栗斑腹鹀鸣声质量与繁殖投入关系研究
2016-09-22刘丽秋张立世陈艳东陈乐勇姜云垒
刘丽秋,张立世,李 时,陈艳东,陈乐勇,姜云垒
(吉林农业大学动物科学技术学院,吉林 长春 130118)
栗斑腹鹀鸣声质量与繁殖投入关系研究
刘丽秋,张立世,李时,陈艳东,陈乐勇,姜云垒
(吉林农业大学动物科学技术学院,吉林 长春 130118)
利用偏相关分析方法,分析了雄性栗斑腹鹀的鸣声质量与繁殖投入的关系.结果表明:雄性鸣唱率与首枚卵产期呈显著负相关(Pearson偏相关分析:r=-0.421,df=27,P=0.023);句子长度与雏鸟数目呈显著正相关(Pearson偏相关分析:r=0.598,df=16,P= 0.024).雄性递食率与鸣唱特征无显著相关性.说明雌性栗斑腹鹀可能根据雄性鸣唱率评价雄性个体质量和抚育后代的能力.
栗斑腹鹀;鸣唱;繁殖投入
鸟类鸣唱具有吸引配偶和保卫领域的功能,在繁殖过程中起重要作用[1].鸟类鸣唱是由简单到复杂的进化过程,而繁殖期的鸣唱最为精致、复杂、多变,这可能是因为性选择压力影响了雄性鸣声的进化[2].研究表明雌性一般偏爱于那些善于鸣唱的雄性;同时雄性的鸣唱质量也影响雌性的繁殖行为,如雄性鸣唱越频繁、句子越多样化,雌性就越早开始孵卵[3-4].
雌性配偶选择主要依据雄性参与繁殖的能力,包括雄性身体质量、雄性为后代提供食物的能力等[5].雌性会根据羽毛特征和其他一些视觉信号对雄性的领域质量进行评价,还可能利用雄性鸣唱的质量和数量特征来评价父本质量.假设鸣唱对个体损耗很大[6],那么鸣唱的数量和复杂性可能是评价雄性质量极为重要的特征.关于鸟类鸣唱的研究表明:雌性更喜欢大曲目和长句子的鸣唱[7-10].雄性鸣唱复杂性与个体质量之间的关系研究表明:雄性的鸣唱特征能真实反应个体的质量,因为鸣唱学习的时间和大脑结构调节学习的时间是在鸟类的早期发展阶段,而这一阶段正是鸟类最可能面临食物压力和其他压力的阶段[11].
栗斑腹鹀(Emberizajankowskii)隶属于雀形目(Embrizidae)鹀科(Emberiza)鹀属(Emberiza),无亚种分化,属单型种,为我国特有种[12-13].近年来,由于栗斑腹鹀种群数量急剧下降,现被世界自然保护联盟(IUCN)列为濒危种,保护级别为濒危鸟类[14].了解栗斑腹鹀在面临生境变化、人为干扰、捕食等压力下的繁殖情况和该物种的行为对该物种保护非常重要.目前,国内已对栗斑腹鹀的分布和繁殖习性[15]、繁殖生态[16-17]以及栗斑腹鹀种群数量变化[18]等进行了研究,但对栗斑腹鹀鸣声的研究尚未见报道.栗斑腹鹀属于单配制且雄性参与后代抚育,雄性鸣声质量是否影响其自身和雌性的繁殖投入尚不清楚,为此本文对雄性栗斑腹鹀鸣声特征与双亲繁殖投入之间的关系进行了研究.
1 研究区域概况
研究区域位于内蒙古通辽市扎鲁特旗(119°34′48″~121°56′50″E,43°50′13″~45°35′31″N),海拔179.2~1 444.2 m,属于中纬度温带大陆性气候,地貌特征以低山丘陵和倾斜冲积平原为主.主要植被有芨芨草(Achnatherumsplendens)、羊草(Leymuschinensis)、贝加尔针茅(StipabaicalensisRoshev)等禾本科植物,线叶蒿(Artemisiasubulata)、线叶菊(Filifoliumsibiricum)、麻花头(Serratulacentauroides)等菊科植物,以及西伯利亚山杏(Prunussibirica)、蒙古黄榆(Ulmusmacrocarpa)两种木本植物.
