小麦-滨麦附加易位系DM 5911的创制及其细胞遗传学鉴定
2016-09-21程雪妮庞玉辉雷忠萍陈新宏杨群慧赵继新
程雪妮,刘 洋,庞玉辉,3,雷忠萍,武 军,陈新宏,杨群慧,赵继新
(1.西北农林科技大学生命学院,陕西杨凌 712100; 2.西北农林科技大学农学院/陕西省植物遗传工程育种重点实验室,陕西杨凌 712100; 3.河南科技大学农学院,河南洛阳 471023)
小麦-滨麦附加易位系DM 5911的创制及其细胞遗传学鉴定
程雪妮1,刘 洋2,庞玉辉2,3,雷忠萍1,武 军2,陈新宏2,杨群慧2,赵继新2
(1.西北农林科技大学生命学院,陕西杨凌 712100; 2.西北农林科技大学农学院/陕西省植物遗传工程育种重点实验室,陕西杨凌 712100; 3.河南科技大学农学院,河南洛阳 471023)
滨麦(Leymusmollis(Trin.) Pilger,NsNsXmXm,2n=28)具有抗旱、耐寒、抗多种真菌和细菌病害、茎秆粗壮、大穗多花等特性,是麦类作物品种改良的优异种质资源。本研究采用细胞遗传学方法对由八倍体小滨麦M842-16与硬粒小麦品种D 4286杂交F6代选育出的1个附加易位系DM 5911进行了鉴定和农艺性状分析。细胞学镜检显示,DM 5911的染色体构型为2n=44=22Ⅱ;根尖体细胞原位杂交鉴定显示,DM 5911含有2条完整的Ns染色体和2条易位的Ns染色体;花粉母细胞原位杂交鉴定显示,DM 5911的2条完整Ns染色体和2条易位Ns染色体可以进行正常的联会配对和遗传。表明DM 5911是1个遗传稳定、包含1对完整的Ns染色体和1对易位Ns染色体的小滨麦附加易位系。形态学调查显示,DM 5911在株高和分蘖数等农艺性状方面得到了明显的改善。该附加易位系作为进一步创制小滨麦小片段染色体易位系的重要种质材料,可用于小麦染色体工程育种与遗传改良。
小麦; 滨麦; 附加; 易位; 基因组原位杂交
小麦远缘杂交是研究小麦进化及亲缘关系、人工合成新物种、利用外源遗传物质丰富小麦遗传基础的重要途径和手段[1-2]。滨麦(Leymusmollis(Trin.) Pilger,NsNsXmXm,2n=28),又名柔软赖草,属禾本科大麦亚族赖草属野生杂草,具有抗旱、耐寒、耐盐碱、耐瘠薄、抗多种真菌与细菌病害、茎秆粗壮、大穗多花等特性,是麦类作物品种改良的优异种质资源[3-4]。滨麦基因组为NsNsXmXm,其中Ns基因组来自于新麦草(PsathyrostachysKeng),而Xm基因组的来源目前尚未确定[5-6]。
20世纪50-60年代,前苏联的Tsitsin等[7]通过幼胚培养,获得了四倍体、六倍体小麦与大赖草、滨麦和沙生赖草的杂种。Anamthawat-Jónsson 等[8-9]采用胚拯救和秋水仙碱加倍,获得了普通小麦、波斯小麦与滨麦的杂种双二倍体,并报道了1个包含全部A和B基因组染色体、1对D基因组染色体和6对滨麦染色体的小麦-滨麦双二倍体材料AD 99,利用AD 99与普通小麦杂交,筛选出了6个抗白粉病的纯合株系。陈漱阳等[10]、傅 杰等[11]自20世纪80-90年代开始进行普通小麦与滨麦的杂交研究,通过胚拯救和秋水仙碱染色体加倍,获得了具有滨麦的大穗、多花、大粒、抗病等特性的八倍体小滨麦(octoploidTritileymus) M 842。王献平等[12]对6 种类型的八倍体小滨麦的染色体组构成进行了分子细胞遗传学分析,将八倍体小滨麦M 842-4、M 842-8、M 842-12、M 842-16 的染色体组构成确定为AABBDDNN,M 842-10 的染色体组构成确定为AABBDDNN+2Ta-2Lm(N),M 842-13 的染色体组构成确定为AABBDDJJ。