乡土植物生活型构成对川渝地区边坡植被水土保持效益的影响
2016-10-24潘声旺雷志华胡明成
潘声旺,袁 馨,雷志华,胡明成
1 成都大学城乡建设学院,成都 610106 2 解放军后勤工程学院国防建筑与环境工程系,重庆 401131 3 信阳职业技术学院检验技术学院,信阳 464000
乡土植物生活型构成对川渝地区边坡植被水土保持效益的影响
潘声旺1,*,袁馨2,雷志华3,胡明成1
1 成都大学城乡建设学院,成都610106 2 解放军后勤工程学院国防建筑与环境工程系,重庆401131 3 信阳职业技术学院检验技术学院,信阳464000
乡土植物灌木化建植是高速公路边坡防护的重要生态模式。为了探讨生态恢复过程中乡土植物的生活型构成对边坡植被水土保持效益的影响,揭示乡土植物生活型-物种多样性-生态系统功能间的偶合关系,借助3 个物种配置试验,于2009 年4 月构建了以草本、灌木或乔木为主体的草本型、灌木型、乔木型绿化配置及草-灌-乔混合型试验区。自建植次年(2010年)起,对试验区进行持续5a生态监测。结果表明:1)边坡植被的物种丰富度与乡土植物的生活型有关,呈现乔木型>灌木型>草-灌-乔混合型>草本型趋势;2)植被的物种多样性(Shannon-Wiener指数、Pielou指数)与乡土植物生活型构成及建植年限有关:建植后第1、第2年,多样性水平呈草本型>草-灌-乔混合型>灌木型>乔木型变化趋势;自建植后第三起(2012—2014年),呈草-灌-乔混合型>草本型>灌木型>乔木型波动;3)植被的水土保持性能(径流系数、侵蚀模数)与群落的物种多样性密切相关:多样性水平越高,水土保持性能越强。可见,乡土植物的生活型构成对提高边坡植被的物种多样性、改善生态性能至关重要。
乡土植物;生活型;物种多样性;水土保持性能
在恢复生态学理论的影响下,借助乡土植物的生态优势,在裸露边坡上重建边坡植被的生态固坡技术已成为改善路域景观、减少水土流失的重要途径[1- 3]。研究表明,草灌(乔)混合群落地上茎叶对雨滴的分层拦截和缓冲作用,地下根系对土壤的加筋作用和锚固作用,有利于减小坡面水土流失、增强坡体稳定性[4-5]。因灌(乔)木生长缓慢,需配合草本植物防护坡面,以控制早期土壤侵蚀;草本类在建植初期生长过旺,草、灌(乔)间的激烈竞争,常导致灌、乔类生长受抑制,甚至被淘汰[6]。如何建立结构合理的植物群落、缩短人工植被的演化进程,形成以本地乡土植物为优势种的稳定群落,已成为遏制生态固坡技术的瓶颈。
重建后的植被群落是否稳定、能否实现自然演替是衡量人工植被恢复成功与否的重要标志。虽然边坡植被的人工恢复技术在我国已有10余年研究历史,但多数研究尚限于乡土植物的选配对植被水土保持效益的影响等方面[1,7-8],缺乏对重建后植被群落特征的持续跟踪研究,在一定程度上也制约了我国植被恢复质量的提高。近年来,潘声旺[2,9]等就乡土植物的丰富度对边坡植被群落特征及水土保持性能的影响进行了初步研究,但在乡土植物生活型构成对边坡植被群落特征、水土保持性能等方面的影响还鲜见报道。事实上,乡土植物的生活型构成潜在地影响着边坡植被的群落结构和多样性水平[10- 12],进而影响其水土保持性能。据此,本研究以草本、灌木和小乔木为主体分别构建草本型、灌木型、乔木型或草-灌-乔混合型初始配置的边坡绿化带,持续观测试验区内植被的物种多样性及其水土流失状况,主要目的:1)探讨乡土植物的生活型构成对边坡植被水土保持性能的影响效应;2)分析乡土植物生活型-物种多样性-生态系统功能间耦合关系,阐明乡土植物生活型构成对边坡植被水土保持性能的影响机制,为生态防护提供理论指导。
1 材料与方法
1.1研究区概况
研究区域为川东、渝西接壤处的丘陵地带(105°38′—106°05′ E、28°56′—29°34′ N),年均气温18.6 ℃,年均降雨量1165.2 mm,属亚热带湿润季风气候。土壤多为黄壤、黄棕壤,由白垩纪夹关组砖红色长石、块状钙质岩屑砂岩的风化残积物发育而成,土层厚度多在10—70 cm,呈微酸性至酸性。区内丘陵广布、溪沟纵横,平均海拔250—350 m,丘谷高差50—100 m;植被以松(PinusLinn)、柏(Sabina)、蕨(Pteridium)类为主,包括杉科、松科、桦木科、壳斗科49 科150 余种,具有典型的亚热带常绿阔叶林特征,森林覆被率低(<7.5%),水土流失严重,地质灾害频发。
