆盖研究区主要生境地类,适宜作为焦点物种。
遵循以上原则,重点从鸟类生态类群、居留类型、栖息地特征、生活习性及人为干扰敏感程度、保护级别等出发,选取了白鹭(Egrettagarzetta)、鸳鸯(Aixgalericulata)和雉鸡(Phasianuscolchicus)3种鸟类作为焦点物种[24- 34](表1)。

表1 研究区焦点物种基本特性
2.2.3地类划分
综合考虑焦点物种生境特点、数据精度以及土地利用规划与管理工作实际,将研究区地类划分为水田、旱地、园地、乔木林地、灌木林地、草地、城乡建设用地、交通用地、滩涂沼泽、湖泊水库、河流、沟渠/运河和其他未利用地13类。
2.2.4生境斑块识别
依据表1中焦点物种特性,分别构建出与之对应的生境斑块约束条件模型[27](表2)。通过ArcGIS 10.0实现生境斑块识别。

表2 焦点物种生境斑块约束条件
* 鸳鸯在苏锡常地区为冬候鸟,不应考虑其繁殖需求,所以不设筑巢地地类约束条件
2.2.5生境网络构建
物种迁徙模拟需要大量物种分布和生境结构数据以及对影响物种迁移因素(生境质量、水质情况、人为干扰强度等[35])的考虑。模拟时,可通过采用考察不同影响因素(猎物密度、高程、坡度、坡向、承载能力、道路密度等)的方法实现[36- 38];或者通过采用考察土地利用类型适宜性的方法实现[14,39- 40]。本文选择后者与最小成本路径法相结合的方法。
对鸟类土地利用类型适宜性认知上的差异是造成迁移阻力赋值差异[32,40- 41]的主要原因。参照已有研究结果[23,32],设定文中阻力值(表3)。白鹭和鸳鸯飞行能力强受地类影响小,阻力值在1—100;雉鸡以地面行走为主受地类影响较大,阻力值在1—500。借助ArcGIS 10.0生成潜在迁移廊道。
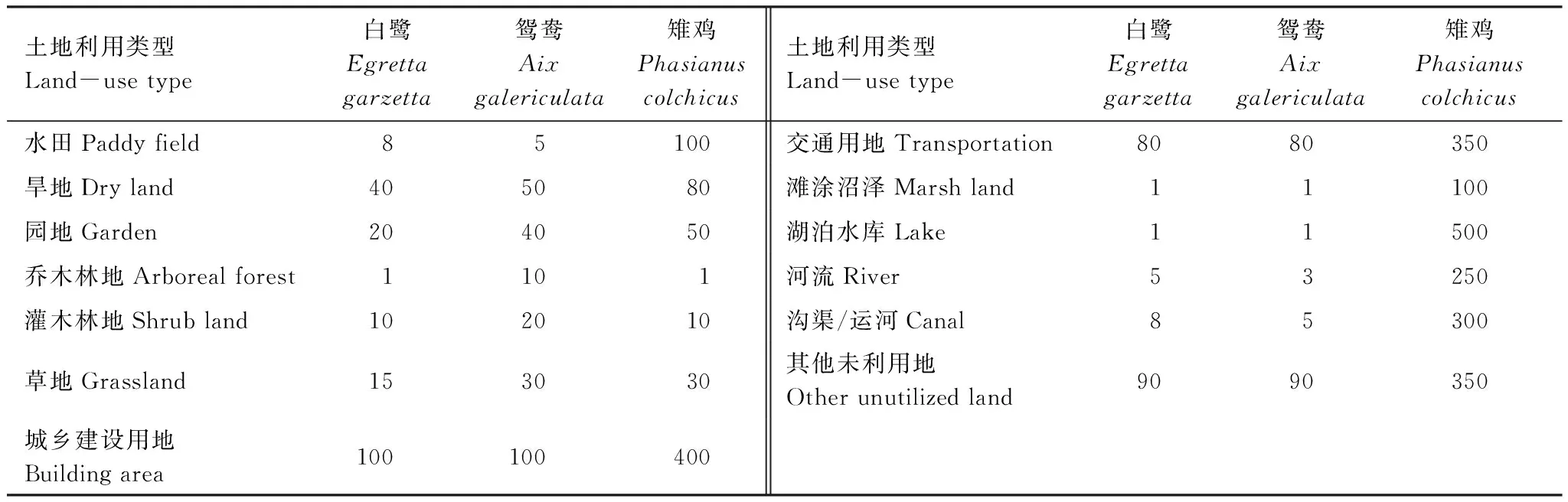
表3 不同土地利用类型对焦点物种迁移的阻力值
2.2.6生境网络复合
空间效能(成长性、稳定性)是衡量网络复合可行性的重要指标。效能越高,对应斑块拥有的生态功能越强,网络连接形态越有效[11]。生态系统服务强调生态系统结构、过程和功能具有的直接或间接生命支持产品和服务。价值高低反映了生态系统服务功能的强弱[42],适于衡量网络复合可行性。可以采用评价生态系统服务价值的方法进行廊道间的复合。参照相关研究结果[43- 44],对研究区土地利用类型生态系统服务价值进行修正(表4)。
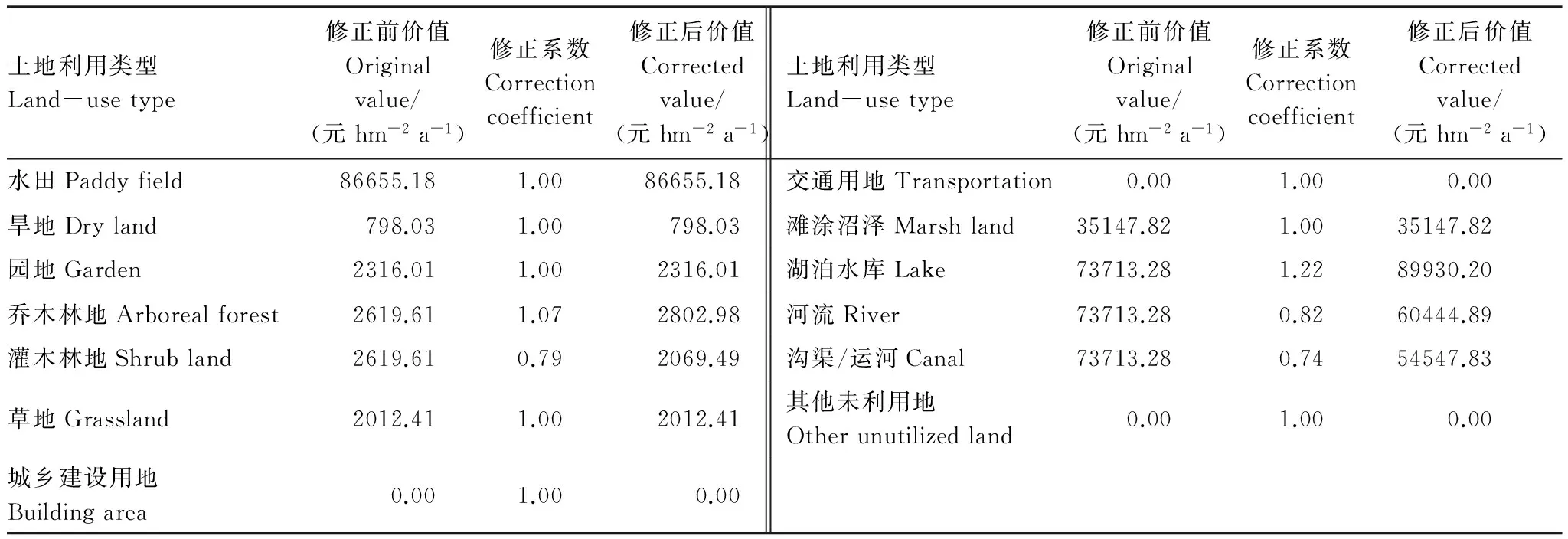
