淹水胁迫下江南牡丹生长及光合特性研究
2016-09-27朱向涛金松恒哀建国时浩杰
朱向涛, 金松恒, 哀建国, 时浩杰
( 浙江农林大学, 浙江 临安, 311300 )
淹水胁迫下江南牡丹生长及光合特性研究
朱向涛, 金松恒*, 哀建国, 时浩杰
( 浙江农林大学, 浙江 临安, 311300 )
以3年生江南牡丹品种‘凤丹白’为材料,利用盆栽淹水法,设置正常管理、轻度胁迫和重度胁迫3个水平,研究不同淹水胁迫水平对牡丹生长和光合特性的影响。结果表明:经过30 d胁迫后,正常管理、轻度胁迫和重度胁迫下的江南牡丹苗高生长量分别为3.6、1.1和0.73 cm,地径生长量分别为0.21、0.11和0.06 cm,植株总生物量增加量分别为7.0、3.0和2.75 g,淹水胁迫和正常生长差异显著,淹水胁迫严重影响了江南牡丹的生长。同时,在正常管理时,牡丹总叶绿素含量升高,而在淹水胁迫下呈下降趋势。淹水胁迫不同时间根系活力均呈下降趋势且随着胁迫程度的增加下降越大。正常管理下光合速率逐渐增加而胁迫条件下光合速率逐渐降低。同时胁迫条件下,牡丹蒸腾速率、气孔导度均明显下降;轻度淹水胁迫下胞间CO2浓度先升高后降低;而重度胁迫下胞间CO2浓度呈现逐渐升高的变化趋势。淹水胁迫对牡丹根系活力、茎段生长和叶片光合特性影响较大。该研究结果为江南牡丹耐涝胁迫机理研究奠定了理论基础。
江南牡丹, 淹水胁迫, 根系活力, 生长, 光合作用
牡丹(Paeoniasuffruticosa)是我国特产的传统名花,属芍药科(Paeoniaceae)芍药属(Paeonia)落叶灌木,为中国十大名花之一,有“花中之王”的美誉,根系入药,具较高的药用价值(李嘉珏,1998)。江南牡丹品种群作为中国四大牡丹品种群之一,主要分布在上海、江苏南部、浙江杭州以及安徽宁国、铜陵等地(李嘉珏和何丽霞,2003)。由于江南地区降水多、气温高而牡丹具有不耐湿热的特点,目前仅存20余品种,而有些品种如‘紫云芳’、‘凤尾’等数量极少,已濒临灭绝(王佳,2009),湿热环境是制约江南牡丹生长发育的主要原因,特别是在江南地区的梅雨季节,长时间降雨导致江南牡丹处于涝害胁迫下生长受到影响。江南牡丹已有一千多年的栽培历史,使江南牡丹重现辉煌具有重要意义,而解决好耐湿热品种的选育、品种结构调整和相应的栽培问题将是重中之重。因此,探索涝害对江南牡丹影响的机制并在保护现有江南牡丹品种的基础上培育耐湿热的牡丹品种成为迫在眉睫的任务。
淹水会引起植物形态、解剖、生理、代谢等方面一系列的变化,其本质是淹水使植物根系处于无氧环境,从而引起植物体各部分生长变化(Yanagawa & Komatsu, 2012; Naumann et al, 2008; Farwell et al, 2007; Dunn & Gaydon, 2011; Voesenek et al, 2004)。光合作用是植物生长的能量基础,不同物种对淹水胁迫下光合特性响应各不相同,而淹水胁迫下牡丹的生长和光合响应尚未见报道,因此研究并掌握淹水胁迫下牡丹生长发育及光合特性规律能为牡丹耐涝机理研究提供参考,对耐涝江南牡丹品种筛选、新品种培育尤为重要。本研究以3年生江南牡丹品种‘凤丹白’为材料,研究淹水胁迫下牡丹苗高、地径、生物量、根系活力等指标及其光合特性,分析淹水胁迫对江南牡丹生长发育和光合特性的影响,为阐明江南牡丹的耐涝机理提供理论依据。
1 材料与方法
1.1 材料与处理方法
试验材料选用3年生江南牡丹品种‘凤丹白’,2012年9月引自安徽宁国,盆栽于浙江农林大学官塘基地,苗木生长期间进行正常的水肥管理,2013年6月,选取长势、大小基本相同的苗木30盆,栽植于上口径28 cm、下口径19 cm、高23 cm 的塑料盆中,基质是由园土∶沙∶珍珠岩(5∶3∶2)组成的混合基质。
