丘脑前核与记忆障碍的研究进展
2016-09-19刘晓亭朱幼玲席春华
刘晓亭, 唐 敏, 朱幼玲, 席春华
丘脑前核与记忆障碍的研究进展
刘晓亭,唐敏,朱幼玲,席春华
丘脑损伤后会出现各种记忆功能的障碍,Morris迷宫测试发现通过电刺激大鼠的丘脑前核深部,可明显改善记忆功能[1]。同样的方法电刺激癫痫患者的丘脑前核深部,也可以使受损的记忆功能得到显著提高[2]。目前的解剖已证实丘脑前核与海马、扣带回之间存在往返神经纤维联系,并且参与形成Papez环路,故丘脑前核是参与记忆过程的重要部位之一[3]。丘脑前核损害引起的记忆功能障碍主要表现为空间工作记忆及情景记忆的受损,本文就丘脑前核与记忆作一总结。
1 丘脑前核的解剖及神经纤维联系
丘脑前核由前腹侧核(Anterior ventral nuclei,AV)、前背侧核(Anterior dorsal nuclei,AD)、前内侧核(Anterior medial nuclei,AM)3个亚核组成,位于丘脑前结节的深部,其中前腹侧核最大,前内侧核次之,前背侧核最小。
丘脑前核与海马及皮质有着直接及间接的联系,其中海马发出的纤维通过穹窿到达丘脑前核为直接通路,间接通路是指由海马发出的传出纤维,通过弯窿中转后到达乳头体,后通过乳头丘脑束传出至丘脑前核。另外,海马也可以接受来自丘脑前核直接发出的纤维,因此海马与丘脑前核之间存在往返的神经纤维联系。丘脑前核也可以发出神经纤维到扣带回,海马接受来自扣带回的传出纤维,同样,扣带回也可以发出纤维到丘脑前核,因此丘脑前核与扣带回也相互联系,海马-穹窿-乳头体-乳头丘脑束-丘脑前核-扣带回-海马之间形成一个完整的神经环路。丘脑前核内不同的亚核神经纤维联系也不完全一致,内侧乳头体发出的纤维到达丘脑前腹侧核,外侧乳头体发出的纤维传出到达丘脑前背侧核,而且海马发出的神经纤维也可以被丘脑前背侧核接收。从丘脑前核的解剖及神经纤维联系发现,丘脑前核处于记忆环路的重要位置,因此丘脑前核与记忆密切相关。图为丘脑及丘脑前核的神经纤维通路(见图1)。
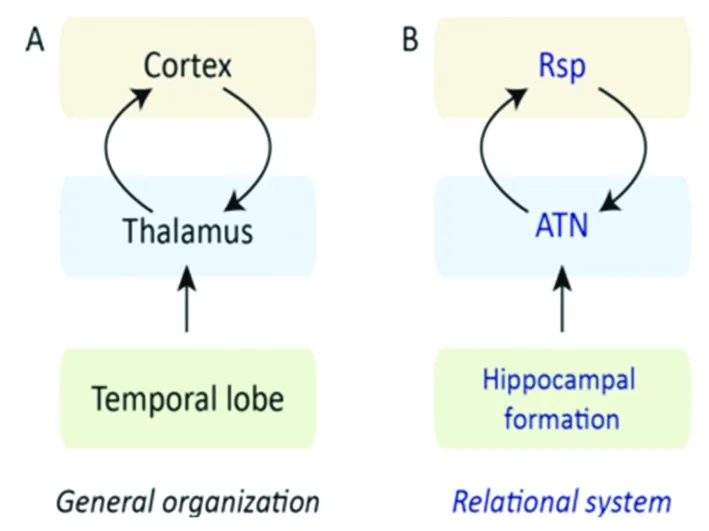
图1丘脑的神经纤维联系
2 丘脑前核与记忆的动物研究
2.1丘脑前核与空间记忆空间工作记忆又称短时记忆,目前研究认为空间工作记忆主要由以下通路完成:包括皮质-纹状体-丘脑-背侧前额叶-纹状体神经通路及枕叶皮质-颞叶下部皮质-纹状体-丘脑-腹侧前额叶-纹状体神经通路。丘脑前核损伤引起的记忆受损已被证实多次[4],而且已证明是丘脑前核不是临近核团的损伤引起的记忆障碍。丘脑背外侧、板内侧核、中央核甚至背内侧核的损害对空间工作记忆影响均不大,这进一步表明丘脑前核是参与记忆加工的重要脑区。