试验边坡位于成渝高速(G85)永川段路基边坡(K305+140-K305+920)处,路基全长780 m、路面高程305—307 m,坡高5—6 m(填方最大高程为14.7 m)、整理成二级边坡,坡比1∶1.5,坡向NE25°。边坡为黄壤区砂泥岩互层高边坡,表层覆土(0—20 cm)微酸性(pH 6.5—6.9),透水性好,保水、保肥性能差,初渗速率为14.2—42.4 mm/min,稳渗速率为0.8—5.7 mm/min,贮水力为6.814—12.441 t/km2。
1.2试验材料
以川渝地区3 类(草类、灌类、乔类)代表性绿化配置——芒(Miscanthussinensis)+狗牙根(Cynodondactylon)、慈竹(Neosinocalamusaffinis)+马棘(Indigoferapseudotinctoria)、柏木(Cupressusfunebris)+银合欢(Leucaenaleucocephala)为基础,分别与野牛草(Buchloedactyloides)、黑麦草(Loliumperenne)、黄荆(Vitexnegundo)、棉槐(Amorphafruticosa)、臭椿(Ailanthusaltissima)、刺槐(Robiniapseudoacacia)等路域优势种中任一种随机组合,建立3 个物种配置试验,构建以草本(Hi)、灌木(Si)、小乔木(Ai)为主体的草本型(HHXi)、灌木型(SSXi)、乔木型(AAXi)绿化配置,或由不同路域优势种构建草-灌-乔混合型(HSA)绿化配置(表1)。

表1 不同配置模式中绿化植物的组成
1.3试验设计
选择坡比、坡向及其它立地条件基本一致的边坡地段,整理成108 个矩形小区(长8 m、宽6 m),使小区的长边与公路边坡的等高线垂直、短边与等高线平行;四周用石棉瓦围埂(埂高25 cm)、塑料薄膜密封(埋入地下30 cm),小区上方及两侧开排水沟拦截小区外径流,下方修截流沟、集水池。
将待测配置模式随机分配于4 个小区。2009 年4 月,参照萌发试验结果,将催芽后的种子以10 g/m2播种密度撒播于相应小区、覆撒0.5—1 cm细砂后,无纺布覆盖。另设4 个对照区:2 个裸地观测区(CK0),定期除草以观测裸露边坡的产沙、产流特征;2 个次生裸地区(CK1)不除草以观测边坡的自然演替。
1.4测定项目
(1)群落结构2010 年8 月下旬,在各样地距路面边缘1—6 m处坡面上设置5×5 m样方,调查乔、灌类植物种类、株高、株数及盖度;草本植物在3 个品字分布的1×1 m样方内调查。2011—2014 年8 月,重复布样。

(3)产沙量将集水池内泥水搅拌30 min,取中层径流泥沙样品500 mL,105 ℃下烘干、称重,由含沙量推算每次降雨小区内土壤流失量;将1 年内各小区土壤侵蚀量相加、换算成t/km2,即为侵蚀模数。
降雨量、径流量和泥沙量观测时段为2010—2014 年汛期(5—9 月)。
1.5物种多样性

1.6数据处理
数据经Excel 13.0软件处理后,用SPSS 13.0统计分析;将同类配置模式的调查数据归类、排序,分别求其上四分位数(Q3)、中位数(Q2)、下四分位数(Q1)及四分位距(Interquartile range,IQR=Q3-Q1)。
2 结果与分析
2.1边坡植被的物种多样性
2.1.1物种丰富度
不同绿化配置试验区的物种数差异很大(n=24,P<0.05),乔木型(AAXi)配置区内的物种数(中位数,Q2)明显多于其他配置,呈AAXi>SSXi>HSA>HHXi趋势(P<0.05);HHXi、SSXi、AAXi等同类配置中,乔木类取代种(Ai)试验区内物种数(Q3)明显多于灌、草类取代种(Si、Hi)试验区(n=8,P<0.05)(图1)。如2014 年雨季结束时,HHAi、SSAi、AAAi配置区内的物种数(Q3)分别为17.3、20.4和21.5,HHSi、SSSi、AASi配置区内的物种数(Q2)分别为16.5、19和19.7,而HHHi、SSHi、AAHi配置区内的物种数(Q1)则分别为15.4、17.7和18.5。同龄群落间,以AAXi配置区内的物种数四分位距(IQR)最大、SSXi次之,HSA最小。