利用浸盆法进行淹水处理,A水平进行正常的水肥管理,土壤含水量为田间持水量的75%左右,B水平模拟轻度水分胁迫,将花盆浸入水箱内,每天观察水面与土面的距离维持淹水高度比盆土低5 cm,C水平模拟涝害,土壤含水量过饱和,将花盆浸入水箱内,每天观察水面与土面的距离维持水面高于土面 5 cm。分别在处理后第5、10、15天测定其光合参数、第5、10、15、20天根系活力。苗高、地径、叶绿素含量、根茎叶生物量于第0、30天分别测定,所有测定完成3次重复。
1.2 生物量测定
用卷尺测定苗高(精确到0.1 cm),用游标卡尺测量地径(距离土面6 cm的高度),每株用游标卡尺交叉测定 2 次(精确到 0. 01 mm)取其平均值。生物量测定胁迫试验结束后在苗高、地径测量的基础上,从各处理随机选3株,全株挖出,洗净,分根、茎、叶3部分放入烘箱,105 ℃杀青0.5 h,然后80 ℃烘约72 h至恒重。用1/100电子天平称取各处理各株干物质质量。
苗高净生长量=第30天的苗高-第0天的苗高
地径净生长量=第30天的地径-第0天的地径
生物量净增长量=第30天的总生物量-第0天的总生物量
1.3 叶绿素含量和根系活力的测定
叶绿素含量测定在淹水胁迫的第0天和第30天采集植株顶端的第 4~5 片完全展开的叶片,去掉主脉,用于分析,每处理完成3次重复,按照李合生(2000)的方法测定。在处理后第5、10、15、20天分别用TTC还原法(邹琦,2000)测定根系活力。
1.4 光合测定
采用LI-6400便携式光合仪(LI-COR,USA),选择晴朗无风的天气于室外自然条件下进行测定,随机选择各处理下的植株各5株,每次测定固定植株顶端第5片完全展开的叶片,于上午9: 00-11: 00 进行,光照强度设定为1 000 μmol·m-2·s-1,测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和气孔限制值(Ls),其中Ls=1-Ci/Ca,Ci为胞间CO2浓度,Ca为大气CO2浓度。
1.5 数据分析
采用Excel 2007和Spss 19.0软件进行单因素方差分析(ANOVA)和多重比较(Duncan)分析。
2 结果与分析
2.1 淹水胁迫对牡丹苗高和地径生长的影响
淹水胁迫对牡丹的苗高和地径生长影响结果如表1所示,总体上来看,牡丹淹水处理对苗高和地径的净生长量有明显抑制作用。从结果来看,与对照相比,水淹胁迫显著降低苗高和地径的生长,但B、C两处理之间对株高生长的抑制效果差异不明显,而对地径的生长抑制效果显著,因此,随着淹水胁迫程度的逐渐增强,水淹胁迫对牡丹增粗生长影响较大。从第20天开始,B、C两个处理的牡丹叶片开始萎蔫并下垂,在第30天时,B处理叶片已经有部分发黄,C处理已经有叶片开始脱落。
2.2 淹水胁迫对牡丹生物量和根冠比的影响
从表2可以看出,正常管理情况下,牡丹的根茎叶生物量在30 d内有不同程度的增加,牡丹处于旺盛生长的状态,第30天时根冠比略有下降。B处理状态下,牡丹的根茎叶生物量也呈不同程度增加,但增加幅度相对较低,而此时根冠比下降较少。C处理下,叶片的生物量下降,主要是地上部分有叶片脱落,此时根系和茎段的生物量有上升,总生物量略有增加。根系由于对水涝胁迫的适应开始出现新生呼吸根而使地下部生物量上升,说明牡丹在C处理下,地下部生长加快,此时地上部分已生长受到抑制,叶片生物量降低,茎段生物量有少量增加,根系的生物量增加较大,根冠比有所增加。
2.3 淹水胁迫对牡丹叶绿素含量和根系活力的影响
如表3所示,在最初的处理阶段,牡丹叶绿素a含量相差不大。正常管理下,叶绿素a和总叶绿素在第30天时有了明显增加,叶绿素b则变化略微下降,类胡萝卜素含量和叶绿素a/b略有升高。B处理条件下,所有的色素含量和叶绿素a/b都比处理前有所下降,叶绿素a和叶绿素a/叶绿素b变化差异不明显,叶绿素b、总叶绿素和类胡萝卜素下降幅度较大,与开始时差异明显。C处理下,各个指标也是有不同程度的下降,分析结果来看,叶绿素b、总叶绿素和类胡萝卜素下降明显,而叶绿素a和叶绿素a/b下降不明显。