表4 研究区不同土地利用类型的生态系统服务价值
步骤为:最小成本路径法识别的廊道没有宽度。依据已有研究结果[45]设定鸟类迁移廊道宽度为200 m。对廊道复合情境作以下设定:(1)当两个生境节点(生境斑块)间存在多条同一物种廊道时(图1a),保留单位面积生态系统服务价值最高者(图1a′);(2)当存在多条不同物种廊道时(图1b),以保证物种迁移为前提,将生态系统服务价值转换为阻力值(表5),采用最小成本路径法模拟新廊道,保留单位面积生态系统服务价值最高者(图1b′)。叠加保留的廊道和生境斑块,即构成复合生境网络。
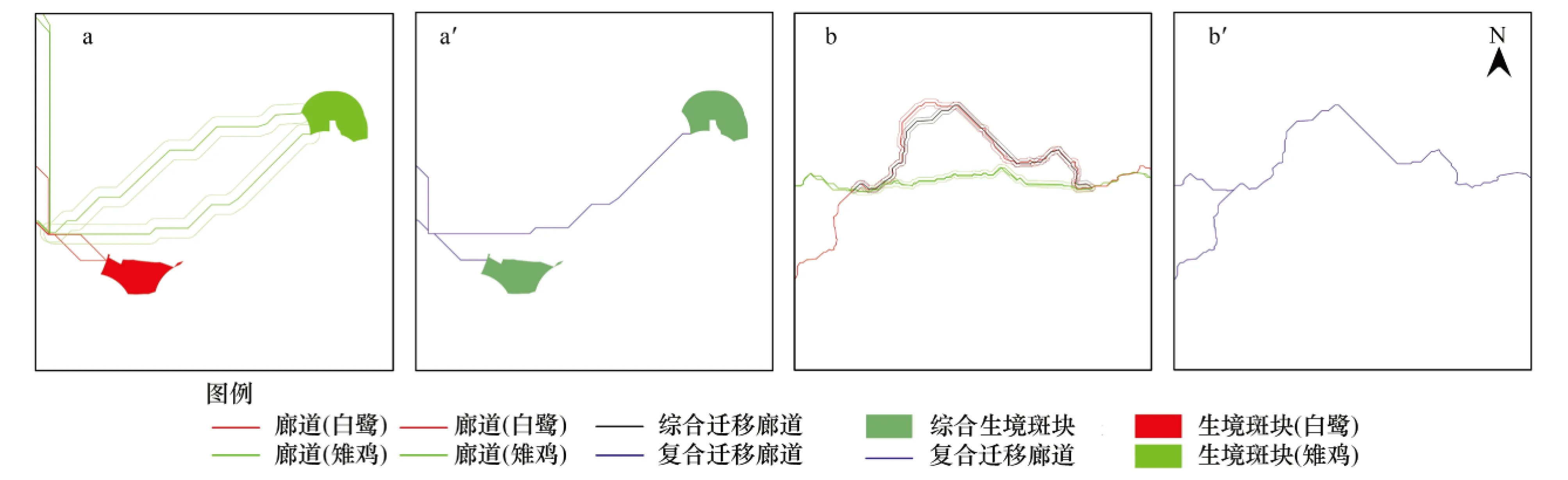
图1 生境网络复合情景设定Fig.1 Scenarios of habitat network recombination
2.2.7复合生境网络优化
观测数据是生境斑块识别及生境网络优化的重要依据。采用观测数据优化生境网络的方法步骤为:整理研究区焦点物种观测点及观测数量,绘制观测点分布图。与复合生境网络重叠。考虑到观测误差,将生境斑块及廊道边界外缘50 m范围以内的观测点按位于复合生境网络上处理,其余按位于复合生境网络外处理。综合观测次数、物种出现频率、观测点面积等因素,筛选位于复合生境网络外的观测点。以筛选出的观测点为生境斑块和迁移廊道增补依据,采用2.2.5和2.2.6节方法,优化复合生境网络。通过ArcGIS 10.0实现。
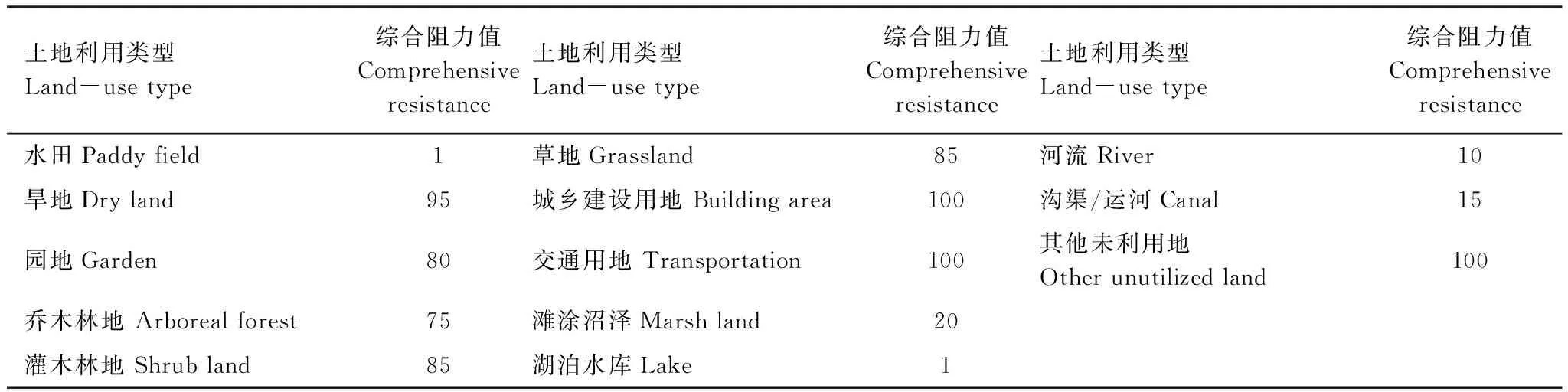
表5 不同土地利用类型对物种迁移的综合阻力值
3 结果
3.1生境斑块和生境网络
图2分别为2010年苏锡常地区白鹭、鸳鸯、雉鸡的生境网络。将全部太湖作为白鹭和鸳鸯的生境与实际情况不符。依据水中活动能力分别剔除边界纵深10 km(白鹭)和12 km(鸳鸯)以外部分;雉鸡为陆禽,不具备从陆地迁移至湖中岛屿的能力,不存在湖中岛屿与陆地之间的迁移廊道。
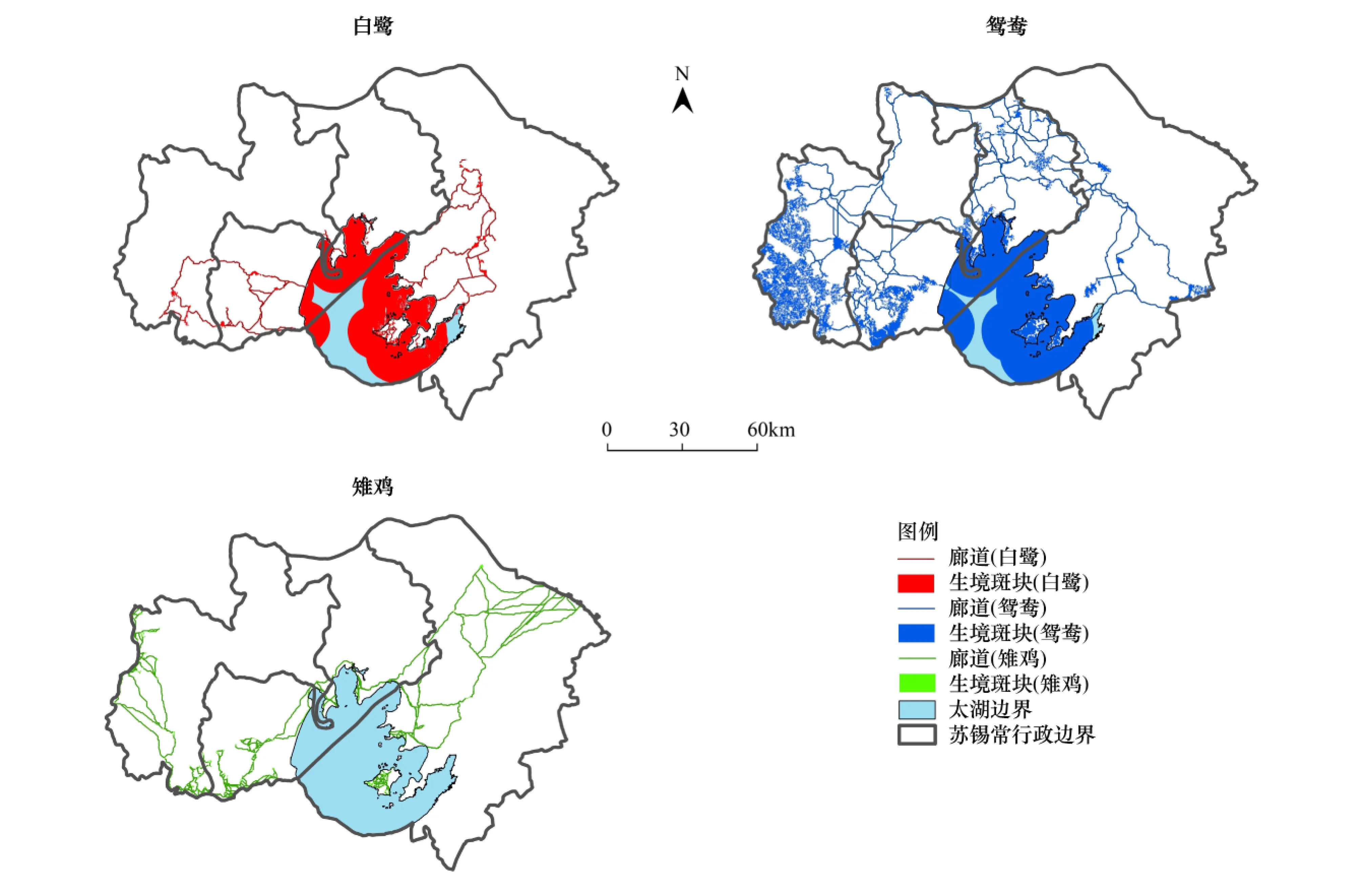
图2 研究区焦点物种的生境网络Fig.2 The habitat networks of the focal species in the study area
表6为焦点物种生境斑块和廊道情况。白鹭生境斑块以环太湖为主;鸳鸯生境斑块除太湖外,多集中在溧阳—宜兴,江阴也有分布;雉鸡生境斑块主要在溧阳—宜兴和苏州。鸳鸯生境斑块平均面积最大、白鹭次之、雉鸡则小而多;鸳鸯迁移廊道平均长度最长、雉鸡次之、白鹭最短。