利用八倍体小滨麦M 842与普通小麦品种7182、烟农15和缺体小麦等杂交,前人已培育出3个异附加系[13]、2个二体异代换系[14]和10多个易位系[15-18](M 853、M 8657、WM-10、93748、山农0096、山农6343、WL 24-4等)新种质。此外,M 842与八倍体小偃麦、八倍体小簇麦和八倍体小黑麦杂交,获得了一些小麦-滨麦-偃麦草[19]、小麦-滨麦-簇毛麦[20]和小麦-黑麦-滨麦[21]等三属杂种,并将滨麦的抗条锈病、抗白粉病等基因导入到了普通小麦中。
近年来,本课题组利用八倍体小滨麦M 842与硬粒小麦品种D 4286和Trs-372进行了杂交研究[22],并在后代中鉴定出了一些新种质。在八倍体小滨麦M 842-12与硬粒小麦Trs-372的杂交后代中,鉴定出1个小麦1D、5D、6D染色体被滨麦1Ns、5Ns、6Ns染色体替换了的多重异代换系05DM6[23];在八倍体小滨麦M 842-16与硬粒小麦D 4286的杂交后代中,鉴定出1个小麦3D、6D、7D染色体被滨麦3Ns、6Ns、7Ns染色体替换了的多重异代换系10DM50[24]和1个小麦3D染色体被滨麦3Ns染色体替换了的二体异代换系10DM57[25]。
本研究在M 842-16×D 4286的杂交后代中,筛选出了1个染色体数目为2n=44,含有2条完整滨麦Ns染色体和2条易位Ns染色体的株系DM 5911,对其进行细胞遗传学和基因组原位杂交鉴定,并对其农艺性状进行调查,以期为小麦品种改良和染色体工程育种提供新的种质资源。
1 材料与方法
1.1供试材料
小滨麦附加易位系DM 5911,来自M 842-16×D 4286的F6世代;八倍体小滨麦M 842-16是普通小麦7182(Triticumaestivumcv. 7182,AABBDD,2n=42)与滨麦(Leymusmollis)杂交F1代经胚拯救而获得的双二倍体,其染色体组成为AABBDDNsNs,2n=56;华山新麦草(Psathyrostachyshuashanica,NsNs,2n=14)作为基因组原位杂交鉴定时Ns基因组特异探针,用来检测杂交后代中的滨麦Ns染色体;硬粒小麦品种D 4286(Triticumdurumcv. D 4286,AABB,2n=28)和普通小麦品种7182为农艺性状调查时的参考对照;这些材料均保存于西北农林科技大学农学院。
1.2试验方法
1.2.1细胞遗传学分析
小麦根尖体细胞观察:小麦种子于室内发芽,待根长至1~2 cm 时,剪取根尖于0~4 ℃冰水预处理24 h,卡诺固定液(无水乙醇∶冰乙酸= 3∶1) 固定,70%乙醇保存,醋酸洋红染色压片;Olympus BX60 显微镜观察并照相。
花粉母细胞观察:田间取适龄幼穗,取花粉用卡诺固定液(无水乙醇∶氯仿∶冰乙酸= 6∶3∶1)固定,醋酸洋红染色压片;Olympus BX60 显微镜观察并照相。
1.2.2基因组原位杂交(Genomicinsituhybridization,GISH)
DNA提取及探针标记:采用CTAB法[26]提取滨麦和华山新麦草全基因组DNA。依照Dig-Nick-Translation Mix试剂盒(Roche,Germany)使用说明对华山新麦草的全基因组DNA进行标记。