图1 2010—2014年间不同配置模式试验区内物种丰富度Fig.1 Richness of plant species of slope vegetation differed in indigenous species′ life form during years from 2010 to 2014高低线上连接点(Q3、Q2、Q1)与被标注的初始生活型配置所在群落的物种丰富度(均值)相对应
试验期间,观测区内共出现29 个迁入种,隶属10 科15 属。其中灌木4 科6 属11 种,占总种数37.93%;草本7 科10 属18 种,占总种数62.07%。菊科、禾本科、豆科、蔷薇科种类最多,占总种数58.4%,为川渝地区植被恢复过程中的先驱种[9]。灌木多为豆科、蔷薇科、桑科;草本多为菊科、禾本科、蔷薇科、豆科;种数较多的属为悬钩子属(Rubus)、蒲公英属(Taraxacums)。其中,艾蒿(Artemisiaargyi)、三叶鬼针草(Bidenspilosa)、野菊花(Dendranthemainsicum)、白茅(Imperatacylindrica)、酢浆草(Oxaliscorniculata)、飞蓬(Erigeronacer)、苦荬菜(Ixerispolycephala)、牡蒿(Artemisiajaponica)、地果(Ficustikoua)、构树(Broussonetiapapyrifera)、马桑(Broussonetiapapyrifera)等出现频率较高,为川渝地区植被恢复过程中主要建群种或共建种。
2.1.2Shannon-Wiener指数
图2为试验期间不同绿化配置所在群落的Shannon-Wiener指数。可以看出,群落内Shannon-Wiener指数与乡土植物的生活型构成及建植年限有关。建坪后的第1、第2年(2010、2011年),草本型(HHXi)配置的Shannon-Wiener指数(Q2)远高于其他类型,HSA次之,AAXi最低,表现为HHXi>HSA>SSXi>AAXi(P<0.05);自建坪后第3年开始,HSA内Shannon-Wiener指数(Q2)最高,HHXi次之,AAXi最低,表现为HSA>HHXi>SSXi>AAXi(P<0.05)。不同配置间,以乔木型(AAXi)配置区的Shannon-Wiener指数四分位距(IQR)最大、SSXi次之,HHXi最小,说明乔木型配置区的Shannon-Wiener指数变异幅度大,草本型(HHXi)配置区内Shannon-Wiener指数较小、群落较稳定。同类配置但取代种(Ai、Si、Hi)不同的试验区间,除HHXi自建坪后第2年起呈HHSi>HHAi>HHHi波动(P<0.05)外,其他群落均表现为乔木类取代种(Ai)试验区内Shannon-Wiener指数最大,草本类取代种(Hi)试验区内最小。

图2 不同初始绿化配置边坡植被所在群落内Shannon-Wiener指数的变化差异Fig.2 Variation of Shannon-Wiener index in community differed in indigenous species′ life form during years from 2010 to 2014高低线上连接点与被标注的生活型配置所在群落的Shannon-Wiener指数相对应
2.1.3Pielou指数
图3为试验期间不同绿化配置试验区的Pielou指数。不难看出,4 类绿化配置的Pielou指数及年际间波动与试验区内Shannon-Wiener指数变化趋势大致相同:建坪后的第1年、第2年,HHXi内Pielou指数最高,HSA次之,AAXi最低,呈HHXi>HSA>SSXi>AAXi波动(P<0.05);自第3年起,HSA内Pielou指数最高,HHXi次之,AAXi最低,呈HSA>HHXi>SSXi>AAXi趋势(P<0.05)。