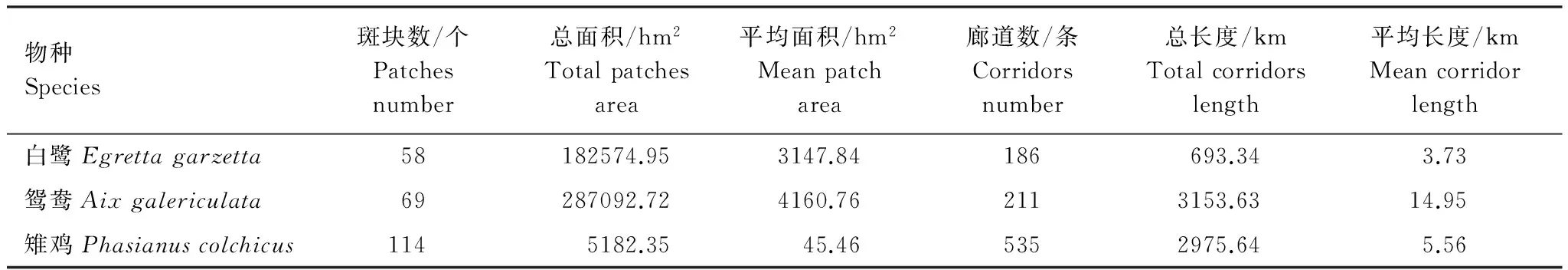
表6 焦点物种生境斑块和廊道情况
3.2叠加生境网络的复合
图3分别为3种鸟类生境网络的叠加和复合结果。生境斑块叠加后总面积为293871.93 hm2,为三者生境面积总和的62%。太湖是白鹭和鸳鸯最大的共有生境斑块,分别占各自生境总面积的95%和79%以上(表7)。
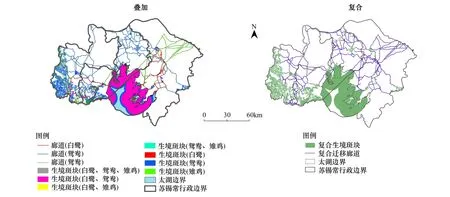
图3 焦点物种生境网络的叠加和复合结果Fig.3 The overlapped habitat network of the focal species and its recombined network

物种Species斑块数/个Patchesnumber斑块总面积/hm2Totalpatchesarea斑块平均面积/hm2MeanpatchareaA381298.4934.17B72177943.482471.44C331303.2939.49D793029.8138.35
A: 白鹭、鸳鸯、雉鸡;B: 白鹭、鸳鸯;C: 白鹭、雉鸡;D: 鸳鸯、雉鸡
复合后,廊道总面积、总长度及生态系统服务价值分别减少了16%、68%和10%,实现了集合覆盖问题。构成地类中,减少最多的地类依次为旱地(65%)、草地(50%)和灌木林地(46%),单位生态系统服务价值较高的地类如滩涂沼泽、河流、水田、湖泊水库和沟渠/运河得到较大程度保留(表8)。
3.3复合生境网络的优化
图4为2003—2014年间观测点分布情况。白鹭、鸳鸯、雉鸡的观测点分别有77、9和47处。其中,5处同时观测到3个物种、3处同时观测到白鹭和鸳鸯、33处同时观测到白鹭和雉鸡、没有同时观测到鸳鸯和雉鸡的记录。研究区实际观测点为87处。与复合生境网络空间叠置,53处与生境斑块重叠、22处位于复合廊道上、12处位于复合生境网络之外(表9)。对上述12处观测点地类、面积、观测时间及观测物种数量进行分析,确定了点2、4、5、6、8、9、10、11、12为增补生境斑块。
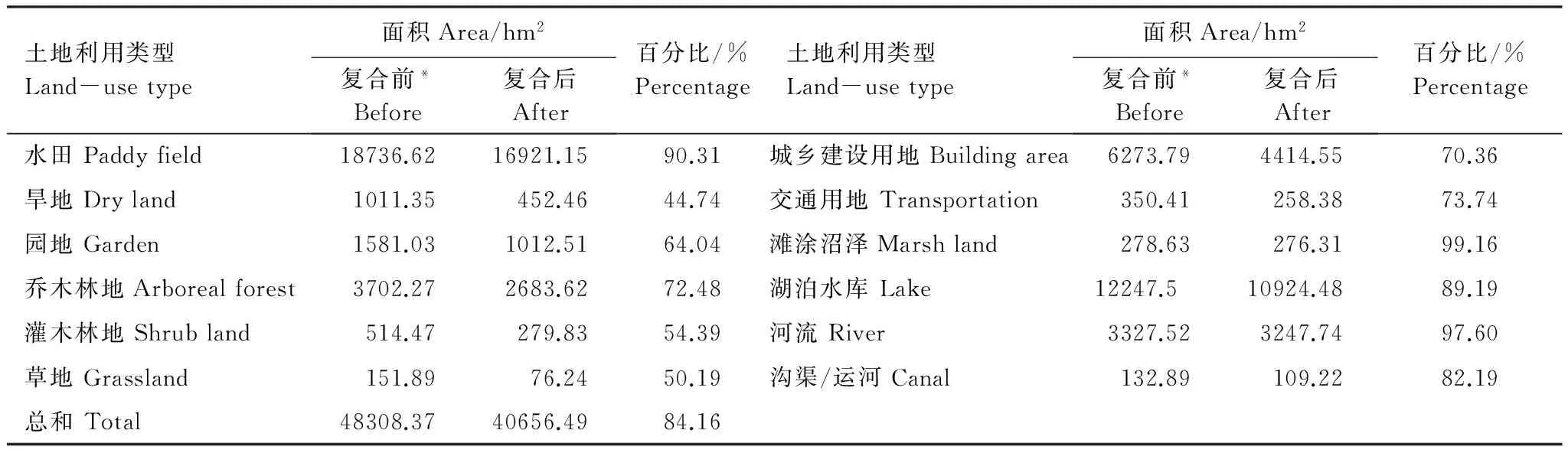
表8 复合前后廊道地类构成数量变化情况
*表内复合前廊道不包括被合并到复合生境斑块中的廊道
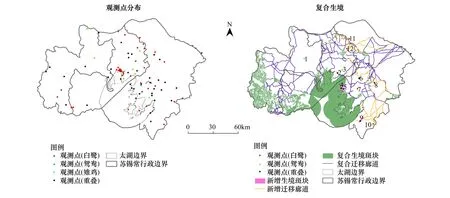