染色体制片及原位杂交:参照文献[12]与[22]的方法进行。每张制片加40 μL杂交液,包含20×SSC 4 μL,ssDNA(鲑鱼精DNA,5 μg·μL-1) 1 μL,10%(W/V)SDS(十二烷基磺酸钠) 1 μL,50%(W/V)DS(硫酸葡聚糖) 8 μL,去离子甲酰胺20 μL,探针DNA 100 ng,无菌去离子水加至40 μL。95 ℃变性8 min,杂交在hybrite原位杂交仪(Thermo,USA)上依照程序(75 ℃ 8 min,37 ℃ 16 h)进行。Olympus BX60荧光显微镜观察,Pixera Penguin 150CL显微数码CCD照相。
1.2.3农艺性状调查
供试材料于2013-2015年度先在实验室发芽后,将幼苗移栽于发芽盘,待幼苗长至一叶一心时,移栽至西北农林科技大学小麦试验地。在成熟期调查供试材料的株高、分蘖数、穗长、小穗数、穗下茎节长度、结实率等性状。均随机调查10个样本,取平均值。采用t测验方法对DM 5911和普通小麦亲本7182在各性状上的差异进行显著性分析。
2 结果与分析
2.1DM 5911根尖体细胞染色体的鉴定结果
利用M842-16(AABBDDNN,2n=56)与硬粒小麦品种D 4286(AABB,2n=28)杂交,后代经套袋自交,在F6世代中,通过根尖体细胞染色体镜检,筛选到1株2n=44的植株,收获该单株。随机选取15粒该单株种子进行细胞学镜检,结果表明,15粒种子的根尖体细胞染色体数目都为44条(图1A)。随机选取该株系6个单株,用华山新麦草基因组DNA为探针进行GISH鉴定,结果(图1B)显示,这6个单株除了2条完整的染色体显示黄绿色荧光信号外,还有2条染色体部分显示黄绿色荧光信号,由此确定该株系含有2条完整的Ns染色体和2条易位的Ns染色体,将该株系命名为DM 5911。
2.2DM 5911花粉母细胞染色体鉴定结果
田间选取DM 5911的适龄幼穗,对45个花粉母细胞减数分裂中期Ⅰ染色体配对情况进行镜检,结果显示,DM 5911有38个花粉母细胞的染色体构型为2n=22Ⅱ,占观察细胞数的84.5%;在二价体中,环状二价体占有很高的比例,达到82.6%,平均每个细胞有18.0个,而棒状二价体仅占17.4%,平均每个细胞有3.8个(图2A,表1)。以华山新麦草总基因组DNA作为探针,对其花粉母细胞进行基因组原位杂交鉴定,结果显示,DM 5911花粉母细胞减数分裂中期Ⅰ细胞除了含有1个完整的二价体显示黄绿色信号外,还有1个棒状二价体在两端显示黄绿色信号(图2B),在减数分裂后期Ⅰ,伴随着细胞分裂,完整的Ns二价体和易位的Ns二价体均正常的分离为2条染色单体向细胞两极移动(图2C)。表明DM 5911中的2条完整的Ns染色体和2条易位的Ns染色体都进行了正常的联会配对和遗传。由此说明,DM 5911是1个遗传稳定、包含1对完整的Ns染色体和1对易位Ns染色体的小滨麦附加易位系。
2.3DM 5911的形态学特征
对DM 5911及其亲本硬粒小麦品种D 4286和八倍体小滨麦M 842-16的形态学性状进行了调查,结果(图3)显示,DM 5911株高明显变矮,平均株高为65.9 cm,比双亲都低(图3A);穗形与亲本M 842-16较为接近,都为纺锤形,芒的长度明显变短,与亲本M 842-16较为接近(图3B)。DM 5911的分蘖数、穗长、小穗数以及结实率均介于双亲之间,而株高和穗下茎节长度均明显低于双亲。方差分析表明,DM 5911与亲本M 842-16、D 4286、7182相比,分蘖数和穗下茎节长度差异显著,株高、小穗数、穗粒数和结实率等性状差异极显著,穗长差异不显著(表2)。
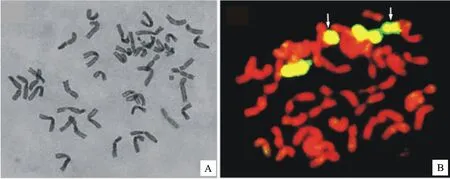
箭头指示小麦-滨麦Ns易位染色体。
Arrows show two chromosomes translocated between wheat and Ns chromosomes.