Morris水迷宫提供了更加准确及灵活的相关空间记忆的研究方法。例如,采用恒定的起点训练,然后把大鼠从不同的单一起点释放,来评估其空间表现的灵活性,在这一实验中即使是开始采用恒定的起点训练,丘脑前核受损的大鼠也表现出明显的缺损。但是随着测试时间延长,他们最终可以完成。不过后来在新的起点测试中却发现这些大鼠表现出不相符的行为减弱,这表明丘脑前核受损的大鼠不能够灵活应用学习过的信息[5],这一研究证明丘脑前核在空间记忆中的重要作用。支持这一观点的并不局限于Morris水迷宫实验,放射臂迷宫同样验证了丘脑前核损伤会导致工作记忆和空间记忆的损害[6]。
丘脑前核和海马都参与内侧颞叶-间脑的神经记忆通路,通路中任何环节中断都会影响记忆功能,并且很多不同记忆过程的完成都需要丘脑前核和海马相互依赖。当单侧丘脑和对侧海马受损再加上扣带皮质受损时,记忆功能缺损会更严重[7]。破坏大鼠的单侧丘脑前核和对侧海马作为实验组,同时选择同侧丘脑前核及海马同时破坏的大鼠或者仅有单侧海马受到破坏的大鼠作为对照,让这些大鼠同时在Morris水迷宫和T迷宫中进行测试,结果发现实验组大鼠的空间记忆受损明显,而对照组的空间记忆几乎不受影响,在Morris水迷宫中,丘脑前核和海马对侧的损害会引起相似的缺损症状[8]。丘脑前核损伤比穹窿损伤产生更严重的记忆缺损及搜索策略的效率下降,这表明丘脑前核的损伤引起的空间处理能力的缺损是由丘脑前核传入海马的神经纤维损害引起,不能够完全由海马通过穹窿传入丘脑前核的纤维损害来解释。穹窿部和丘脑前核的连接处受损会产生更严重的记忆功能缺损[9],这表明海马和丘脑前核之间的许多通路可以维持空间记忆。因此丘脑前核在空间记忆通路中占据重要地位,并且依赖于丘脑前核和海马的相互作用。
2.2丘脑前核与情景记忆情景记忆是发生在一定时间和地点的事件(情景)记忆,由编码、巩固、储存、提取4个过程组成,参与情景记忆的神经纤维传导的重要脑区为海马,信息的存入和取出主要由额叶及内侧颞叶负责,扣带回负责信息的成功提取[10]。额叶、颞叶、皮质下损伤均会引起情景记忆编码受损,其中丘脑投射至额叶皮质的神经纤维联系中断这一皮质下损伤会使情景记忆受损严重[11]。丘脑前核损伤不仅会导致记忆减弱,还会引起扣带皮质的功能障碍[12]。扣带皮质也参与空间记忆及情景记忆[13]。研究发现[14]恒河猴双侧丘脑损害会使情景记忆明显受损。
研究大鼠最初被要求连续闻到6种气味,随后同时给予这6种气味中的两种进行测试,大鼠必须选择一系列气味中最先闻到的气味才能获得奖励。丘脑前核损伤会破坏识别之前的气味的能力,但是不影响识别新的气味的能力。这表明[15]大鼠具有使用情景信息去处理嗅觉学习列表中任务的冲突能力,而丘脑前核损伤会妨碍动物这一功能,这表明丘脑前核在情景记忆中的作用。
2.3丘脑前核与参考记忆及学习记忆丘脑前核不仅与空间工作记忆相关,还可能与空间参考记忆相关。在Morris水迷宫中丘脑前核受损的大鼠遇到障碍后逃避反应时间显著延长,而且穿台的能力明显受损,这表明丘脑前核受损会影响空间参考记忆的过程。Aggleton等[16]通过单纯破坏大鼠的丘脑前核或者弯窿来研究结构性学习记忆的受损情况,采用空间转换实验及几何学习实验对受损的大鼠进行测评,结果发现结构性学习记忆能力无明显改变,这表明丘脑前核不参与结构性学习记忆过程。