图4 研究区焦点物种观测点分布情况及复合生境网络优化结果Fig.4 The observation spots distribution and the recombined habitat network optimization of the focal species in the study area
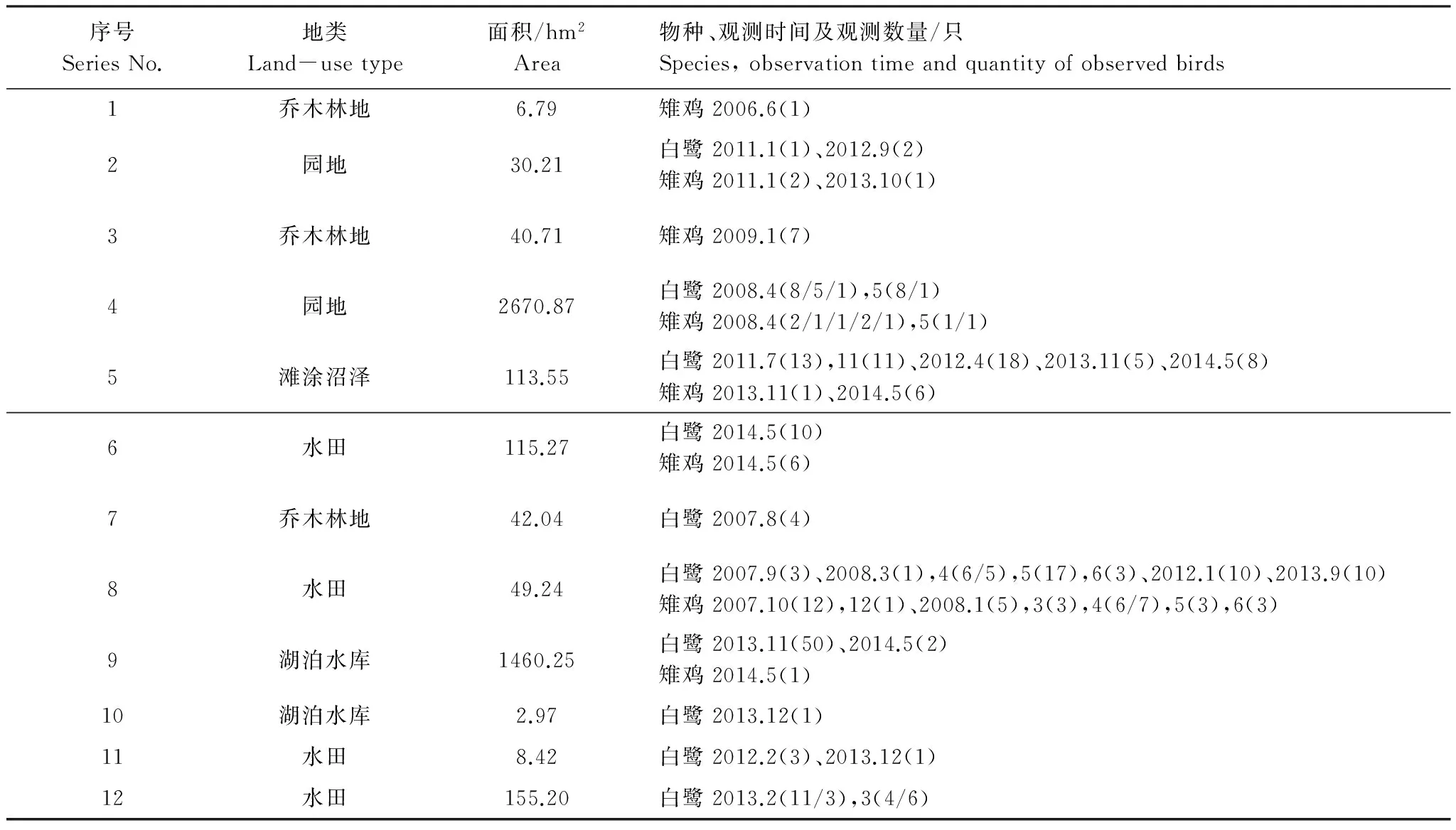
序号SeriesNo.地类Land-usetype面积/hm2Area物种、观测时间及观测数量/只Species,observationtimeandquantityofobservedbirds1乔木林地6.79雉鸡2006.6(1)2园地30.21白鹭2011.1(1)、2012.9(2)雉鸡2011.1(2)、2013.10(1)3乔木林地40.71雉鸡2009.1(7)4园地2670.87白鹭2008.4(8/5/1),5(8/1)雉鸡2008.4(2/1/1/2/1),5(1/1)5滩涂沼泽113.55白鹭2011.7(13),11(11)、2012.4(18)、2013.11(5)、2014.5(8)雉鸡2013.11(1)、2014.5(6)6水田115.27白鹭2014.5(10)雉鸡2014.5(6)7乔木林地42.04白鹭2007.8(4)8水田49.24白鹭2007.9(3)、2008.3(1),4(6/5),5(17),6(3)、2012.1(10)、2013.9(10)雉鸡2007.10(12),12(1)、2008.1(5),3(3),4(6/7),5(3),6(3)9湖泊水库1460.25白鹭2013.11(50)、2014.5(2)雉鸡2014.5(1)10湖泊水库2.97白鹭2013.12(1)11水田8.42白鹭2012.2(3)、2013.12(1)12水田155.20白鹭2013.2(11/3),3(4/6)
资料来源:中国观鸟记录中心http://www.birdtalker.net/
图4为复合生境网络优化结果。通过优化,生境网络覆盖了2010年及以后全部的84处观测点;新增廊道占廊道总长度、总面积及生态系统服务价值总量的比重分别为19%、21%和27%;按地类分,新增生态系统服务价值主要来自水田、湖泊水库、河流、乔木林地及沟渠/运河等(表10)。

表10 不同地类在新增廊道生态系统服务价值中的贡献率
复合前后廊道地类构成中均无其他未利用地
4 结论与讨论