从图1可以看出,在不同的水涝胁迫处理下,牡丹的根系活力具有一定的变化规律,A正常管理情况下,江南牡丹的根系活力变化不明显,但在各个处理阶段都比淹水胁迫下都高。处理5 d时,B、C两个处理的根系活力均下降,但方差分析显示两者差异不明显,随着处理时间的延长,两个处理均差异显著,说明C处理下对根系活力影响较大。在B、C处理情况下,牡丹根系活力随着时间的延长呈现逐渐降低的变化趋势,处理后,根系活力明显下降,在20 d时,B、C处理的根系活力达到最低。在处理的每个时间段,根系活力均为A>B>C,说明在淹水胁迫程度越深,根系活力下降越多。

图 1 淹水胁迫下江南牡丹根系活力变化情况Fig. 1 Effects of different treatments to root activity of P. suffruticosa
2.4 淹水胁迫对牡丹光合特性的影响
光合速率随着处理时间的延长而发生变化,从图2可以看出,A正常管理条件下,光合速率随着时间的延长呈现逐渐增加的变化趋势,且在第15天时达到13.53 μmol·m-2·s-1,可见这段时间牡丹处于旺盛生长期,叶片的光合能力逐渐增强,光合速率逐渐提高。B轻度水涝胁迫下,最初第5天时,光合速率与正常管理条件下相差不大,但随着处理时间的增长,第10天时,光合速率已经相对较小,仅为0.4 μmol·m-2·s-1,在第15天时,光合速率接近于0 μmol·m-2·s-1,原因是第15天时,叶片开始出现萎蔫现象,叶片光合能力下降光合速率较低。C重度水涝胁迫下,在处理后的第5天,光合速率与正常管理条件下牡丹的光合速率相似,变化不大,没有呈现明显的下降趋势, 但是在处理后的第10天, 光合速率明显下降, 大约为0.2 μmol·m-2·s-1, 到第15

表 1 不同处理下牡丹苗高与地径变化情况
注: 表中数据为平均值 ± SE; 同一参数同一行内不同小写字母表示差异达显著水平(P= 0.01)。下同。
Note: Values are means ± SE; Different common letters of the same parameters in the same row mean significant differences according to LSD analysis atP= 0.01 level. The same below.

表 2 不同处理下牡丹生物量变化情况

表 3 淹水胁迫对牡丹叶片光合色素含量及叶绿素a/b值的影响
天时,光合速率为负值,即表示这个阶段呼吸作用要大于光合作用,本阶段江南牡丹叶片开始萎蔫并出现叶片边缘枯黄的现象。方差分析显示,处理5 d时光合速率差异性不显著,处理10 d时,正常管理与B、C两处理差异性显著,而B、C差异性不显著,处理15 d时,三个处理均差异性显著。

图 2 淹水胁迫对光合速率的影响Fig. 2 Effects of flooding stress on Pn

图 3 淹水胁迫对气孔导度的影响Fig. 3 Effects of flooding stress on Gs