2 研究方法
2.1鸣声采集
2014—2015年5—7月,在研究地区内利用直接搜寻法确定栗斑腹鹀的繁殖巢,用GPS对巢进行定位并做好标记,隔天观察巢内的变化情况.利用TASCAM HD-P2 便携式数字录音机(日本)和Sennheiser MKH416 P48 外接强指向性话筒(德国),对栗斑腹鹀孵化期鸣声进行录制.录制时间在每日6:00—11:00,录音距离一般距巢小于30 m.共录制了47巢雄性栗斑腹鹀个体的鸣唱.录音参数设置如下:编码格式(Encoder)WAV,二进制(Bit Depth)16,采样频率(Sample Rate)44.1 Hz,通道(Channels)Stereo,输入状态(Input Level)L.
2.2繁殖投入
记录首枚卵产期、窝卵数、孵化期、成功出飞的雏鸟数量等繁殖指标,并采集这些巢雄体的鸣唱.雏鸟随着日龄的增加对食物的需求量也随之增加[19],因此在对亲鸟递食频率进行观察时,将育雏期划分为两个阶段:第一阶段为雏鸟2~3 d;第二阶段为雏鸟6~7 d.利用在巢上方架设的微型摄像头并在距巢约30 m处用望远镜录制、观察记录繁殖投入情况,录制和观察的时间为6:00—18:00.共录制了13巢栗斑腹鹀育雏期的视频.
3 数据处理及统计分析
应用Avisoft SAS Lab Pro 5.2鸣声处理软件对栗斑腹鹀鸣声进行声学特征参数采集,随机选取每只雄性个体的10个鸣唱句子进行鸣声参数采集分析.软件语图参数设置为:快速傅旦叶变换(FFT length)256,帧长(Frame size)100%,窗口(Window)Hamming,频率分辨率(Frequency resolution)172 Hz,重叠(Overlap)50%,时间分辨率(Time resolution)2.9 ms.音节的鉴定和分类通过目测和比较声谱图,鸣声分析包括以下变量:
鸣唱句子的多功能性:用鸣唱句子中音节的类型数与同一句子音节总数的比值作为评估句子复杂性的指数,其中音节数不包括起始音节.参考Tracy 和Baker(1999)[20]的方法对音节进行划分:相邻音素间隔在0.1 s以上的视作不同音节,时间间隔小于0.02 s的则视为一个音节.而时间间隔在0.02~0.1 s 之间的音素,依以下情况而定:假如音素复合体中的每个音素都没有在其他鸣唱中单独出现过,那么这一音素复合体应被视为一个音节;反之,这些音素应被视作不同音节[21].
鸣唱的多功能性:样本中10个句子的音节类型数与同一样本音节总数的比值.句子长度:鸣唱句子中从开始的第一个音节到最后一个音节的持续时间.频率范围:声谱图上最大频率和最小频率的差值(四舍五入到0.01 kHz).鸣唱率:鸣唱句子数量与记录时间的比值[22].
采用SPSS21.0软件进行统计分析,通过单样本Kolmogorov-Smirnov检验判定各组数据是否符合正态分布,利用单样本t检验验证不同巢之间雄鸟的句子多功能性、鸣唱多功能性、句子长度、频率范围、鸣唱率是否存在显著差异.如果存在显著差异符合正态分布则选择双尾Pearson相关分析方法进行分析,否则采用Spearman相关分析方法.考虑到鸣声参数之间的相互影响,利用偏相关分析方法对鸣声参数与繁殖投入的关系进行分析.
4 结果
4.1鸣唱特征
随机选取每巢雄性10个清晰的语句进行鸣声参数采集,得到的栗斑腹鹀鸣声特征参数见表1.鸣声参数显示了个体之间的变化(变异系数>20%),其中句子长度显示了相对较大的变化,句子多功能性的变化相对较小.