本文以苏锡常地区为研究区域,选择了3种不同习性的鸟类为焦点物种,采用生境斑块约束条件模型和最小成本路径相结合的方法构建出叠加生境网络;从集合覆盖问题视角出发,通过比选出单位面积生态系统服务价值最高值廊道的方法实现了生境网络复合,采用与观测数据检测结合的方法实现了复合生境网络优化。结果表明:叠加生境网络经过复合,廊道总面积、总长度及生态系统服务价值在分别减少16%、68%和10%的情况下,保持了斑块间的连接度;在兼顾经济效益和生态效益最大化的同时,覆盖了占总数86%以上的75处观测点;复合生境网络经过优化,覆盖了2010年及以后所有84处观测点,廊道总面积、总长度及生态系统服务价值分别增加了19%、21%和27%,证实了设计的生境网络复合优化方法具有可行性。
优化前位于复合生境网络外的12处观测点被遗漏的可能原因是:(1)2010年之前符合生境斑块约束条件,但因斑块破碎(点1)或其它约束条件改变(点3、7)未被识别出;(2)2010年不符合约束条件,后因达到临界面积(点11)或其它约束条件改变(点6、12;点10、9、5)而符合所致;(3)符合约束条件,但未被设定为生境斑块的部分地类,如园地(点2、4);(4)不符合约束条件,被遗漏的生境斑块(点8)。可能是对鸟类亲善的传统或保护意识增强缩短了惊飞距离、或食物丰富等因素致使鸟类多次出现;(5)未知或偶然性原因(点1、3、7、10)。
将生境、生境网络及生物多样性保护纳入土地利用规划管理中,具有一定指导意义,益于研究结果实施。但物种生境地类与我国土地利用分类体系不完全衔接,物种生境斑块的识别存在误差,同时采用最小成本路径法模拟廊道时以土地利用类型设定阻力值,也存在一定主观性,需要继续改进和完善。
以鸟类为焦点物种探讨生境网络复合优化的研究思路和量化分析方法,具有一定普适性。但网络复合优化因物种差异而会出现很大差别,如小型兽类需要高精度遥感影像[39]和较完整的道路数据[9]、两栖类动物需要详细的水体水质资料等,所以该方法仍有待检验。
生境网络是复杂的开放网络系统。区域内生境网络复合优化还应考虑与周边区域或更大范围生境网络的合理衔接。从大尺度或多尺度转换视角深入探讨[1]——对于候鸟等长距离迁徙物种而言——更具意义。
以观测数据为依据优化生境网络需要建立长期连续的数据库,当前发展水平的大尺度研究还很困难。本文鸟类分布数据在连续性和完整性上都存在不足,可以作为验证依据,但不能作为识别依据。即便有较完整的数据库,还应考虑土地覆盖现状、道路密度、自然区密度等其它因素的影响[9]。随着全球对地观测技术水平的发展,上述问题将逐一得到解决[46],应得到密切关注。
