寄生对集团内捕食系统中物种入侵的影响
2016-10-24王红,苏敏,潘峰
王 红,苏 敏,潘 峰
合肥工业大学数学学院, 合肥 230009
寄生对集团内捕食系统中物种入侵的影响
王红,苏敏*,潘峰
合肥工业大学数学学院, 合肥230009
寄生已被证明是影响生物入侵动态的关键因素,可通过直接或间接作用影响物种间相互作用。基于元胞自动机模型,探讨了寄生感染对宿主种群的间接作用(寄生对宿主的密度调节和特征调节效应)在外来种入侵动态中的作用。结果显示,寄生对宿主的3种调节效应均对外来种的入侵起到了抑制作用;同时,相对于密度调节效应,寄生的特征调节对捕食者种群入侵的抑制作用更明显,而两者的耦合调节效应对入侵的抑制能力明显高于单种效应的作用。综上,该研究在一定意义上丰富和发展了生物入侵的理论,为生物入侵的有效控制提供了理论参考。
生物入侵;寄生;密度调节;特征调节;间接效应;集团内捕食系统;元胞自动机模型
自然系统中生物入侵现象已非常普遍,并严重威胁了生态系统功能与可持续性,从而引发了一系列的生态问题,导致了生物多样性的丧失[1-2]。生物入侵带来的危害主要表现在:(1)与本地物种产生资源竞争、捕食等关系,造成本地物种减少甚至灭绝[3-4];(2)外来种携带各种致病病毒或潜在病原体,通过不同途径传播给本地生物种群并导致灭绝[5-6];(3)与本地物种杂交并逐渐取代本地种,造成本地种的基因灭绝或改变本地种的进化方向[7]。因此,为了有效控制或减缓生物入侵,必须充分了解影响生物入侵的关键因素及其内在机制。
大量研究表明,入侵种可能与本地种之间存在集团内捕食作用,并且入侵种处在捕食者的营养级水平,其与本地种之间存在的竞争与捕食关系显著威胁了本地种的长期续存[8-9]。同时,寄生感染被认为是决定入侵结果的关键性因素,在生物入侵中占据着重要地位[10- 12]。寄生能够通过调节宿主间的竞争、捕食等行为特征,从而对与本地种之间存在竞争或捕食关系的外来种入侵动态产生重要影响[13]。近期研究表明,集团内捕食在入侵和生物控制中的重要性已经越来越明显,而寄生作用能充分改变集团内捕食种群的动态,在某些情况下能让本地种续存,但在另一些情况中会加速其灭绝[13-14]。爱尔兰的片脚类水生入侵系统恰好印证了寄生在集团内捕食系统中的重要性[13]。本地片脚类物种G.d.Celticus和3个入侵片脚类之间可能发生竞争和集团内捕食作用。而寄生虫P.mulleri专攻本地片脚类物种,它对宿主生存没有直接影响,但会因为特征调节的间接效应而有利于外来种的入侵。
同时,空间结构已被证明是影响疾病传播的重要因素之一,尤其对于通过感染与健康个体的直接接触而发生传播的传染性疾病[15]。然而,有关寄生调节的相互作用在空间尺度上对生物入侵影响的研究目前还比较少。本文将以G.pulex/G.d.Celticus系统[13]为研究背景,采用元胞自动机模型来探讨寄生调节的特征效应、密度效应及其耦合作用对具有空间结构的外来捕食种群入侵动态的影响。