气孔导度也随着淹水胁迫的加重发生一系列变化,从图3可以看出,A正常管理条件下,在第5天、第10天、第15天气孔导度变化不明显。B轻度胁迫状态下,气孔导度变化趋势明显,在第5天时,与正常管理的相比,气孔导度基本维持不变甚至有所升高,这一阶段,江南牡丹对水涝胁迫本能反应,出现气孔增大的现象。但第10天时,气孔导度大幅度下降,第15天相比第5天变化幅度较大,但与第10天的变化幅度不明显,总体上来看呈现逐渐下降的变化趋势。C处理条件下,气孔导度下降幅度更大,处理的最初阶段,气孔导度变化不大,但在第10天、第15天下降明显,总体上呈现逐渐下降的变化趋势,可见在第5天以后淹水胁迫的影响较大,之后气孔导度变化较小。总体上来看,正常管理条件下,气孔导度变化不大,B处理条件下气孔导度的下降幅度小于C处理条件下的下降幅度,可见气孔导度随着胁迫程度的加深而下降越大。方差分析显示,处理5 d时,三处理气孔导度差异不显著,处理10 d和15 d时,三处理均差异性显著。
淹水胁迫下牡丹胞间CO2浓度变化规律如图4所示,正常管理条件下,胞间CO2浓度变化不大,这与气孔导度变化规律基本一致,说明此时牡丹的气孔处于正常开放状态,胞间CO2没有受到气孔开度的影响。在B胁迫条件下,胞间CO2浓度呈现先升高后降低的变化规律,但是变化幅度不大,与气孔开度的变化规律不一致,可见影响胞间CO2浓度的因素不仅是气孔导度,可能与植株的适应性有很大关系,也就是说光合作用下降可能是非气孔限制因素导致的。C处理条件下,胞间的CO2浓度反而呈现出逐渐上升的变化趋势,与气孔导度的下降趋势相反。处理5 d时,正常管理胞间CO2浓度与B、C处理差异显著,B、C处理之间差异不明显;处理10 d时,三者差异不显著;处理15 d时,正常管理与处理B差异不显著,B、C差异显著。

图 4 淹水胁迫对胞间CO2浓度的影响Fig. 4 Effects of flooding stress on Ci

图 5 淹水胁迫对蒸腾速率的影响Fig. 5 Effects of flooding stress on Tr
淹水胁迫下,牡丹蒸腾速率变化也呈现一定规律性(图5)。正常管理条件下,蒸腾速率变化不大,基本呈现逐渐下降的趋势。在B处理条件下,在第5天时,比正常管理条件下蒸腾速率提高明显,可能本阶段水分吸收加快,蒸腾速率提高,而第10天和第15天时,蒸腾速率相比第5天时下降幅度较大,且变化不明显。C处理下呈现和B 处理下相似的变化趋势,第10天比第5天呈现大幅度的下降,第15天比第5天下降幅度较大,但是第10天和第15天蒸腾速率差异不显著。
从图6可以看出,处理5 d时,气孔限制值随着处理程度的加深而逐渐增大,因此此时主要是气孔原因影响光合速率。在处理10 d、15 d时,气孔限制值随着处理程度的加深而呈现下降的变化趋势,处理后期主要非气孔限制原因造成光合速率下降。
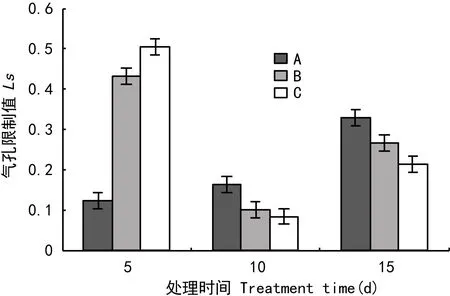
图 6 淹水胁迫对气孔限制值的影响Fig. 6 Effects of flooding stress on Ls
3 讨论
3.1 淹水胁迫对牡丹表型和根系活力的影响
淹水胁迫下牡丹苗高、地径的生长速率比正常管理有所降低,主要由于淹水胁迫下,土壤缺氧,根系活力下降,根系水分饱和使牡丹根系周围被水包围,导致通气不足从而出现“水套现象”(余叔文和汤章成,1998)。本研究中,淹水胁迫下牡丹根系活力的显著降低,说明在淹水过程中,牡丹根系生长受到抑制。当土壤相对含水量升高到一定水平时,根系生长发育所需的ATP主要由根系通过无氧呼吸提供,因此能量供不应求,加上植物在缺氧环境下能产生一些代谢毒素和诱发膜脂过氧化作用,从而抑制根系的正常发育,严重时导致根尖细胞的结构和功能受损,造成根系死亡。正常条件下,根冠比呈现降低趋势,而在淹水胁迫下,根冠比逐渐增加,说明淹水胁迫对地上部分生长的影响大于对地下部分生长的影响(曹福亮等,2010)。在淹水胁迫后期,能看到牡丹地下部分有许多白色须根出现,在一定程度上增加了地下干物质的量。白根产生的原因可能是水涝胁迫造成牡丹根系缺氧,为了能够更好地吸收更多的氧气,满足能量代谢的需要,会产生许多白色的不定根,不定根的出现在一定程度上增加了根的呼吸面积,从而加强根系对氧气的需要。因此,在缺氧条件下,耐涝植物通过产生不定根,增加根系中氧气的浓度,从而消耗更多的光合产物(汪贵斌等,2010)。与正常管理相比,淹水胁迫下,牡丹的生物量增长降低明显,与Chen et al(2010)对香蒲的研究结果相似。