网络复合是网络构建与优化研究的组成部分。现有研究,或者从微观生境网络出发,或者从宏观生态网络出发,对于中观及宏微观之间的衔接现有研究还不多。网络复合研究在此提供了新视野和理论基础。
引入生态系统服务价值是对“格局-过程-服务-福祉”机理系统研究的一种尝试,通过分析生态系统服务变化实现定量考察生态过程,为探索景观格局及网络构建与优化提供了新思路。
致谢:南京农业大学动物科技学院连新明副教授帮助写作,特此致谢。
[1]尹海伟, 孔繁花, 祈毅, 王红扬, 周艳妮, 秦正茂. 湖南省城市群生态网络构建与优化. 生态学报, 2011, 31(10): 2863- 2874.
[2]王海珍, 张利权. 基于GIS、景观格局和网络分析法的厦门本岛生态网络规划. 植物生态学报, 2005, 29(1): 144- 152.
[3]Jongman R H G. Nature conservation planning in Europe: developing ecological networks. Landscape and Urban Planning, 1995, 32(3): 169- 183.
[4]Jongman R H G. Landscape planning for biological diversity in Europe. Landscape Research, 2002, 27(2): 187- 195.
[5]Saura S, Pascual-Hortal L. A new habitat availability index to integrate connectivity in landscape conservation planning: comparison with existing indices and application to a case study. Landscape and Urban Planning, 2007, 83(2/3): 91- 103.
[6]Lookingbill T R, Gardner R H, Ferrari J R, Keller C E. Combining a dispersal model with network theory to assess habitat connectivity. Ecological Applications, 2010, 20(2): 427- 441.