动物实验研究表明丘脑前核损害会引起各种记忆功能受损,主要表现为空间工作记忆、空间参考记忆及情景记忆的受损,而丘脑前核在这一记忆通路中处于重要位置,并且与海马扣带回之间相互依赖。
3 丘脑前核亚核及生化
为了明确丘脑前核中三大亚核在功能上的区别与联系,开放式水迷宫的研究中发现[17],注射鹅膏覃氨酸破坏大鼠丘脑前核中单独一个核团或者同时破坏两个核团,其空间工作记忆仅轻度受损,然而丘脑前核三大亚核同时受到破坏时,大鼠的空间工作记忆能力明显减弱,因此丘脑前核的三大亚核之间相互依赖,丘脑前核单个亚核的损害不会引起空间工作记忆的严重受损,这表明丘脑前核三大亚核共同作为整体参与空间学习记忆。
丘脑和认知的神经环路模式经过亲代谢型谷氨酸受体发挥作用[18]。有研究发现[1]电刺激大鼠的丘脑前核深部,p-细胞外调节蛋白激酶的表达会较前明显增加,这表明丘脑前核通过细胞内的信号转导参与记忆的过程。
4 丘脑前核与记忆的临床研究
周爱红等[19]对丘脑前核卒中的8例老年患者进行了广泛记忆功能测评,通过左侧与右侧对比表明,左侧丘脑前核可能参与言语性记忆过程及延时回忆过程,而右侧损伤引起的言语性记忆障碍比左侧微弱。也有研究表明双背内侧丘脑损伤的患者而没有丘脑前核的损伤,却发现其出现典型的记忆障碍综合征,包括顺行性及逆行性记忆,而内隐性记忆相对保留[20]。临床对于丘脑不同核团与各种记忆的神经通路及机制的研究目前尚少,有待进一步的研究。
5 回顾与展望
丘脑前核损害会引起记忆功能受损,主要表现为空间工作记忆、空间参考记忆及情景记忆的受损,而丘脑前核在这一记忆通路中处于重要位置,并且与海马、扣带回之间相互依赖。丘脑前核参与形成Papez环路,这可以解释丘脑前核在空间记忆及工作记忆中的作用。丘脑前核不同核团之间作为一个整体影响空间学习记忆,然而对于不同核团与记忆之间的区别与联系仍需要进一步的探讨。丘脑前核是否参与其它类型的记忆,丘脑其他核团是否与记忆有关,目前的研究均尚少。丘脑各核团参与记忆的神经机制目前尚存在争议,关于丘脑参与记忆的加工过程以及参与记忆加工的具体神经通路目前也尚不明确。从目前的实验来看,动物实验居多,且目前临床多以丘脑为观察对象,细分至丘脑前核的研究少,临床患者没有大样本数据观察和解释丘脑前核与记忆的关系。下一步的研究可针对丘脑各个核团损伤的患者进行各种记忆功能的评估,借助高分辨率磁共振对丘脑核团进行更精确的定位及细分,采用功能磁共振及其他影像学方法观察各种记忆的功能联系,为观察及干预丘脑前核在各种通路中的作用提供影像学的依据。
[1]Chen N,Dong S,Yan T,et al.High-frequency stimulation of anterior nucleus thalamus improves impaired cognitive function induced by intea-hippocampal injection of Aβ1-40in rats[J].Chin Med J (Engl),2014,127(1):125-129.