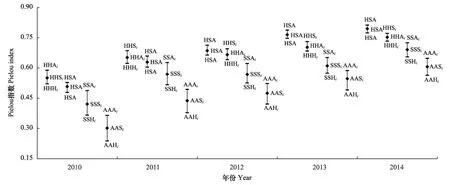
图3 不同初始绿化配置边坡植被所在群落内Pielou指数的变化差异Fig.3 Variation of Pielou index in community differed in indigenous species′ life form during years from 2010 to 2014高低线上连接点与被标注的生活型配置所在群落的Shannon-Wiener指数相对应
2.2护坡效益分析
2.2.1裸露边坡的产流、产沙特征
2010—2014 年汛期,试验区内自然降雨量及裸露边坡的产流、产沙特征如表2。可以看出,区内雨季集中(占年降雨量71.36%)、产流量大(年均降雨量830.28 mm;年均产流量150.54 mm),水土流失严重(年均产沙量:1177.24 g/m2),属地质灾害多发区。
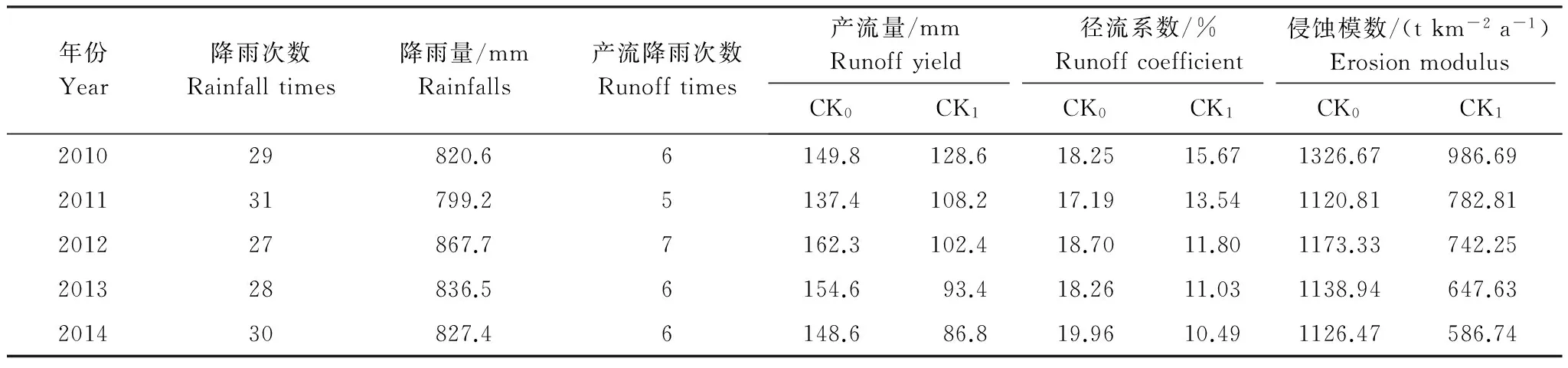
表2 2010—2014年汛期降雨情况及裸露边坡产流、产沙特征
* CK0为无植被裸地,CK1为自然演替的次生裸地
2.2.2试验区内的产流、产沙特征
径流系数、侵蚀模数是衡量植被涵养水源、固土护坡的主要指标,也是表征生态系统功能的重要尺度。图4显示,试验区内绿化配置不同,其径流系数、侵蚀模数也不一样(n=24,P<0.05),年际差异明显。
试验期间,边坡植被的产流、产沙特征与乡土植物的生活型构成及其建植年限有关。建坪后的第1、第2年汛期,草本型配置表现出较强的水土保持性能,小区内径流系数(7.81%)、侵蚀模数(92.68 t km-2a-1)明显小于其他试验区(n=24,P<0.05),不同生活型配置间的水土保持性能呈HHXi>HSA>SSXi>AAXi趋势,且差异明显(P<0.05);同类配置中,草本类取代种Hi试验区的水土保持性能远小于灌乔类取代种(Si、Ai)试验区(P<0.05)。但在第二年汛期,HHHi内的径流系数(6.32%)、侵蚀模数(92.39 t km-2a-1)却大于HHSi、HHAi区(分别为5.84%、5.51%和76.14、61.52 t km-2a-1) (P<0.05)。

图4 2010—2014年间不同绿化配置模式所在边坡的产流、产沙特征Fig.4 Characteristic of runoff and sediment yield in slope plots tested during rainy seasons ranging from 2010 to 2014高低线上连接点与被标注的生活型配置所在小区内的产流系数、侵蚀模数相对应
自建坪后第3年(2012 年)开始,HSA混合型配置却表现出较好的水土保持性能,小区内径流系数(分别为4.16%、3.76%和3.44%)、侵蚀模数(分别为51.85、35.52和28.46 t km-2a-1)明显低于其他观测区(n=24,P<0.05),不同生活型配置间的水土保持性能呈HSA>HHXi>SSXi>AAXi趋势,且差异明显(P<0.05);同类配置中,除HHXi呈现HHSi>HHAi>HHHi波动(P<0.05)外,其他群落中草本类取代种Hi所在小区的径流系数、侵蚀模数均明显小于灌乔类取代种(Si、Ai)所在小区(P<0.05)。
同一汛期内,AAXi配置区内径流系数、侵蚀模数的四分位距(IQR)最大,SSXi次之,HSA、HHXi较小;建植年限越久,趋势越明显。
2.3多样性水平与水土保持性能的相关性
试验期间,观测区内边坡植被的水土保持性能与所在群落的Shannon-Wiener指数密切相关:建坪后的第1、第2年(2010、2011 年)汛期,二者呈线性变量关系,R2>0.951(P<0.05);自建坪后第3年(2012—2014 年)汛期开始,二者呈对数函数关系,R2>0.982(P<0.05)(表3)。整个试验期内,边坡植被所在群落的Pielou指数与观测区内边坡植被的水土保持性能间也呈现类似于Shannon-Wiener指数的相关性。说明植物群落的物种多样性水平越高,生态系统的结构和功能越稳定、抗干扰(雨水冲蚀)能力越强[13]。