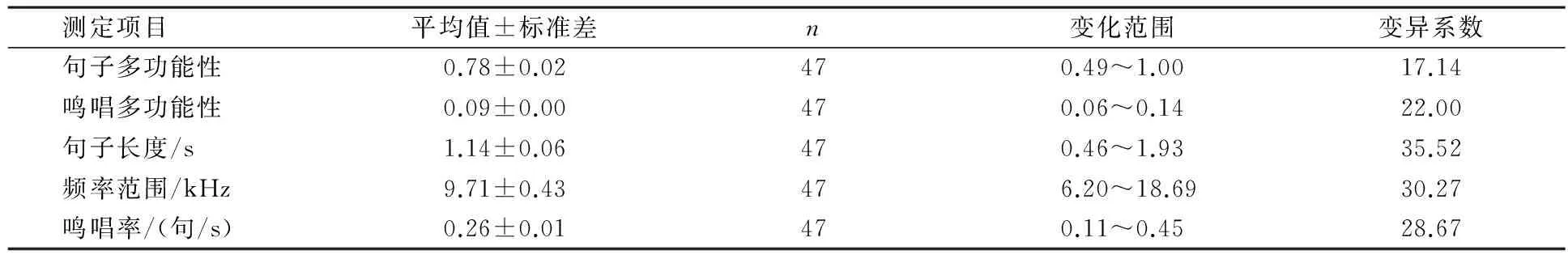
表1 栗斑腹鹀鸣唱特征
4.2鸣唱特征与首枚卵产期的关系
雄性的鸣唱率与首枚卵产期呈显著负相关(Pearson偏相关分析:r=-0.421,df=27,P=0.023)(见图1).首枚卵产期与句子多功能性(Pearson偏相关分析:r=0.018,df=27,P=0.928)、鸣唱多功能性(Pearson偏相关分析:r=-0.087,df=27,P=0.654)、句子长度(Pearson偏相关分析:r=0.158,df=27,P=0.412)、频率范围(Pearson偏相关分析:r=-0.061,df=27,P=0.753)之间没有显著的相关性.
4.3鸣唱特征与亲鸟投入的关系
育雏期第一阶段雄性的递食率为1.36次/h,第二阶段的递食率为1.67次/h,两阶段合并雄性递食率为1.46次/h;育雏期第一阶段雌性的递食率为1.53次/h,第二阶段的递食率为1.99次/h,两阶段合并雌性递食率为1.66次/h(见表2).双亲递食率呈极显著正相关(Pearson偏相关分析:r=0.874,n=13,P<0.001),雄性递食率与鸣唱特征无显著的相关性(见表3).
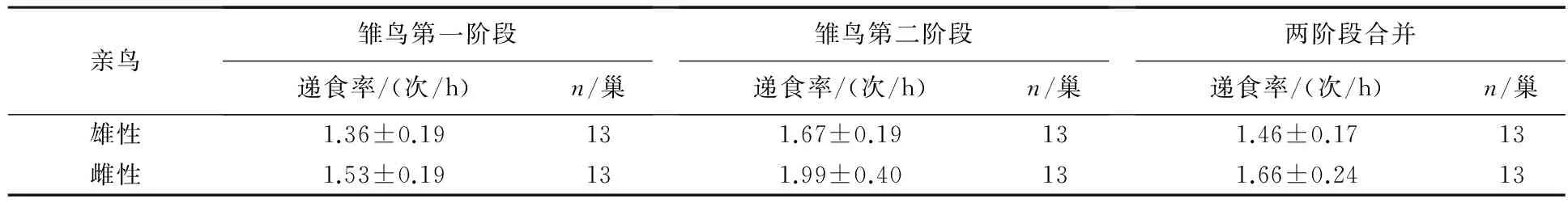
表2 栗斑腹鹀雏鸟阶段递食率
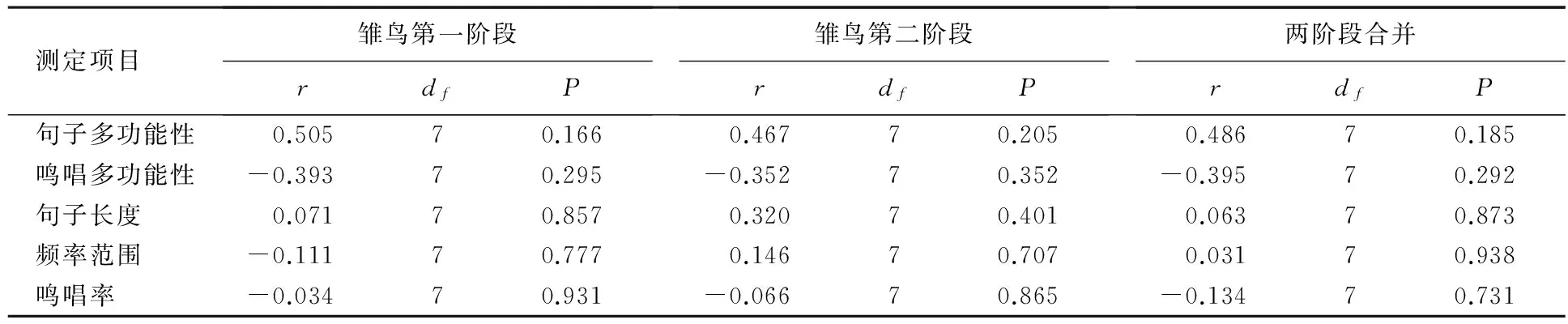
表3 鸣唱特征与雄性递食率之间的相关分析
4.4鸣唱特征与雏鸟数目的关系