图1DM 5911的根尖体细胞染色体(A)及其基因组原位杂交鉴定(B)
Fig.1 Somatic chromosomes in the root tips(A) and its identification by GISH(B) in DM 5911

图B中,箭头指示Ns易位二价体;图C中,白色箭头指示Ns易位染色体,蓝色箭头指示Ns完整染色体。
Arrows show translocated Ns bivalent in Fig.B;Blue and white arrows in Fig.C show whole Ns chromosomes and translocation Ns chromosomes,respectively.
图2DM 5911花粉母细胞减数分裂中期Ⅰ染色体(A)及其减数分裂中期Ⅰ和后期 Ⅰ基因组原位杂交鉴定(B和C)
Fig.2Chromosome behavior in PMCs at metaphase Ⅰ(A) and its GISH identification at metaphase Ⅰ and anaphase Ⅰ(B and C) in PMCs in DM 5911

材料Material观察细胞数No.ofcellsobserved2n染色体构型Chromosomeconfiguration22Ⅱ21Ⅱ+2Ⅰ20Ⅱ+4Ⅰ染色体配对ChromosomepairⅠⅡ总数Total环状Rings棒状RodsDM5911454438(84.5%)5(11.1%)2(4.4%)0.4(0~4)21.8(20~22)3.8(0~7)18.0(15~21)
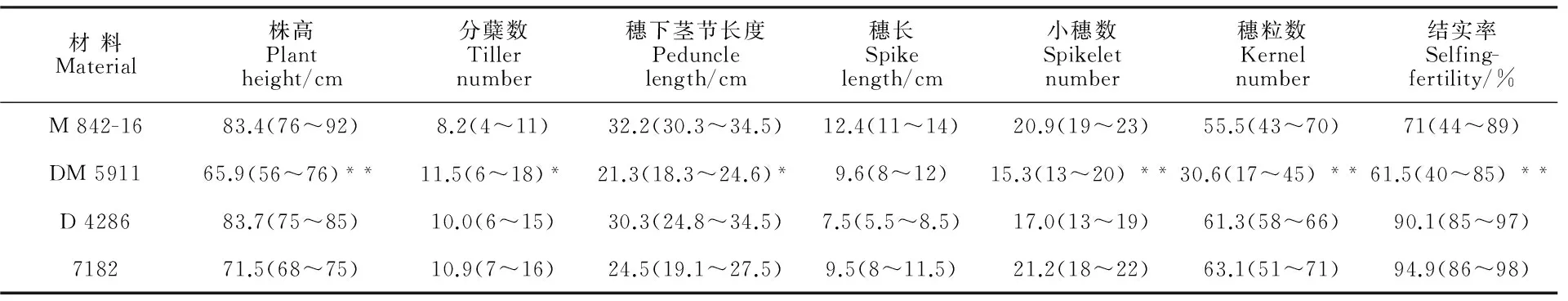
表2 DM 5911及其亲本的形态学特征
括号内数据为性状变异范围;*和**分别表示在P<0.05 和P<0.01水平上与其他3个材料差异显著。
Agronomic trait variation range are shown in brackets; * and ** indicate significant differences with other three materials atP<0.05 andP<0.01,respectively.