1 模拟模型
为了讨论寄生感染对入侵动态的影响,构建了寄生调节的外来捕食种群入侵本地食饵种群的空间动态模型。模型假设入侵种与本地种与基础资源形成了集团内捕食系统,其中捕食率较高的入侵种称为捕食者(IGPredator),而捕食率较低的本地种称为食饵(IGPrey)。寄生能够调节已感染宿主的死亡率[15],即寄生导致的宿主额外致死率称为寄生的密度调节;寄生作用也可能会导致已感染宿主捕食能力下降,并且其被捕食的概率将会提高,这称为寄生的特征调节。文中用参数f,k来表示(具体参数意义见表1)。
在元胞自动机模型中,将生境划分为100×100的网格,每个网格的可能状态有五种:空斑块(O),本地健康食饵(S1),本地感染食饵(I1),入侵的健康捕食者(S2),入侵的感染捕食者(I2),同时采取同步更新以及摩尔邻体结构(八邻体)。具体的模拟规则如下:1)初始时刻,生境中只有本地食饵,并且随机分布在100×100的网格中,最终达到稳定的平衡状态;2)外来种携带病毒入侵该本地种群(模拟时引入3×3个外来种,放在网格的中心部分),并且病毒可寄生于本地种,假设由于免疫反应,病毒对本地种的危害较大。外来种与本地种之间的相互作用如下:①资源竞争,本地种和外来种的个体都以一定的概率侵占空斑块,但是本地食饵种群侵占能力强于外来捕食者种群(假设相同物种的健康与已感染个体的侵占能力相同);②捕食作用,本地种与入侵种之间相互捕食,感染种因寄生的特征调节使其捕食其他物种的能力减弱(假设感染种捕食健康种和感染种的可能性相同),同时被健康种捕食的可能性会提高;③种间与种内感染,假设病毒是单纯的水平传播,即被感染个体产生的后代都是健康的,并且种内个体间的感染强度高于种间感染;3)死亡过程,物种个体除了以概率mi自然死亡外,寄生引起的密度调节效应将会导致已感染个体额外的死亡率μi。t时刻每个斑块的状态转移规则见表2[16-17]。

表1 各参数意义及取值
参数下标1指的是本地的食饵,2指的是入侵的捕食者
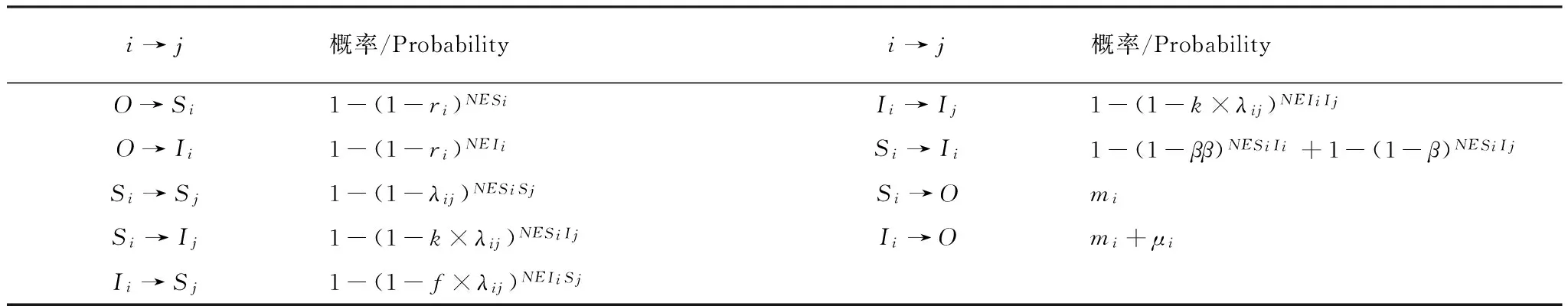
表2 网格模型中斑块转移的概率
NESi和NEIi分别表示空斑块周围邻体中健康的和感染的食饵或捕食者个体数量,所以(1-ri)NESi和(1-ri)NEIi分别表示未被健康种和感染种侵占的空斑块,则1-(1-ri)NESi和1-(1-ri)NEIi分别表示被健康种和感染种侵占的空斑块;NESiSj和NESiIj分别表示健康种i周围邻体中健康种j和感染种j的个体数; NEIiSj和NEIiIj分别表示感染种i周围邻体中健康种j和感染种j的个体数;NESiIi表示健康种i周围邻体中感染种j的个体数(i,j=1,2,且i≠j)(捕食概率和感染概率的原理同侵占概率)