雏鸟数目与句子长度呈显著正相关(Pearson偏相关分析:r=0.598,df=16,P=0.024)(见图2).雏鸟数目与句子多功能性(Pearson偏相关分析:r=0.410,df=16,P=0.146)、鸣唱多功能性(Pearson
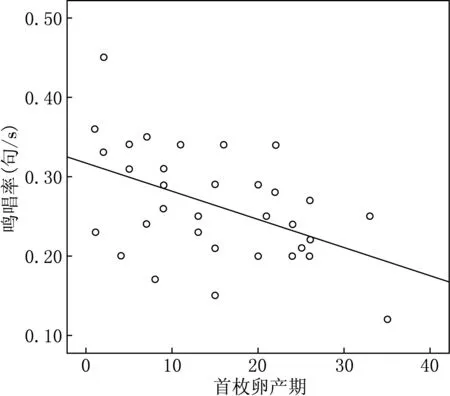
0:5月17日,10:5月27日,以此类推图1 栗斑腹鹀首枚卵产期与雄性鸣唱率之间的关系
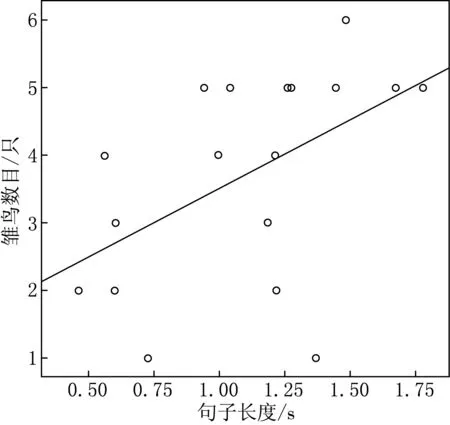
图2 栗斑腹鹀句子长度与雏鸟数目之间的关系
偏相关分析:r=-0.237,df=16,P=0.415)、频率范围(Pearson偏相关分析:r=-0.438,df=16,P=0.117)、鸣唱率(Pearson偏相关分析:r=0.037,df=16,P=0.899)之间没有显著的相关性.
5 讨论
5.1鸣唱特征与首枚卵产期的关系
研究中发现,鸣唱率越高雄性越早开始繁殖,而鸣唱复杂性与首枚卵产期没有相关,表明了雌体在配偶选择时对鸣唱率的偏爱.可能原因的是:鸣唱率的高低反映了雄性的领域质量及身体质量[23],雄性通过高的鸣唱率来提高自身吸引异性的能力,进而能够更快地吸引雌性并与之交配繁殖.Yasukawa(1981)[24]的研究发现,鸣唱率较高的雄性在寻找食物和领域保护方面比其他的雄性更有能力;Gottlander(1987)[25]的研究指出,鸣唱率会反映雄性的领域质量,雌性可能会更倾向于选择高鸣唱率的雄性进行交配,高鸣唱率的雄性能够为后代提供充足的食物,这对雌性是非常重要的.也有研究表明,石雀的鸣唱率能够反映其年龄和繁殖成功率,年轻的石雀繁殖成功率较低,他们会通过增加鸣唱率来提升社会地位,而年长的石雀鸣唱率低繁殖成功率高.