3 讨论
理论生态学中,生物多样性与系统稳定性的关系始终未能取得共识。究其原因,相关研究多集中在自然生态系统,而自然生态系统是与周围环境紧密结合在一起的生物地理群落,在相邻的同等环境条件下,不可能分布在结构上和种类成分上有明显差异的群落类型,很难找到不同生物多样性的群落结构进行比较研究;人工生态系统中,农业生态系统的经营对象多为短期作物,难于取得长期连续的数据;加之概念上的混乱及繁杂,也给系统稳定性研究带来一定困难[14]。探讨边坡植被人工恢复过程中乡土植物的生活型-物种多样性-水土保持性能间的耦合关系有望为多样性-稳定性研究开拓新的视野:降雨对不同多样性水平小区的作用相当于干扰、水土保持性能的变化则是系统的反应;有物质输出、有一致的干扰条件,便于分析系统内部结构变化、外部干扰及物质输出关系。
表3观测区内的径流系数(y)、侵蚀模数(y′)与所在群落Shannon-Wiener指数(x)、Pielou指数(x′)的关系
Table 3Relationship between Shannon-Wiener index, Pielou index and their capacity of soil and water conservation

年份Year径流系数Runoffcoefficient(y)/%侵蚀模数Erosionmodulus(y')/tkm-2a-1Shannon-Wienerindex(x)Pielouindex(x')Shannon-Wienerindex(x)Pielouindex(x')关系式RelationshipR2关系式RelationshipR2关系式RelationshipR2关系式RelationshipR22010y=-16.23x+16.9280.987y=-19.755x'+18.8540.947y'=-992.44x+648.520.987y'=-1275.2x'+796.390.9482011y=-22.061x+20.9490.951y=-23.239x'+21.3990.959y'=-1141.4x+843.480.969y'=-1199.1x'+864.880.9742012y=-12.598ln(x)+1.1510.982y=-13.786ln(x')-0.8420.990y'=-554.98ln(x)-98.200.997y'=-595.8ln(x')-179.980.9682013y=-12.645ln(x)+1.0010.999y=-13.343ln(x')-0.1320.941y'=-526.88ln(x)-79.620.999y'=-557.01ln(x')-127.310.9452014y=-14.903ln(x)+0.4620.994y=-13.123ln(x')+0.2780.993y'=-639.35ln(x)-96.9220.995y'=-562.52ln(x')-104.660.993
3.1生活型构成对边坡植被物种多样性的影响
对于特定区域的次生裸地(如边坡)而言,种子雨、种子库的区域异质相对较小,乡土植物所营造的微环境是影响其他物种侵入潜力和成功定居的主导因素[9,11]。绿化植物的种类不同,对光、热、水、气等环境资源的利用方式、利用强度及其根际沉积(rhizodeposition)、根际效应(rhizosphere effect)等生物学过程也存在一定差异,对局域生境的影响程度、范围也不一样[12]。与草本植物相比,乔、灌类拥有更丰富的空间层次结构,局域生境的异质性增强,为接纳其它物种提供了多样化生境。本研究显示,绿化植物的生活型构成对所在群落的物种丰富度影响很大,乔木型配置的物种数明显多于其他配置;同类配置中,乔木类取代种所在小区的物种数远多于灌草类,说明乔木类植物对局域环境的影响程度较大、生境的异质性较高。