[7]Bazelet C S, Samways M J. Relative importance of management vs. design for implementation of large-scale ecological networks. Landscape ecology, 2011, 26(3): 341- 353.
[9]Huber P R, Greco S E, Thorne J H. Spatial scale effects on conservation network design: trade-offs and omissions in regional versus local scale planning. Landscape Ecology, 2010, 25(5): 683- 695.
[10]Ayram C A C, Mendoza M E, Salicrup P S D R, Granados E L. Identifying potential conservation areas in the Cuitzeo Lake basin, Mexico by multitemporal analysis of landscape connectivity. Journal for Nature Conservation, 2014, 22(5): 424- 435.
[11]陈剑阳, 尹海伟, 孔繁花, 幺贵鹏. 环太湖复合型生态网络构建. 生态学报, 2015, 35(9): 3113- 3123.
[12]张小飞, 王仰麟, 李正国. 基于景观功能网络概念的景观格局优化——以台湾地区乌溪流域典型区为例. 生态学报, 2005, 25(7): 1707- 1713.
[13]Zetterberg A, Mörtberg U M, Balfors B. Making graph theory operational for landscape ecological assessments, planning, and design. Landscape and Urban Planning, 2010, 95(4): 181- 191.
[14]Gurrutxaga M, Rubio L, Saura S. Key connectors in protected forest area networks and the impact of highways: a transnational case study from the Cantabrian Range to the Western Alps (SW Europe). Landscape and Urban Planning, 2011, 101(4): 310- 320.
[15]许文雯, 孙翔, 朱晓东, 宗跃光, 李杨帆. 基于生态网络分析的南京主城区重要生态斑块识别. 生态学报, 2012, 32(4): 1264- 1272.
[16]王宜成. 最优化设计连续的自然保护区. 生态学报, 2011, 31(17): 5033- 5041.
[17]李晖, 易娜, 姚文璟, 王思琪, 李志英, 杨树华. 基于景观安全格局的香格里拉县生态用地规划. 生态学报, 2011, 31(20): 5928- 5936.
[18]Lambeck R J. Focal species: a multi-species umbrella for nature conservation. Conservation Biology, 1997, 11(4): 849- 856.
[19]Zacharias M A, Roff J C. Use of focal species in marine conservation and management: a review and critique. Aquatic Conservation: Marine and Freshwater Ecosystems, 2001, 11(1): 59- 76.
[20]Watts K, Eycott A E, Handley P, Ray D, Humphrey J W, Quine C P. Targeting and evaluating biodiversity conservation action within fragmented landscapes: an approach based on generic focal species and least-cost networks. Landscape Ecology, 2010, 25(9): 1305- 1318.
[21]俞孔坚, 李迪华, 段铁武. 生物多样性保护的景观规划途径. 生物多样性, 1998, 6(3): 45- 52.
[22]俞孔坚, 王思思, 李迪华, 李春波. 北京市生态安全格局及城市增长预景. 生态学报, 2009, 29(3): 1189- 1204.
[23]胡望舒, 王思思, 李迪华. 基于焦点物种的北京市生物保护安全格局规划. 生态学报, 2010, 30(16): 4266- 4276.
[24]阮禄章, 张迎梅, 赵东芹, 董元华, Mauro F. 白鹭作为无锡太湖地区环境污染指示生物的研究. 应用生态学报, 2003, 14(2): 263- 268.
[25]张迎梅, 阮禄章, 董元华, 龚钟明, 李运东, 王辉, Mauro F. 无锡太湖地区夜鹭及白鹭繁殖生物学研究. 动物学研究, 2000, 21(4): 275- 278.
[26]Palomino D, Carrascal L M. Threshold distances to nearby cities and roads influence the bird community of a mosaic landscape. Biological Conservation, 2007, 140(1/2): 100- 109.
[27]吴未, 张敏, 许丽萍, 欧名豪. 土地利用变化对生境网络的影响分析——以苏锡常地区白鹭为例. 生态学报, 2015, 35(14), doi: 10.5846/stxb201312112930.
[28]Hinsley S A, Bellamy P E, Newton I, Sparks T H. Habitat and landscape factors influencing the presence of individual breeding bird species in woodland fragment. Journal of Avian Biology, 1995, 26(2): 94- 104.
[29]赵正阶. 中国鸟类志. 吉林: 吉林科学技术出版社, 2001.
[30]郑光美, 王岐山. 中国濒危动物红皮书——鸟类. 北京: 科学出版社, 1998.
[31]孔凡前, 何万存, 张毅, 邹琴, 许仕. 鸳鸯庐山停歇地生态因子的研究. 江西林业科技, 2013, (4): 26- 28.
[32]刘海龙. 基于过程视角的城市地区生物保护规划——以浙江台州为例. 生态学杂志, 2010, 29(1): 8- 15.
[33]Li H Q, Lian Z M, Chen C G. Winter foraging habitat selection of brown-eared pheasant (Crossoptilonmantchuricum) and the common pheasant (Phasianuscolchicus) in Huanglong Mountains, Shaanxi Province. Acta Ecologica Sinica, 2009, 29(6): 335- 340.
[34]The Cornell Lab of Ornithology. http://www.allaboutbirds.org/guide/Ring-necked_Pheasant/lifehistory.
[35]朱曦, 邹小平. 中国鹭类. 北京: 中国林业出版社, 2001.
[36]曲艺, 栾晓峰. 基于最小费用距离模型的东北虎核心栖息地确定与空缺分析. 生态学杂志, 2010, 29(9): 1866- 1874.
[37]Peterman W E, Feist S M, Semlitsch R D, Eggert L S. Conservation and management of peripheral populations: spatial and temporal influences on the genetic structure of wood frog (Ranasylvatica) populations. Biological Conservation, 2013, 158: 351- 358.
[38]Peterman W E, Feist S M, Semlitsch R D, Eggert L S. Conservation and management of peripheral populations: spatial and temporal influences on the genetic structure of wood frog (Ranasylvatica) populations. Biological Conservation, 2013, 158: 351- 358.