3 讨 论
自20世纪50-60年代以来,前人通过胚拯救和秋水仙碱染色体加倍等技术,先后获得了许多六倍体和八倍体小麦-滨麦杂种[3,7-8,11]。以八倍体小滨麦M 842为亲本,国内研究者创制出了一系列异附加系、异代换系和易位系等小滨麦衍生后代材料。傅 杰等[13-14]通过M842与普通小麦和缺体小麦杂交,培育出了3个异附加系和2个异代换系。周兖晨等[18]在M 842与普通小麦品种7182的杂交后代中鉴定出了抗条锈病的小滨麦易位系93748。王献平等[17]利用含杀配子基因的小麦-离果山羊草(AegilopstriuncialisL.) 3C异附加系与小滨麦异代换系M 8724、M 852杂交筛选出了3个小滨麦易位系。陆文辉等[15]、王金平等[16]通过M 842与小麦品种烟农15杂交,筛选到了1个综合性状优良、抗小麦条锈病的小滨麦易位系山农0096和1个兼抗小麦白粉病和条锈病的小滨麦易位系山农6343。本研究利用八倍体小滨麦M 842与硬粒小麦品种D 4286杂交,在F6代选育出了1个染色体数为2n=44=22Ⅱ的小滨麦衍生株系DM 5911,根尖体细胞和花粉母细胞的原位杂交研究表明,该株系含有1对完整的Ns染色体和1对易位Ns染色体,且外源Ns染色体可以稳定遗传,由此确定该株系为小滨麦附加易位系,推断其染色体组成应为2n=44=40W + 2Ns + 2T(W表示小麦染色体,Ns表示滨麦染色体,T表示易位染色体)。
本研究中,八倍体小滨麦(2n=56,AABBDDNsNs)与硬粒小麦(2n=28,AABB)进行杂交,来自八倍体小滨麦的包含单倍A、B、D、Ns基因组的配子和来自硬粒小麦的含A、B基因组的孢子结合后,F1的染色体组成为2n=42=AABB+7D+7Ns,其中,来源于八倍体小滨麦的A和B基因组与来源于硬粒小麦的A和B基因组可以很好的配对,而来自八倍体小滨麦的D和Ns基因组,由于没有可以配对的染色体而处于单价体状态,因此在杂交后代自交过程中,能完全联会配对的A、B基因组染色体可以进行正常解旋分离,从而遗传到子代;而处于单价体状态的D和Ns基因组染色体,由于处于游离状态,因此,在减数分裂形成配子时进行随机分离,并有可能丢失,自交后代中会出现染色体数不足42条的情况,我们在对F2和F3代株系进行细胞学鉴定的时候均观察到了这种现象。在自交过程中,含有相同染色体组成的配子结合后就会形成稳定的包含多种染色体组成的小滨麦衍生后代。我们已经在M 842-12与Trs-372的杂交后代中,选育出1个包含滨麦1Ns、5Ns和6Ns染色体的多重异代换系05DM6[23];在M 842-16与D 4286的杂交后代中,鉴定出了1个包含滨麦3Ns、6Ns和7Ns染色体的小滨麦多重异代换系10DM50[24]和1个小滨麦3Ns/3D二体异代换系10DM57[25]。本研究中,在M 842-16与D 4286的杂交F6代中,鉴定出了1个2n=44、包含1对完整的Ns染色体和1对易位Ns染色体附加易位系DM 5911。农艺性状调查显示,DM 5911具有株高变矮、分蘖数增多等特性,尤其是株高仅65.9 cm,比亲本均低,而滨麦的株高为60 cm[12],推测该株系中附加和易位的滨麦Ns染色体可能携带有滨麦的矮杆基因。该株系可作为进一步创制小滨麦小片段染色体易位系、研究滨麦染色体的形态结构特征及其所载滨麦农艺性状的重要种质材料,用于小麦染色体工程育种与遗传改良研究。
[1]JIANG J M,FRIBE B,GILL B S.Recent advances in alien gene transfer in wheat [J].Euphytica,1994,73:199-212.
[2]钟冠昌,穆素梅,张正斌.麦类远缘杂交[M].北京:科学出版社,2002:1-5.
ZHONG G C,MU S M,ZHANG Z B.Wheat Distance Hybridization [M].Beijing:Science Press,2002:1-5.
[3]FORSSTRÖM P O,MERKER A.Sources of wheat powdery mildew resistance from wheat-rye and wheat-Leymushybrids [J].Hereditas,2001,134:115-119.
[4]KISHII M,WANG R R C,TSUJIMOTO H.Characteristics and behaviour of the chromosomes ofLeymusmollisandL.racemosus(Triticeae,Poaceae) during mitosis and meiosis [J].ChromosomeResearch,2003,11:741-748.
[5]LIU Z P,CHEN Z Y,PAN J,etal.Phylogenetic relationships inLeymus(Poaceae:Triticeae) revealed by the nuclear ribosomal internal transcribed spacer and chloroplasttrnL-Fsequences [J].MolecularPhylogeneticsandEvolution,2008,46:278-289.
[6]YEN C,YANG J L,BAUM B R.Synopsis ofLeymusHochst.(Triticeae:Poaceae) [J].JournalofSystematicsEvolution,2009,47(1):67-86.
[7]TSITSIN N V.Remote hybridization as a method of creating new species and varieties of plants [J].Euphytica,1965,14:326-330.