2 结果分析
2.1寄生调节效应下的物种空间分布
基于元胞自动机模型,在一组参数λ12和ββ(这组参数下,本地种与外来种最终是达到平衡状态)下,分析4种状态(无寄生状态,寄生的密度调节,特征调节及其耦合作用)下本地食饵和入侵捕食者的空间分布及其时间动态。从动态图可以看出如下结果:(1) 4种状态下入侵捕食者的密度均随着时间变化呈现递增趋势,而本地食饵密度却减小,且两物种在平衡时刻将达到共存(图1);(2) 4种状态下物种平衡时刻的密度并不相同,其中无寄生的情况下,本地食饵密度最小,而入侵的捕食者种群密度最大,这说明寄生的存在对捕食者的入侵产生抑制作用;(3) 4种不同效应对外来种入侵动态影响不同。初始时刻,本地食饵随机分布在100×100的网格中,系统达到平衡状态后引入外来捕食者。随着时间的增加,由于捕食和感染作用,捕食者向四周扩散,产生环形波,并且捕食者总是围绕着食饵,而其中的感染种总是围绕着健康种。最后,入侵种完全扩散到边界,与本地种达到平衡状态。从空间分布图可以看出,同一时刻入侵种的扩散波大小不同,不同效应会影响入侵种的扩散速度。
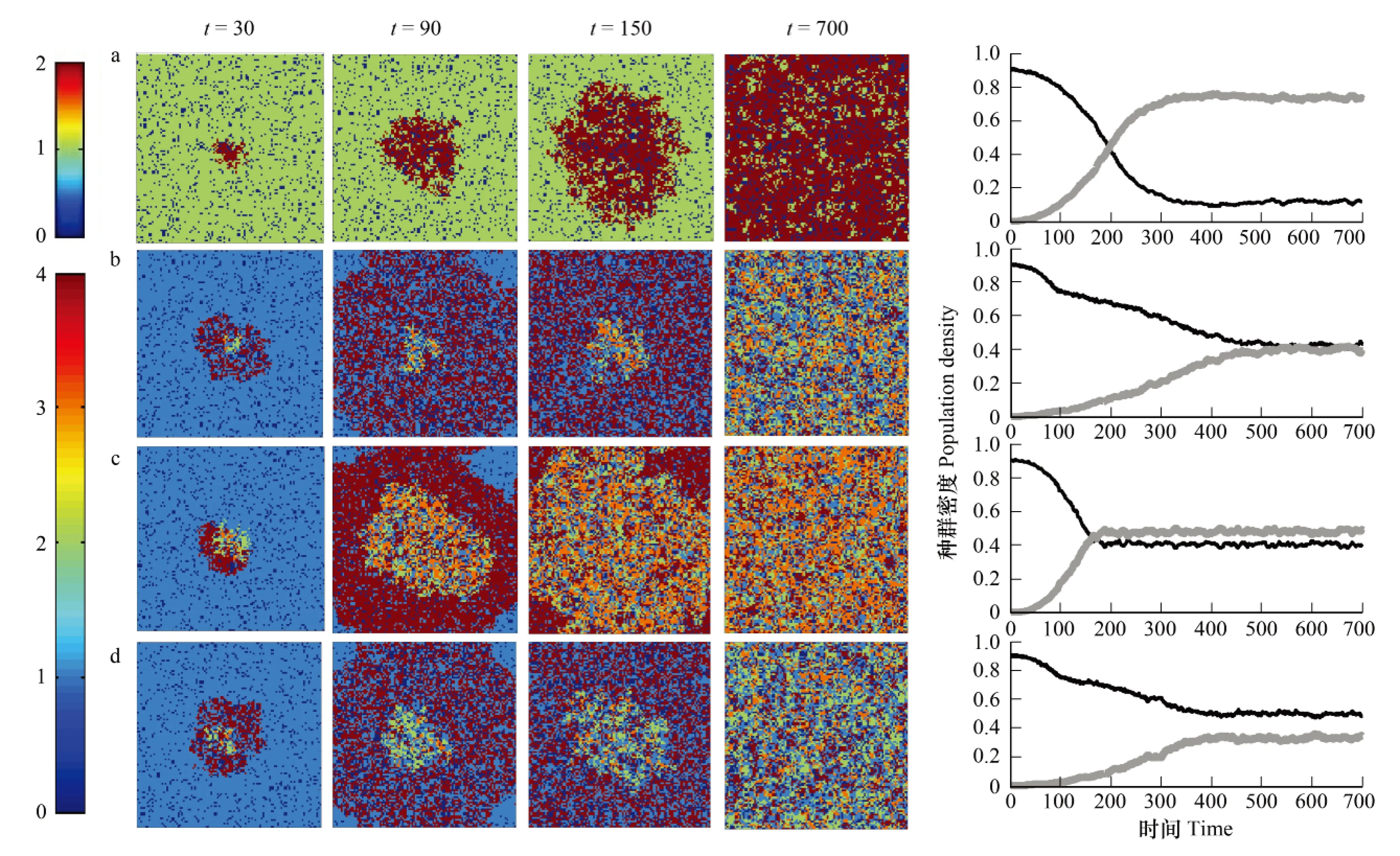
图1 100×100的网格中,4种状态下物种在不同时刻的空间分布情况和时间动态图Fig.1 Spatial patterns at various time steps and the temporal dynamics of each species under four status in a 100×100 lattice(a)无寄生状态;(b)只有密度调节的影响(μ2=0.1,μ1=0.3;f=k=1);(c)只有特征调节的影响(μ2=μ1=0;f=1.5,k=0.5);(d)同时有密度调节和特征调节的影响(μ2=0.1,μ1=0.3;f=1.5,k=0.5);(a—d)中,λ12=0.1;(b—d)中,ββ=0.25;黑色实线表示本地食饵,灰色实线表示入侵的捕食者;(a)中,深蓝色表示空斑块,绿色表示本地种,深红色表示入侵种;(b—d)中,深蓝色表示空斑块,浅蓝色表示本地的健康食饵,绿色表示入侵的健康捕食者,橙色表示入侵的被感染捕食者,深红色表示本地的被感染食饵
2.2寄生调节效应下物种入侵动态对捕食率的响应