3.2 淹水胁迫对牡丹叶绿素含量的影响
光合色素参与光合作用过程中光能的吸收、传递和转化,光合色素含量的降低使光能转化和能量供应能力降低,从而影响光合作用的进行。叶绿素含量是植物叶片光合性能、营养状况和衰老程度的直观表现,其含量是鉴定涝害的有效指标。水涝胁迫下,植物体内的叶绿素的合成与分解之间的平衡受到影响,叶绿素含量降低,光合作用受到影响。叶绿素含量降低是植物叶片对淹水胁迫的生化响应(张柔等,2012)。在本研究过程中,正常管理条件下,光合色素含量基本呈现升高的变化趋势。在水涝胁迫下,叶绿素a、叶绿素b、类胡萝卜素均有不同程度的下降。光合色素的降低会影响光合作用的进行(张斌斌等,2013)。潘向燕等(2006)对杂种鹅掌楸的研究发现在淹水4 d时叶片即出现失绿现象,继续淹水则叶片成黄褐色,在落羽杉及绒毛白蜡的研究也得到了类似的结论(侯嫦英,2003)。对水稻孕穗期叶片Chl a/Chl b比值研究发现,淹水导致Chl a/Chl b比值上升,表明叶绿素b受影响大于叶绿素a(利容千和王建波,2002)。模拟淹水胁迫27 d后,枫杨和栓皮栎Chl a/Chl b下降,认为淹水胁迫导致对淹水敏感的植物叶片光合反应中心的降解程度要高于捕光蛋白色素复合物的降解(衣英华等,2006)。本研究结果中,牡丹在轻度水涝胁迫下Chl a/Chl b不变,而重度水涝胁迫下Chl a/Chl b升高,原因在于叶绿素a 更多地结合在光系统反应中心上,叶绿素b主要结合在捕光蛋白复合物上(李艳等,2013)。由此可见,淹水胁迫影响了牡丹砧木叶片叶绿素的整体水平,使叶绿素不同形态间的比值发生改变,进而影响叶片的光合作用。
3.3 淹水胁迫对牡丹光合特性的影响
淹水胁迫对牡丹光合速率和蒸腾速率影响较大,本研究中,光合速率、气孔导度和蒸腾速率基本呈现下降的变化趋势。随着胁迫程度的加深,下降越明显。除了叶绿素含量降低的影响之外,光合速率降低的原因首先是根系缺氧导致不能有效吸收水分和矿质营养,限制了光合作用的进行。植物光合作用受到抑制的原因包括两个方面,一是气孔限制,气孔的关闭使叶绿体内CO2的供应受阻;二是非气孔限制,叶肉细胞光合能力的下降,使叶肉细胞利用CO2的能力降低,从而使胞间CO2含量升高(Farquhar & Sharkey,1982)。胞间CO2浓度并不随着气孔导度降低而下降,反而有所升高,按照Farquhar & Sharkey(1982)的观点,即只有当胞间CO2浓度的降低和气孔限制值增大时,才可以做出光合速率的下降是由气孔导度的下降所引起的结论。本研究中,淹水处理10 d后,气孔导度大幅度下降,但胞间CO2浓度并没有下降,这表明牡丹在淹水后期光合速率的下降主要是由非气孔限制所致。
CAO FL,CAI JF,WANG GB,et al, 2010. Effects of waterlogging stress on the growth and photosynthesis ofSapiumsebiferum[J]. Sci Silv Sin, 46(10):57-61. [曹福亮,蔡金峰,汪贵斌,等, 2010. 淹水胁迫对乌桕生长及光合作用的影响 [J]. 林业科学,46(10):57-61.]
CHEN HJ,ZAMORANO MF,IVANOFF D, 2010. Effect of flooding depth of growth,biomass,photosynthesis,and chlorophyll fluorescence ofTyphadomingensis[J]. Wetlands,30: 957-965.