[10]陈漱阳,付 杰,高立贞.普通小麦与滨麦的杂交[J].西北植物学报,1985,5(4):260-266.
CHEN S Y,FU J,GAO L Z.The hybridization betweenTriticumaestivumandLeymusmollis[J].ActaBotanicaBoreali-occidentaliaSinica,1985,5(4):260-266.
[11]傅 杰,陈漱阳,张安静.八倍体小滨麦的形成及细胞遗传学研究[J].遗传学报,1993,20(4):317-323.
FU J,CHEN S Y,ZHANG A J.Studies of the formation and cytogenetics of octoploidTritileymus[J].ActaGeneticaSinica,1993,20(4):317-323.
[12]王献平,傅 杰,张相岐,等.八倍体小滨麦染色体组构成的分子细胞遗传学研究[J].植物学报,2000,42(6):582-586.
WANG X P,FU J,ZHANG X Q,etal.Molecular cytogenetic study on genome constitutions of octoploidTritileymus[J].ActaBotanicaSinica,2000,42(6):582-586.
[13]傅 杰,陈漱阳,张安静,等.八倍体小滨麦与普通小麦杂交后代的细胞遗传学研究[J].遗传学报,1996,23(1):24-31.
FU J,CHEN S Y,ZHANG A J,etal.Cytogenetic studies on the cross progenies between octoploidTritileymusandTriticumaestivum[J].ActaGeneticaSinica,1996,23(1):24-31.
[14]傅 杰,徐 霞,杨群慧,等.八倍体小滨麦与缺体小麦杂交的细胞遗传学研究[J].遗传学报,1997,24(4):350-357.
FU J,XU X,YANG Q H,etal.Cytogenetic studies on the cross between octoploidTritileymusand nullisomic wheat [J].ActaGeneticaSinica,1997,24(4):350-357.
[15]陆文辉,林小虎,李兴峰,等.抗条锈小滨麦易位系的鉴定[J].作物学报,2005,31(1):88-91.
LU W H,LIN X H,LI X F,etal.Identification ofTritileymustranslocation line with stripe rust resistance [J].ActaAgronomicaSinica,2005,31(1):88-91.
[16]王金平,王洪刚.兼抗白粉和条锈病小滨麦种质系山农6343的细胞学和SSR鉴定[J].植物遗传资源学报,2009,10(1):46-50.
WANG J P,WANG H G.Cytological and SSR analysis ofTritileymusgermplasm line Shannong 6343 with resistance to both powdery mildew and yellow rust [J].JournalofPlantGeneticResource,2009,10(1):46-50.
[17]王献平,初敬华,张相岐.小麦异源易位系的高效诱导和分子细胞遗传学鉴定[J].遗传学报,2003,30(7):619-624.
WANG X P,CHU J H,ZHANG X Q.Efficient production of wheat alien translocation lines and characterization by molecular cytogenetics [J].ActaGeneticaSinica,2003,30(7):619-624.
[18]周兖晨,张相岐,王献平,等.滨麦抗条锈病基因的染色体定位和分子标记[J].遗传学报,2001,28(9):864-869.
ZHOU Y C,ZHANG X Q,WANG X P,etal.Chromosomal location and molecular marker of resistance gene toPucciniastriiformiswest.inLeymusmollisTrin.Hara [J].ActaGeneticaSinica,2001,28(9):864-869.
[19]赵继新,傅 杰,武 军,等.小麦-滨麦-偃麦草三属杂交后代的细胞遗传学和形态学研究[J].西北农业学报,2001,10(4):20-24.
ZHAO J X,FU J,WU J,etal.Studies on cytogenetics and morphology of the progenies of trigeneric hybridization involvingTriticum,LeymusandThinopyrum[J].ActaAgriculturaeBoreali-occidentalisSinica,2001,10(4):20-24.
[20]赵继新,陈新宏,武 军,等.普通小麦-滨麦-簇毛麦三属杂交后代的细胞遗传学研究[J].西北农林科技大学学报(自然科学版),2002,30(5):1-6.