乡土植物的生活型构成同样也影响群落中Shannon-Wiener指数、Pielou指数。建坪初期,草本类绿化种种子较小,植株密度大、分布均匀,其伴生植物也呈均匀分布,物种多样性水平较高;乔、灌类绿化种的层次结构较强,资源分配远不及草本类群落均匀,其伴生植物呈斑块状分布,多样性水平较低。本研究中,草本类试验区内多样性水平在建坪后前两年均高于乔灌类;自建坪后第2年开始,草-灌-乔配置内生态位互补(niche complementarity)效应日趋明显,多样性水平逐渐超越其他配置。该结论也印证了Pacala和Tilman的抽样向互补转型假说[15]:建群初期,多样性作用机制主要是抽样效应;随着时间推移,植物个体密度增加,种间资源利用性竞争成为群落动态的决定性因素,生态位互补逐渐成为主要的作用机制。
3.2生活型构成对边坡植被水土保持性能的影响
边坡植被的林冠层、灌草层、枯枝落叶层均有保水固土和维持坡面稳定的作用,且乔木、灌木和草本植物在整个群落中均有不可替代的作用。但植被类型不同,各层次的蓄水保土功能也不同,草本植物生长快,对防止早期的土壤侵蚀、减少水土流失效果好;豆科灌木植物具有固氮作用,改良土壤效果好;乡土灌木适应性强,能促进群落顺向演替;乔木可固土护坡、维持边坡的稳固,对边坡生态恢复的作用明显[5,16]。本研究中,草本型配置在建坪后第1、第2年的水土保持性能最强;随着时间推移,乔、灌木绿化种在群落中的优势度逐渐增强,草-灌-乔配置内层次结构日益明显,茎叶对雨滴的分层拦截和缓冲作用、根系对土壤的加筋作用和锚固作用更有利于减小坡面水土流失、增强坡体的稳定性。可见,植被的水土保持性能是林冠层、灌草层、枯枝落叶层和植物根系等结构层次,通过截持降水、拦蓄径流、保护地表层以及固土改土以提高土壤抗蚀抗冲性能来综合体现的。
3.3生活型-物种多样性-生态系统功能间耦合关系
生态护坡过程中,因资源利用、环境行为差异,不同生活型的绿化种对环境的影响程度及其生态效应均存在一定差异,群落中物种组成、数量和空间分布也不一样,进而影响群落中物种多样性[9,17]。物种多样性可通过物种互补-互促的确定性过程、功能优势种在生态系统中概率增大的随机过程,改变植物群落结构、生态系统功能[15]。物种丰富度增大,包含的生活型和垂直分层增多,叶面积指数增大、覆盖度随之提高[11];地上部分的层次结构如密度、覆盖度增大,可减缓降雨的侵蚀作用[9,13];根系空间结构变化,将直接影响根系的加筋、锚固效应及其抗蚀性能[12]。本研究显示,随着植物多样性增加,边坡植物的群落结构向有利于减小地表径流和土壤侵蚀方向发展。该结论与张全国等的理论预测相吻合[18]。
重建后的植被群落是否稳定、能否实现自然演替是衡量人工植被恢复成功与否的重要标志,但群落中优势种更新、群落演替是一个漫长的生态过程。本研究对观测区内物种多样性、水土保持性能的监测只有5a,时间尺度较短,不足以揭示生物多样性与系统稳定性的关系,相关内容尚待进一步研究。
4 结论
持续5 年的生态观测表明,建植条件、绿化植物物种数相同情况下,乡土植物的生活型构成与边坡植被的群落特征密切相关,并通过物种组成、多样性水平等特征影响植被的水土保持性能。主要表现在:
(1)边坡植被的生活型构成与所在群落的物种丰富度密切相关,呈现AAXi>SSXi>HSA>HHXi趋势;同类配置中,乔木类(Ai)取代种所在群落的物种数最高,灌木类(Si)次之,草本类(Hi)最低。
(2)生活型构成决定着边坡植被的Shannon-Wiener指数、Pielou指数:建坪后第1、第2年,不同配置试验区内Shannon-Wiener指数、Pielou指数呈现HHXi>HSA>SSXi>AAXi;自建坪后第3年开始,呈现HSA>HHXi>SSXi>AAXi。同类配置中,除HHXi自建坪后第二年起呈HHSi>HHAi>HHHi波动外,其他群落中,Ai取代种观测区内多样性水平最高,Si取代种次之,Hi取代种最小。
(3)生活型构成决定着植被的水土保持性能。建坪后第1、第2年汛期,不同配置试验区内水土保持性能呈AAXi>SSXi>HSA>HHXi;自建坪后第3年开始,表现为AAXi>SSXi>HHXi>HSA。