DUNN BW, GAYDON DS, 2011. Rice growth, yield and water productivity responses to irrigation scheduling prior to the delayed application of continuous flooding in south-east Australia [J]. Agric Water Manag, 98(12):1 799-1 807.
FARWELL AJ, VESELY S, NERO V, et al, 2007. Tolerance of transgenic canola plants (Brassicanapus) amended with plant growth-promoting bacteria to flooding stress at a metal-contaminated field site [J]. Environ Poll, 147(3):540-545.FARQUHAR GD, SHARKEY TD, 1982. Stomatal conductance and photosynthesis [J]. Ann Rev Plant Physiol, 33: 317-345.
HONGJUN CHEN, MANUEL F. ZAMORANO,DELIA IVANOFF, 2010. Effect of flooding depth on growth, biomass, photosynthesis, and chlorophyll fluorescence ofTyphadomingensis[J]. Wetlands, 30:957-965.
HOU CY, 2003. Effeets of waterstress on the one-year seedling growth and physiology properties ofPterocetistatarinowii,TaxodiumdistichumandFraxinusvelutina[D]. Nanjing: Nanjing Forestry University. [侯嫦英, 2003. 水分胁迫对青檀等树种生长及生理特性的影响 [D]. 南京: 南京林业大学.]
LI HS, 2000. Plant physiology and biochemistry test principles and techniques [M]. Beijing: Higher Education Press. [李合生, 2000. 植物生理生化试验原理和技术 [M]. 北京: 高等教育出版社.]
LI JJ, HE LX, 2003. JiangnanP.suffruticosahistory, species composition, suitable land and spend issues [J]. Chin Flower Gard, (12):8-10. [李嘉珏, 何丽霞, 2003. 江南牡丹发展历史、品种构成与适地适花问题 [J]. 中国花卉园艺,(12):8-10.]
LI JJ, 1998. Chinese tree peony and herbaceous peony [M]. Beijing: China Forestry Publishing House. [李嘉珏, 1998. 中国牡丹与芍药 [M]. 北京: 中国林业出版社.]LI RQ,WANG JB, 2002. Plant cells and physiological stress [M]. Wuhan: Wuhan University Press. [利容千,王建波, 2002. 植物逆境细胞及生理学 [M]. 武汉: 武汉大学出版社.]
LI Y,FU YD,MA YC,et al. 2013. Effects of waterlogging on characteristics of growth and photosynthesis in different grape rootstocks [J]. Sci Agric Sin, 46(5):995-1 004. [李艳,付艳东,马艳春,等. 2013. 淹水对不同葡萄砧木生长及光合特性的影响 [J]. 中国农业科学, 46(5):995-1 004.]
LI Y,DU YP,FU YD,et al, 2013. Physiological responses of waterlogging on different rootstock combinations of cabernet sauvignon grape [J]. Acta Hortic Sin,40(11):2 105-2 114. [李艳,杜远鹏,付艳东,等, 2013. 不同砧木嫁接的赤霞珠葡萄对淹水的生理响应 [J]. 园艺学报,40(11):2 105-2 114.]NAUMANN JC, YOUNG DR, ANDERSON JE, 2008. Leaf chlorophyll fluorescence, reflectance, and physiological response to freshwater and saltwater flooding in the evergreen shrub,Myricacerifera[J]. Environ Exp Bot, 63(1):402-409.
PAN XY, 2006. Responses of different clones of hybrid tulip tree to flooding stress [D]. Nanjing: Nanjing Forestry University. [潘向艳, 2006. 杂交鹅掌楸不同无性系对淹水胁迫的反应 [D]. 南京: 南京林业大学.]
VOESENEK LACJ, RIJNDERS JHGM, PEETERS AJM, et al, 2004. Plant hormones regulate fast shoot elongation under water: from genes to communities [J]. Ecology, 85: 16-27.
WANG GB,CAO FL,ZHANG XY, et al, 2010. Effects of waterlogging on the growth and energy-metabolic enzyme activities of different tree species [J]. Chin J Appl Ecol, 21(3):590-596. [汪贵斌,曹福亮,张晓燕, 等, 2010. 涝渍胁迫对不同树种生长和能量代谢酶活性的影响 [J]. 应用生态学报, 21(3):590-596.]