捕食强度对集团内物种共存的影响非常重要[12-13],因此本文探索了无寄生以及3种不同寄生调节的间接效应下种群密度、入侵结果以及共存区域对捕食率λ12的响应机制。模拟结果显示,系统可能会出现3种结果:外来种入侵不成功,系统只有本地种;外来种成功入侵且两物种共存;外来种成功入侵,并且完全替代本地种。4种情形下,外来捕食者的密度随捕食率的增大均呈逐渐增大并最终平衡的趋势,而本地食饵的密度却是逐渐减少直至灭绝(图2)。首先,通过比较图2,可以发现:(1) 在无寄生的情形下,外来捕食者随着捕食率的增加,很快能够成功入侵,并在较小的捕食率区间和本地种食饵达到共存,然而由于过强的捕食作用最终将导致本地种的灭绝,入侵种完全替代本地种(图2);而在寄生存在的情形下,共存区域显著增大,同时入侵种完全替代本地种的区域减小(图2)。
(2)结果揭示寄生同时具有密度和特征调节的情形下,入侵种和本地种的共存区域最大,在特征调节的间接影响下,共存区域的范围次之,而在密度调节的情形下外来种与本地种关于捕食率的共存区域最为狭窄。而对比无寄生的结果,可以清楚的发现两者的耦合作用产生的结果与无寄生状态的差异最为明显(图2)。这一现象阐明,密度调节产生的宿主额外死亡以及特征调节引起捕食率变化二者的耦合作用导致外来种入侵动态发生更为显著的改变。
(3)结果显示种群的密度及其流行趋势(Prevalence)对捕食率的响应也非常显著(图2)。寄生在本地种与入侵种内部的流行随着捕食强度的改变分别呈现递减和递增的趋势(图2)。产生这一现象的主要原因在于:随着捕食率λ12的增大,已感染食饵更容易被捕获,而食饵密度的下降直接有利于集团内入侵的捕食者对资源的侵占,一方面导致捕食者的种群数量增加,提高了捕食者内部相互接触感染的几率,另一方面食饵密度的下降致使食饵对捕食者的捕食降低,那么最终引起已感染捕食者的相对比例随着捕食率的增大呈现递增的趋势。
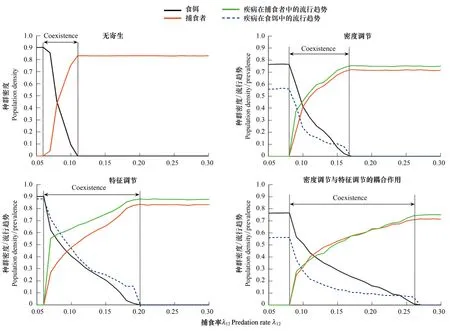
图2 寄生虫在不同捕食率下通过密度调节和特征调节对平衡时刻物种密度的影响Fig.2 Effects of parasitism on species′ density under different rate of predation by density effect and trait effect
2.3寄生调节效应下感染率对物种入侵动态的影响
种内感染率(ββ)是疾病传播的主要影响因素,本文探索了3种间接效应下,ββ对平衡时刻本地种(IGPrey)和入侵种(IGPredator)密度的影响(图3)。由图可知,(1)在两种带有密度调节的状态下,本地食饵与外来捕食者的密度随着ββ的改变分别呈现递增和递减的趋势,最后两个物种密度达到平衡。(2)寄生感染率的增大将导致入侵更为困难,从而抑制了外来种的入侵。(3)疾病在本地种与入侵种内部的流行随着种内感染率的改变都是呈现逐渐增加的现象(图3)。这一现象产生的原因很显然,即种内感染率的增加影响到已感染捕食者和食饵在种群内比例的增大。而在图3中,ββ>0.5时,疾病在入侵种内部的流行变为0,这是因为当ββ过大时,密度和特征调节的耦合作用导致外来种不能成功入侵,使得外来种的种群密度为0。