5.2鸣唱特征与亲鸟投入的关系
本研究没有发现任何鸣唱特性与雄性提供的亲代抚育(喂养雏鸟)能力之间有相关性,而与双亲递食率呈极显著正相关.在雏鸟期的第一阶段,雌性会长时间在巢中暖雏或孵化未出壳的卵,所以雄性较低的递食率是因为雌性递食率比较低,而两性较低的递食率可能与雏鸟较低的食物需求有关.但是对雪鹀的研究发现,鸣唱率与雄性递食率呈正相关[22].在雏鸟期的第二阶段雌性不再暖雏,因此对雌性来说评价雄性是否愿意并且有能力参与雏鸟的喂养是很重要的.研究表明:雄性夜莺的鸣唱特征代表其身体的质量,夜莺的鸣唱复杂性和句子排序与雄性递食率有关;此外鸣唱特征能够预测雄性抚育后代的能力[26].本研究结果表明,栗斑腹鹀的鸣唱特征不能反映亲代抚育的能力,所以对雌性来说评价雄性是否愿意并且有能力参与雏鸟的喂养是很重要的.
5.3鸣唱特征与雏鸟数目的关系
双亲成功抚育后代的数目很大程度上取决于雌性评价雄性质量的能力,而鸣唱特性为这一评价提供了重要线索.在本研究中我们发现,句子长度与雏鸟数目呈显著正相关,其他大部分鸣唱特征与雏鸟数目无关.这可能是因为较长句子的鸣唱可以吸引雌性并促使雌性更多的投入,而长句子的鸣唱可能也反映出雄性具有优良的遗传基因,使得后代有较高的存活率.有研究表明,斑胸草雀的鸣声质量能够预测其繁殖成功率[27];Catchpole等[28]发现鸣唱复杂性与雏鸟数目呈显著正相关;Mcgregor等对大山雀的研究发现,鸣唱复杂性与繁殖投入之间有相关性,鸣唱复杂的雄性繁殖成功率会更高一些,但是鸣唱复杂性的高低是有差异的,这可能是因为采用了不同方法评估和测量的结果.
[1]COLLINS S. Vocal fighting and flirting:the functions of birdsong[G]// MARLER P,SLABBEKOORN H. Nature’s music. Oxford:Academic Press,2005:39-79.
[2]HAMILTON W D,ZUK M. Heritable true fitness and bright birds:a role for parasites[J] Science,1982,218:384-387.
[3]CATCHPOLE C K,SLATER P J B. Bird song:biological themes and variations[M]. Cambirdge:Cambridge University Press,1995.
[4]WRIGHTJ,CUTHILL I. Monogamy in the European starling[J]. Behaviour,1992,120:262-285.
[5]LYON B E,MONTGOMERIE R D. Conspicuous plumage of birds:sexual selection or unprofitable prey[J]. Animal Behaviour,1985,33(3):1038-1040.
[6]HORN A G,LEONARD M L,WEARY D M. Oxygen consumption during crowing by roosters:talk is cheap[J]. Animal Behaviour,1995,50(5):1171-1175.
[7]CATCHPOLE C K. Sexual and the evolution of complex songs among European Warblers of the Genus Acr Ocephal Us[J]. Behaviour,1980,74(1):149-165.
[8]EENS M,PINXTEN R,VERHEVEN R F. Male song as a cue for mate choice in the European starling[J]. Behaviour,1991,116(3):210-238.
[9]KREUTZER M,VALLET E,NAGLE L. Sexual responsiveness of female canaries to song bout organization[J]. Behaviour,1994,129(3):293-305.
[10]LAMPE H M,SAETRE G P. Female pied flycatchers prefer males with larger song repertoires[J]. Proceedings of the Royal Society of London B:Biological Sciences,1995,262(1364):163-167.
[11]NOWICKI S,HASSELQUIST D,BENSCH S,et al. Nestling growth and song repertoire size in great reed warblers:evidence for song learning as an indicator mechanism in mate choice[J]. Proceeding of the Royal Society of London B:Biological Sciences,2000,267(1460):2419-2424.
[12]WANG S,ZHENG G M,WANG Q S. China red data book of endangered animals(Aves)[M]. Beijing,Hong Kong,New York:Science Press,1998:338-339.
[13]雷富民,卢汰春. 中国鸟类特有种[M]. 北京:科学出版社,2006.
[14]BIRD LIFE INTERNATIONAL. Species factsheet:Emberizajankowskii[EB/OL].[2013-04-16].http://www.Birdlife. org. Accessed 3 November 2010.