ZHAO J X,CHEN X H,WU J,etal.Studies on cytogenetics of the progenies of trigeneric hybridization involvingTriticumaestivum,LeymusmollisandHaynaldiavillosa[J].JournalofNorthwestA&FUniversity(NaturalScienceEdition),2002,30(5):1-6.
[21]李兴锋,刘树兵,宋振巧,等.小黑麦×小滨麦后代1RS·1BL易位系的选育和鉴定[J].遗传,2004,26(4):481-485.
LI X F,LIU S B,SONG Z Q,etal.Development and molecular cytogenetic identification of 1RS·1BL translocation lines derived fromTriticale×Tritileymus[J].Hereditas,2004,26(4):481-485.
[22]赵继新,陈新宏,武 军,等.八倍体小滨麦与硬粒小麦杂交F1的细胞遗传学研究[J].西北农业学报,2005,14(2):18-22.
ZHAO J X,CHEN X H,WU J,etal.Cytogenetic studies of F1hybrid between octoploidTritileymusandTriticumdurum[J].ActaAgriculturaeBoreali-occidentalisSinica,2005,14(2):18-22.
[23]ZHAO J X,DU W L,WU J,etal.Development and identification of a wheat-Leymusmollismultiple alien substitution line [J].Euphytica,2013,190:45-52.
[24]PANG Y H,ZHAO J X,DU W L,etal.Cytogenetic and molecular identification of a wheat-Leymusmollisalien multiple substitution line from octoploidTritileymusxTriticumdurum[J].GeneticsandMolecularResearch,2014,13:3903-3913.
[25]PANG Y H,CHEN X H,ZHAO J X,etal.Molecular cytogenetic characterization of a wheat-Leymusmollis3D(3Ns) substitution line with resistance to leaf rust [J].JournalofGeneticsandGenomics,2014,41:204-214.
Development and Cytogenetics Identification of a Wheat-LeymusmollisAddition and Translocation Line DM 5911
CHENG Xueni1,LIU Yang2,PANG Yuhui2,3,LEI Zhongping1,WU Jun2,CHEN Xinhong2,YANG Qunhui2,ZHAO Jixin2
(1.College of Life Science,Northwest A&F University,Yangling,Shaanxi 712100,China; 2.The Key Laboratory of Plant Genetic Engineering and Breeding of Shaanxi Province/College of Agronomy,Northwest A&F University,Yangling,Shaanxi 712100,China;3.College of Agricultural,Henan University of Science and Technology,Luoyang,Henan 471023,China)
Leymusmollis(Trin.) Pilger(NsNsXmXm,2n=28),an excellent germplasm resource for wheat improvement,possesses many potentially valuable traits,such as strong stems,large spikes,multiple spikelets,tolerance of abiotic stresses and resistance to fungal and bacterial diseases. In this study,an addition and translocation line DM 5911 with the chromosome number of 2n=44 was developed from the F6progeny of octoploidTritileymusM 842-16 ×Triticumdurumcv. D 4286 by using of cytogenetics and morphology. Screening of mitosis and meiosis showed that DM 5911 had a chromosome karyotype of 2n=44=22Ⅱ. Mitosis GISH(genomicinsituhybridization) indicated that DM 5911 contains two whole Ns chromosomes and two translocation chromosomes between wheat andL.mollisNs genome. Meiosis GISH indicated that theL.mollisNs chromosomes perform normal synapsis,pairing and inheritance. The results suggested that DM 5911 is a cytogenetically stable wheat-L.mollisaddition and translocation line,containing a pair of whole Ns chromosome and a pair of Ns translocated into wheat chromosomes. Morphology investigation showed that DM 5911 has improved in the agronomic traits,such as plant height and the number of tillers.This line can be used as a new germplasm for creating translocation line with small chromosome fragment in wheat chromosome engineering and genetic breeding.
Wheat;Leymusmollis; Addition; Translocation; Genomicinsituhybridization(GISH)
2016-03-08
2016-03-30
陕西省自然科学基金项目(2015JM3095); 西北农林科技大学唐仲英育种基金项目
E-mail:cxn622@126.com
赵继新(E-mail:zhjx881@163.com)
S512.1;S330
A
1009-1041(2016)08-0996-07
网络出版时间:2016-08-01
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20160801.1120.008.html