[39]郭纪光, 蔡永立, 罗坤, 左俊杰, 刘志国, 倪静雪. 基于目标种保护的生态廊道构建——以崇明岛为例. 生态学杂志, 2009, 28(8): 1668- 1672.
[40]傅强, 宋军, 毛锋, 吴永兴, 姚涵, 唐剑波. 青岛市湿地生态网络评价与构建. 生态学报, 2012, 32(12): 3670- 3680.
[41]王洪新, 路建国, 张利, 门明新, 霍习良. 基于生物多样性保护的曹妃甸新区城镇用地规划研究. 水土保持研究, 2013, 20(6): 298- 301.
[42]Costanza R, d′Arge R, de Groot R, Farber S, Grasso M, Hannon B, Limburg K, Naeem S, O′Neill R V, Paruelo J, Raskin R G, Sutton P, van den Belt M. The value of the world′s ecosystem services and natural capital. Nature, 1997, 387(6630): 253- 260.
[43]王振波, 方创琳, 王婧. 1991年以来长三角快速城市化地区生态经济系统协调度评价及其空间演化模式. 地理学报, 2011, 66(12): 1657- 1668.
[44]何浩, 潘耀忠, 朱文泉, 刘旭拢, 张晴, 朱秀芳. 中国陆地生态系统服务价值测量. 应用生态学报, 2005, 16(6): 1122- 1127.
[45]Stauffer F, Best L B. Habitat selection by birds of riparian communities: evaluating effects of habitat alterations. The Journal of Wildlife Management, 1980, 44(1): 1- 15.
[46]Rose R A, Byler D, Eastman J, Fleishman E, Geller G, Goetz S, Guild L, Hamilton H, Hansen M, Headley R, Hewson J, Horning N, Kaplin B A, Laporte N, Leidner A, Leimgruber P, Morisette J, Musinsky J, Pintea L, Prados A, Radeloff V C, Rowen M, Saatchi S, Schill S, Tabor K, Turner W, Vodacek A, Vogelmann J, Wegmann M, Wilkie D, Wilson C. Ten ways remote sensing can contribute to conservation. Conservation Biology, 2015, 29(2): 350- 359.
Recombination and optimization of bird habitat networks: a case study of the Little Egret (Egrettagarzetta), Mandarin Duck (Aixgalericulata) and Ring-necked Pheasant (Phasianuscolchicus) in Su-Xi-Chang area
WU Wei1, HU Yuting1, FAN Shiwei1, OU Minghao1,2,*
1CollegeofLandManagement,NanjingAgriculturalUniversity,Nanjing210095,China2NationalJointLocalEngineeringResearchCenterforRuralLandResourcesUseandConsolidation,NanjingAgriculturalUniversity,Nanjing210095,China
Habitat networks play an important role in species survival and biodiversity conservation. However, habitat networks vary among species; habitat networks generated by simply overlapping networks of a few species do not meet the needs of most species. It is necessary to develop theories and methods of network recombination so that different habitat networks can be combined into one complex network for empirical implementation in rapidly urbanizing areas. The purpose of this paper was to examine methods of recombining and optimizing habitat networks. The Su-Xi-Chang area of the Yangtze River Delta region was selected for study, and the Little Egret (Egrettagarzetta), Mandarin Duck (Aixgalericulata), and Ring-necked Pheasant (Phasianuscolchicus) were selected as focal species because of their different biological characteristics, habitats, and diets. The habitats of these species were identified using our previously developed method, the Conceptual Constraint Model of Species Habitat Patch (CCMSHP) from land-use data for 2010. Potential corridors were identified based on the identified habitats using the least-cost path method. The habitat networks of the three focal species were overlapped using ArcGIS. From the perspective of thesetcoveringproblem, these three habitat networks were combined by recombining network structural elements, i.e., patches with patches, patches with corridors, and corridors with corridors. Among these scenarios, the recombination of corridors with corridors was the most complex. A method for evaluating the ecosystem service value of corridors was developed, and the combined corridors and habitat network were identified with the model. Optimization of the habitat network was simulated by applying observation points as newly added habitats and corridors, using a dataset (2003—2014) obtained from the website ofChinaBirdReport. All data were calculated using ArcGIS version 10.0. After recombination, the total area, length, and ecosystem service value of all corridors were reduced by 16%, 68%, and 10%, respectively, relative to the original values; however, the connectivity of the network remained the same. The combined network covered 75 observation points (more than 86% of all points) and achieved the target of simultaneously maximizing the economic and ecological benefits. After model optimization, the habitat network covered all 84 observation points after 2010, and the total area, length, and ecosystem service value of all corridors increased by 19%, 21%, and 27%, respectively, relative to the original values. These results demonstrate that the combined quantitative analysis method developed here is reasonable and feasible. This method will be useful for theoretical research on habitat network recombination.
network recombination; network optimization; habitat network; methodology;Egrettagarzetta;Aixgalericulata;Phasianuscolchicus; Su-Xi-Chang area
国家自然科学基金资助项目(41571176);江苏高校哲学社会科学研究项目(2015SJD096);中央高校基本科研业务费人文社会科学研究基金配套项目(SKPT2015018)
2015- 01- 11; 网络出版日期:2015- 11- 16
Corresponding author.E-mail: mhou@njau.edu.cn
10.5846/stxb201501110080
吴未, 胡余挺, 范诗薇, 欧名豪.不同鸟类生境网络复合与优化——以苏锡常地区白鹭、鸳鸯、雉鸡为例.生态学报,2016,36(15):4832- 4842.
Wu W, Hu Y T, Fan S W, Ou M H.Recombination and optimization of bird habitat networks: a case study of the Little Egret (Egrettagarzetta), Mandarin Duck (Aixgalericulata) and Ring-necked Pheasant (Phasianuscolchicus) in Su-Xi-Chang area.Acta Ecologica Sinica,2016,36(15):4832- 4842.