[15]傅桐生,陈鹏. 栗斑腹鹀的分布及其繁殖习性[J]. 动物学报,1966,18(2):195-198.
[16]高玮,王海涛,孙丹婷. 栗斑腹的栖息地和巢址选择[J]. 生态学报,2003,23(4):665-672.
[17]佟富春,肖以华,白哈斯,等. 吉林省白城地区干草原栗斑腹鹀的繁殖生态[J]. 生态学报,2002,22(9):1485-1490.
[18]程瑾瑞,高玮. 栗斑腹鹀种群数量变化的分析[J]. 东北师大学报(自然科学版),2002,34(1):49-53.
[19]陶慧娟,姜云垒,秦博,等. 栗斑腹鹀双亲递食率比较研究[J]. 东北师大学报(自然科学版),2014,46(3):105-108
[20]SZIJJ L J. Hybridization and the nature of the isolating mechanism in sympat ric populations of meadowlarks(Sturnella) in Ontario. Z [J]. Tierpsychol,1966,23:677-690.
[21]BAKER M C,HOWARD T M,SWEET P W. Microgeographic variation and sharing of the gargle vocalization and its component syllables in Black-capped Chickadee(Aves,Paridae,Poecileatricapillus) populations[J]. Ethology,2000,106:819-838.
[22]HOFSTAD E,ESPMARK Y,MOKSNES A,et al. The relationship between song performance and male quality in snow buntings(Plectrophenaxnivalis)[J]. Canadian Journal of Zoology,2002,80(3):524-531.
[23]LAMPE H M,ESPMARK Y O. Mate choice in Pied Flycatchers Ficedula hypoleuca:can females use song to find high-quality males and terriories[J]. Ibis,2003,145(1):E24-E33.
[24]YASUKAWA K. Male quality and female choice of mate in the red-winged blackbird(Agelaiusphoeniceus)[J]. Ecology,1981:922-929.
[25]GOTTLANDER K. Variation in the song rate of the male pied flycatcher Ficedula hypoleuca:causes and consequences[J]. Animal Behaviour,1987,35(4):1037-1043.
[26]BARTSCH C,WEISS M,KIPPER S. Multiple song features are related to paternal effort in common nightingales[J]. BMC Evolutionary Biology,2015,15(1):1.
[27]WOODGATE J L,MARIETTE M M,BENNETT A T D,et al. Male song structure predicts reproductive success in a wild zebra finch population[J]. Animal Behaviour,2012,83(3):773-781.
[28]CATCHPOLE C,LEISLER B,WINKLER H. Polygyny in the great reed warbler,Acrocephalus arundinaceus:a possible case of deception[J]. Behavioral Ecology and Sociobiology,1985,16(3):285-291.
(责任编辑:方林)
The relationship between song performance and reproductive output in Jankowski’s Bunting(Emberizajankowskii)
LIU Li-qiu,ZHANG Li-shi,LI Shi,CHEN Yan-dong,CHEN Yue-yong,JIANG Yun-lei
(School of Animal Science and Technology,Jilin Agricultural University,Changchun 130118,China)
Attracting females is considered to be one of the main functions of bird song. Both the rate and complexity of male song are assumed to be reliable cues,that may be used by the female when choosing a mate. A statistically significant negative correlation was found between song rate and the date for the first egg laid in the clutch(Pearson partial correlation analysis:r=-0.421,df=27,P=0.023) as well a positive correlation between the length of the song strophes and the number of fledglings(Pearson partial correlation analysis:r=0.598,df=16,P=0.024). But no statistically significant relation was found between any of the song parameters and the male’s feeding rate. These results suggest that females might use male song rate to assess male quality and ability to participate in raising chicks.
Jankowski’s Bunting;song;reproductive output
1000-1832(2016)03-0110-05
2016-01-23
国家自然科学基金资助项目(31172109);吉林省自然科学基金资助项目(20150101067JC).
刘丽秋(1989—),女,硕士研究生;通讯作者:姜云垒(1962—),男,博士,教授,博士研究生导师,主要从事鸟类行为生态学研究.
Q 959.7+39[学科代码]180·5744
A
[DOI]10.16163/j.cnki.22-1123/n.2016.03.021