WANG J, 2009. Genetic diversity ofPaeoniaosstiand germplasm resources of treeP.suffruticosacultivars from Chinese Jiangnan area [D]. Beijing: Beijing Forestry University. [王佳, 2009. 杨山牡丹遗传多样性与江南牡丹品种资源研究 [D]. 北京: 北京林业大学.]
YANAGAWA Y, KOMATSU S, 2012. Ubiquitin/proteasome-mediated proteolysis is involved in the response to flooding stress in soybean roots, independent of oxygen limitation [J]. Plant Sci, 185-186: 250-258.
YI YH,FAN DY,XIE ZQ, et al, 2006. Effects of waterlogging on the gas exchange,chlorophyll fluorescence and water potential ofQuercusvariabilisandPterocaryastenoptera[J]. J Plant Ecol, 30(6):960-968. [衣英华,樊大勇,谢宗强,等, 2006. 模拟淹水对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响 [J]. 植物生态学报, 30(6):960-968.]
YU SW, TANG ZC, 1998. Plant physiology and molecular biology [M]. Beijing: Science Press. [余叔文, 汤章城, 1998. 植物生理与分子生物学 [M]. 北京: 科学出版社.]
ZHANG BB,MA DJ,CAI ZX, et al, 2013. Photosynthetic characteristics response to water-logging in 3 peach rootstock seedlings [J]. Acta Bot Boreal-Occident Sin, 33(1): 146-153. [张斌斌,马瑞娟,蔡志翔,等, 2013. 3个桃砧木品种对淹水的光合生理响应特征 [J]. 西北植物学报, 33(1): 146-153.]
ZOU Q, 2000. Plant physiology experimental guidance [M]. Beijing: China Agriculture Press. [邹琦, 2000. 植物生理学实验指导 [M]. 北京: 中国农业出版社.]
Effects of flooding stress on growth and photosynthesis ofPaeoniasuffruticosa
ZHU Xiang-Tao, JIN Song-Heng*, AI Jian-Guo, SHI Hao-Jie
(ZhejiangAgriculturalandForestryUniversity, Lin’an 311300, China )
Growth and photosynthesis of three-year-old seedlings ofPaeoniasuffruticosa“fengdanbai” were investigated in a 30-day flooding experiment. Three treatment levels were set in this experiment: normal management, mild stress and severe stress levels. The results showed that after 30 d of the above three treatments accordingly, the height growth ofP.suffruticosawas 3.6, 1.1 and 0.73 cm, the ground diameter growth ofP.suffruticosawas 0.21, 0.11 and 0.06 cm. The increase in total plant biomass was 7.0, 3.0 and 2.75 g. The difference was remarkable between flooding stress and normal growth, the growth ofP.suffruticosawas seriously affected by flooding stress. At the same time, total chlorophyll content increased under normal management, but declined under stress conditions. The root activity decreased when treated with flooding at different times. Meanwhile, under flooding stress, the transpiration rate and stomatal conductance ofP.suffruticosaalso decreased. Under mild flooding stress, the intercellular carbon dioxide concentration first increased and then decreased, but increased under severe flooding stress. The study suggests that flooding stress affects theP.suffruticosaroot vigor, stem, leaf growth and photosynthetic characteristics. This study will provide the information for stress mechanism of jiangnanP.suffruticosa.
Paeoniasuffruticosa, flooding stress, root activity, growth, photosynthesis
10.11931/guihaia.gxzw201501032朱向涛, 金松恒, 哀建国, 等. 淹水胁迫下江南牡丹生长及光合特性研究 [J]. 广西植物, 2016, 36(8):956-962
ZHU XT, JIN SH, AI JG, et al. Effects of flooding stress on growth and photosynthesis ofPaeoniasuffruticosa[J]. Guihaia, 2016, 36(8):956-962
2015-01-22
2015-04-17
国家自然科学基金(31170584); 浙江省教育厅项目(Y201223938); 浙江农林大学博士科研启动基金(2011FR017) [Supported by the National Natural Science Foundation of China (31170584); Zhejiang Program of Educational Office (Y201223938); Start-up Program for Doctors Research of Zhejiang Agricultural and Forestry University (2011FR017)]。
朱向涛(1982-),山东莱芜人,博士,讲师,主要从事园林植物应用研究,(E-mail)zxt8202@163.com。
金松恒,博士,教授,研究方向为植物生理生态,(E-mail)shjin@zafu.edu.cn。
Q945
A
1000-3142(2016)08-0956-07