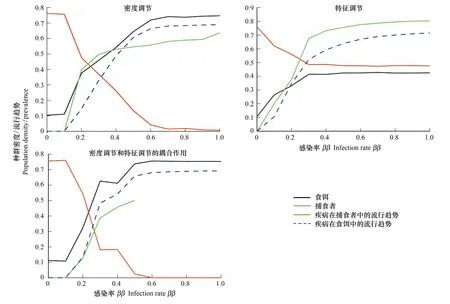
图3 寄生虫在不同种内感染率ββ下通过密度调节和特征调节对平衡时刻物种密度的影响Fig.3 Effects of parasitism on species′ density under different infectious rate between the same species by density effects and trait effects
3 讨论
本文构造了涵盖物种间相互竞争同时又相互捕食的空间集团内捕食系统,并在此基础上引入寄生调节的生物入侵模型。基于元胞自动机模型,模拟了本地种与外来种在4种状态下随着时间变化的空间分布。模拟结果可以看出,空间结构不是像寄生-宿主系统中的那样静态自组织,而是一种动态化的自组织空间结构。这种自组织不仅引起时间动态上的波动,也可以在空间生境上形成环状分布和扩散波。
本文的另一个重要研究结果表明,寄生对宿主的密度与特征调节效应对生物入侵有着重要的影响,共享的寄生虫可抑制外来捕食种群的成功入侵。同时,研究表明寄生能够增加集团内捕食系统中本地种与入侵种的共存区域,这与Hatcher等[18]基于非空间模型所得研究结果相一致。同时,进一步揭示了寄生调节的特征效应相比于寄生的密度调节效应对宿主群落产生的作用更为显著,而两者的耦合作用对入侵的调节明显强于单种效应的作用。
上述结果对于生物控制具有重要的启迪作用,在实际的生物控制中,可以通过引入传染性较强的寄生病毒等措施来抑制外来种的成功入侵。然而,模型假设疾病只能水平传播,并且基于易感染-已感染(SI)模型。而在实际生物学中,由于遗传效应,很多疾病在宿主中可能产生垂直传播,并且已感染个体还可能会恢复(即SIR模型)。已有研究表明,宿主-寄生系统中SIR模型可导致已感染宿主的密度呈现马鞍式变化趋势,这一结果与SI模型产生的动态存在显著差异[19]。因此,未来的研究应该整合群落生态,病毒进化和免疫等理论框架来进一步构建更贴合自然现象的寄生调节的生物入侵模型,从而更好的理解和管理外来种的入侵机制。
[1]Oliveira N M, Hilker F M. Modelling disease introduction as biological control of invasive predators to preserve endangered prey. Bulletin of Mathematical Biology, 2010, 72(2): 444- 468.