(4)边坡植被的水土保持性能与所在群落的物种多样性密切相关。建坪后第1、第2年,二者呈线性变量关系;自建坪后第3年开始,二者呈对数函数关系。多样性水平越高,群落的抗雨水冲蚀能力越强。
可见,在环境条件、建植措施一致的前提下,合理搭配绿化植物的生活型对提高边坡植被的物种多样性、改善水土保持性能至关重要。
[1]肖蓉, 高照良, 宋晓强, 张兴昌, 杨永峰. 高速公路边坡植被特征分析及护坡模式优化研究. 水土保持学报, 2009, 23(2): 90- 94.
[2]潘声旺, 杨秀云, 何茂萍, 杨丽娟. 几种典型道路边坡植被配置模式的护坡效益研究. 四川农业大学学报, 2013, 31(2): 151- 156.
[3]Lee S G, Hencher S R. Recent extreme rainfall-induced landslides and government countermeasures in Korea // Lollino G, Giordan D, Crosta G B, Corominas J, Azzam R, Wasowski J, Sciarra N. Engineering Geology for Society and Territory-Volume 2. Switzerland: Springer International Publishing, 2015: 1393- 1395.
[4]郑煜基, 卓慕宁, 李定强, 李生斌, 郑晓霞. 草灌混播在边坡绿化防护中的应用. 生态环境, 2007, 16(1): 149- 151.
[5]李妮, 陈其兵, 谭昌明. 高速公路边坡乡土灌木建植模式水土保持效益. 水土保持学报, 2012, 26(1): 67- 71.
[6]陶岩, 江源, 顾卫, 田育红. 草灌植被护坡群落中草本对灌木生长的竞争关系研究. 东北师范大学学报: 自然科学版, 2011, 43(2): 140- 144.
[7]刘窑军, 王天巍, 李朝霞, 蔡崇法. 不同植被防护措施对三峡库区土质道路边坡侵蚀的影响. 应用生态学报, 2012, 23(4): 896- 902.
[8]柴宗政, 王得祥, 张丽楠, 张洋, 黄青平, 吴昊. 秦岭西段油松天然次生林种群竞争关系研究. 西北农林科技大学学报: 自然科学版, 2012, 40(12): 57- 63.
[9]潘声旺, 何茂萍, 杨丽娟, 方文. 先锋植物丰富度对公路边坡植物群落生长发育及护坡效益的影响. 林业科学, 2013, 49(11): 24- 31.
[10]Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science, 2001, 294(5543): 804- 808.
[11]Ville V, Muchiru A, Reid R S, Ogutu J O. How pastoralism changes savanna vegetation: impact of old pastoral settlements on plant diversity and abundance in south-western Kenya. Biodiversity and Conservation, 2014, 23(13): 3219- 3240.
[12]Mlambo M C. Not all traits are ‘functional’: Insights from taxonomy and biodiversity-ecosystem functioning research. Biodiversity and Conservation, 2014, 23(3): 781- 790.
[13]王震洪, 段昌群, 侯永平, 杨建松. 植物多样性与生态系统土壤保持功能关系及其生态学意义. 植物生态学报, 2006, 30(3): 392- 403.
[14]冯耀宗. 物种多样性与人工生态系统稳定性探讨. 应用生态学报, 2003, 14(6): 853- 857.