[2]Kolar C S, Lodge D M. Progress in invasion biology: predicting invaders. Trends in Ecology Evolution, 2001, 16(4): 199- 204.
[3]陈兵, 康乐. 生物入侵及其与全球变化的关系. 生态学杂志, 2003, 22(1): 31- 34.
[4]Ludsin S A, Wolfe A D. Biological invasion theory: Darwin′s contributions fromtheoriginofspecies. Bioscience, 2001, 51(9): 780- 789.
[5]高增祥, 季荣, 徐汝梅, 谢宝瑜, 李典谟. 外来种入侵的过程、机理和预测. 生态学报, 2003, 23(3): 559- 570.
[6]Heger T, Trepl L. Predicting biological invasions. Biological Invasions, 2003, 5(4): 313- 321.
[7]Rosenheim J A, Kaya H K, Ehler L E, Marois J J, Jaffee B A. Intraguild predation among biological-control agents: theory and evidence. Biological Control, 1995, 5(3): 303- 335.
[8]Prenter J, MacNeil C, Dick J T A, Dunn A M. Roles of parasites in animal invasions. Trends in Ecology and Evolution, 2004, 19(7): 385- 390.
[9]Yang Y H, Li W D, Wang G. Can we use disease to control biological invasion?—A theoretical research. Ecological Modelling, 2014, 277: 97- 107.
[10]Dunn A M, Torchin M E, Hatcher M J, Kotanen P M, Blumenthal D M, Byers J E, Coon C A C, Frankel V M, Holt R D, Hufbauer R A, Kanarek A R, Schierenbeck K A, Wolfe L M, Perkins S E. Indirect effects of parasites in invasions. Functional Ecology, 2012, 26(6): 1262- 1274.
[11]Hatcher M J, Dick J T A, Dunn A M. Disease emergence and invasions. Functional Ecology, 2012, 26(6): 1275- 1287.
[12]Strauss A, White A, Boots M. Invading with biological weapons: the importance of disease-mediated invasions. Functional Ecology, 2012, 26(6): 1249- 1261.
[13]Tompkins D M, White A R, Boots M. Ecological replacement of native red squirrels by invasive greys driven by disease. Ecology Letters, 2003, 6(3): 189- 196.
[14]Hatcher M J, Dick J T A, Dunn A M. How parasites affect interactions between competitors and predators. Ecology Letters, 2006, 9(11): 1253- 1271.
[15]Hatcher M J, Dick J T A, Dunn A M. A key stone effect for parasites in intraguild predation? Biology Letters, 2008, 4(5): 534- 537.
[16]Su M, Hui C, Zhang Y Y, Li Z Z. How does the spatial structure of habitat loss affect the eco-epidemic dynamics? Ecological Modelling, 2009, 220(1): 51- 59.
[17]Su M, Hui C, Zhang Y Y, Li Z Z. Spatiotemporal dynamics of the epidemic transmission in a predator-prey system. Bulletin of Mathematical Biology, 2008, 70(8): 2195- 2210.
[18]Hatcher M J, Dick J T A, Dunn A M. Parasites that change predator or prey behaviour can have keystone effects on community composition. Biology Letters, 2014, 10(1): 20130879.
[19]Roy M, Holt R D. Effects of predation on host-pathogen dynamics in SIR models. Theoretical Population Biology, 2008, 73(3): 319- 331.
Effects of parasitism on biological invasion in intraguild predation
WANG Hong, SU Min*, PAN Feng
SchoolofMathematics,HefeiUniversityofTechnology,Hefei230009,China
Biological invasions are a global threat to community structure and biodiversity, and it is therefore necessary to understand the fundamental mechanisms of biological invasions. The success of a biological invasion is not only related to the biology and ecological characteristics of the invaders, but it also depends on the invaded community′s vulnerability. A number of experiments have shown that parasitism affects biological invasion both directly and indirectly, but how these two key redistribution processes of biological invasion and disease emergence are interlinked is a relatively new research topic. In this study, we used a cellular automata simulation model to explore how the indirect effects of parasitism (including density-mediated effects and trait-mediated effects) influence the spatial invasion dynamics of an exotic predator. In the spatial model, the native species and invasive species form an intraguild predation with their basal resource. First, we explored the spatial distribution patterns of the native and invader species with and without parasite-mediated effects. Overall, the distribution patterns formed spatial traveling waves. Along the spatial wave from the front to the back, the susceptible native prey was first followed by the infected ones, then by the susceptible invader predators, and finally by the infected invader predators. Due to host-parasite and predator-prey interactions, the parasites have to encircle susceptible hosts for their possible survival, whereas the predators need to chase after the prey for survival. Second, three main coexistence results were obtained under different parasite effects: invade successfully, coexist with local species, and invade unsuccessfully. We mainly focused on the effects of the exotic predator′s predatory rate on native prey parameters (λ12) and the intraspecific infection rate (ββ) on the invasion of invader predator species. Without parasites, the exotic predator can invade successfully and quickly by increasing λ12, whereas the coexistence region for invasive predator and native prey species is very small. However, the coexistence region expands remarkably when parasites are present. Furthermore, the impact of trait-mediated indirect effects on the invasion of predator species was more dramatic than density-mediated effects. Moreover, the coexistence region was the largest when hosts were affected by density and trait effects simultaneously, causing the density and trait effects together to have the largest inhibiting effect on invasion. In addition, the influence of transmission rate ββ on the dynamics of alien invasions was affected by these indirect effects. In the scenarios with density and trait-mediated effects together, as well as with density-only effects, the invader became extinct as parameterββ increased to some thresholds. However, when modeling just trait-only effects, the invasion predators did not go extinct, but coexisted steadily with the native prey species. Overall, from the model, we identified conditions that inhibit exotic invasions by introducing parasites as a biological control. Our research can therefore contribute to the development of biological invasion theories, and provide a basis for the prevention and control strategies to handle biological invasions.
Invasive species; disease emergence; density-mediated; trait-mediated; indirect effect; intraguild predation; cellular automata simulation model
国家自然科学基金资助项目(31000192);中国博士后科学基金资助项目(20100481159);中央高校基本科研业务费专项基金资助(J2014HGXJ0065); 国家留学基金资助项目
2015- 07- 02; 网络出版日期:2015- 12- 08
Corresponding author.E-mail: sum04@163.com
10.5846/stxb201507021406
王红,苏敏,潘峰.寄生对集团内捕食系统中物种入侵的影响.生态学报,2016,36(15):4809- 4815.
Wang H, Su M, Pan F.Effects of parasitism on biological invasion in intraguild predation.Acta Ecologica Sinica,2016,36(15):4809- 4815.