[15]Pacala S, Tilman D. The transition from sampling to complementarity // Kinzig A P, Pacala S W, Tilman D, eds. The Functional Consequences of Biodiversity: Empirical Progress and Theoretical Extensions. Princeton: Princeton University Press, 2002: 151- 166.
[16]邓辅唐, 吕小玲, 喻正富, 孙珮石, 晏雨鸿. 高速公路边坡植物群落的水土保持效应. 中国水土保持, 2007, (5): 41- 46.
[17]王英宇, 宋桂龙, 韩烈保, 李义强, 徐洪雨, 赵斌. 京承高速公路岩石边坡植被重建3年期群落特征分析. 北京林业大学学报, 2013, 35(4): 74- 80.
[18]张全国, 张大勇. 生物多样性与生态系统功能: 最新的进展与动向. 生物多样性, 2003, 11(5): 351- 363.
Effect of life form composition of indigenous plant species on soil and water conservation on slopes in Sichuan-Chongqing region
PAN Shengwang1,*, YUAN Xin2, LEI Zhihua3, HU Mingcheng1
1FacultyofUrbanandRuralConstruction,ChengduUniversity,Chengdu610106,China2DeptofNationalDefenseArchitecturePlanning&EnvironmentalEngineering,LogisticalEngineeringUniversityofPLA,Chongqing401131,China3SchoolofMedicalLaboratorialTechnics,XinyangVocational&TechnicalCollege,Xinyang464000,China
Reconstruction of plant communities on superhighway side slopes using native shrubs is an important ecological practice that is employed for stabilization of the slopes. To advance the ecological restoration process of slope vegetation, we evaluated the impacts of the life form composition of indigenous plant species on soil and water conservation properties on the slopes using three representative ecosystems,Miscanthussinensis+Buchloedactyloides,Neosinocalamusaffinis+Indigoferapseudotinctoria, andCupressusfunebris+Leucaenaleucocephala, in southwest China. Four ecological models represented by tree-, shrub-, and herb-dominated experimental plots (AAXi, SSXi, and HHXi, respectively), which were composed of one of the three ecosystems and one of the indigenous dominant species (Buchloedactyloides,Loliumperenne,Vitexnegundo,Amorphafruticosa,Ailanthusaltissima, andRobiniapseudoacacia), and by herb-shrub-tree-dominated experimental plots (HSA), which were composed of three dominant species, were reconstructed via a three-species assembly experiment in April, 2009. The generation of surface runoff and soil erosion were investigated in the plots and the correlation between the life forms of indigenous species, species diversity, and levels of soil and water conservation in the ecosystems were analyzed from 2010 to 2014. The results showed that the fluctuation in plant species richness between communities of the same age decreased in the order of AAXi> SSXi> HAS > HHXifrom 2010 to 2014. Shannon-Wiener index and Pielou′s index indicated that life forms of the indigenous species growing on the slopes affected plant species diversity, which fluctuated with community phases. From 2010 to 2011, these indices varied in the following order HHXi> HSA > SSXi> AAXi,and from 2012 to 2014, they varied in the order of HSA > HHXi> SSXi> AAXi. Of the four ecological models tested, the runoff coefficient and erosion in the period from 2010 to 2011 was the lowest in HHXiplots, followed by HSA plots, whereas from 2012 to 2014, the HSA plots manifested a more prominent soil and water conservation compared to other ecological models, while these properties continued being the lowest in HHXiplots. The plant species diversity of the slope vegetation was closely related to its capacity for soil and water conservation as indicated by the determination coefficient of more than 0.951; the higher species diversity in a given community, the smaller the runoff coefficient and erosion modulus. The results from this study suggested that the life form of indigenous species plays an important role in increasing plant species diversity and improving soil and water conservation properties of side slopes.
indigenous plant; life form; species diversity; soil and water conservation
国家自然科学基金项目(31300439)
2015- 01- 12; 网络出版日期:2015- 11- 16
Corresponding author.E-mail: panwang@swu.edu.cn
10.5846/stxb201501120094
潘声旺,袁馨,雷志华,胡明成.乡土植物生活型构成对川渝地区边坡植被水土保持效益的影响.生态学报,2016,36(15):4654- 4663.
Pan S W, Yuan X, Lei Z H, Hu M C.Effect of life form composition of indigenous plant species on soil and water conservation on slopes in Sichuan-Chongqing region.Acta Ecologica Sinica,2016,36(15):4654- 4663.