[2]Bokova M,Chladek J,Jurak P,et al.Complex motor-cognitive factors processed in the anterior nucleus of the thalamus:an intracerebral recording study[J].Brain Topogr,2015,28(2):269-278.
[3]Aggleton JP.Looking beyond the hippocampus:old and new neurological targets for understanding memory disorders[J].Proc Biol Sci,2014,281(1786):129-143.
[4]Moreau PH,Tsenkina Y,Lecourtier L,et al.Lesions of the anterior thalamic nuclei and intralaminar thalamic nuclei:place and visual discrimination learning in the water maze[J].Brain Struct Funct,2013,218(3):657-667.
[5]Wolff M,Loukavenko EA,Will BE,et al.The extended hippocampal diencephalic memory system:enriched housing promotes recovery of the flexible use of spatial representations after anterior thalamic lesions[J].Hippocampus ,2008,18(10):996-1007.
[6]Mitchell AS,Dalrymple-Alford JC.Dissociable memory effects after medial thalamus lesions in the rat[J].Eur J Neurosci,2005,22(4):973-985.
[7]Dumont JR,Petrides M,Sziklas V.Fornix and retrosplenial contribution to a hippocampo-thalamic circuit underlying conditional learning[J].Behav Brain Res,2010,209(1):13-20.
[8]Henry J,Petrides M,St-Laurent M,et al.Spatial conditional associative learning:effects of thalamo-hippocampal disconnection in rats[J].Neuroreport,2004,15(15):2427-2431.
[9]Warburton EC,Baird AL,Morgan A,et al.Disconnecting hippocampal projections to the anterior thalamus produces deficits on tests of spatial memory in rats[J].Eur J Neurosci,2000,12(5):1714-1726.
[10]Olson IR,Berryhill M.Some surprising findings on the involvement of the parietal lobe in human memory[J].Neurobiol Learn Mem,2009,91(2):155-165.
[11]郭宗君,肖琳,田玉波,等.脑梗死患者情景记忆编码损伤研究[J].中华行为医学与脑科学杂志,2010,19(12):1060-1062.
[12]Aggleton JP.Understanding retrosplenial amnesia:insights from animal studies[J].Neuropsychologia,2010,48(8):2328-2338.
[13]Hindley EL,Nelson AJ,Aggleton JP,et al.Dysgranular retrosplenialcortex lesions in rats disrupt cross-modal object recognition[J].Learn Mem,2014,21(3):171-179.
[14]Mitchell AS,Caftan D.The magnocellular mediodorsal thalamus is necessary for memory acquisition,but not retrieval[J].J Neurosci,2008,28:258-263.
[15]Law LM,Smith DM.The anterior thalamus is critical for overcoming interference in a context-dependent odor discrimination task[J].Behav Neurosci,2012,126(5):710-719.
[16]Aggleton JP,Poirier GL,Aggleton HS,et al.Lesions of the fornix and anterior thalamic nuclei dissociate different aspects of hippocampal-dependent spatial learning:Implications for the neural basis of scene learning[J].Behav Neurosci,2009,123(3):504-519.
[17]Van Groen T,Kadish I,Michael Wyss J.Role of the anterodorsal and anteroventral nucei of the thalamus in spatial memory in the rat[J].Behav Brain Res,2002,32(1):19-28.
[18]Copeland CS,Neale SA,Salt TE.Neuronal activity pattens in the mediodorsal thalamus and related cognitive circuits are modulated by metabotropic glutamate receptors[J].Neuropharmacology,2015,92(13):16-24.
[19]周爱红,魏翠柏,贾建平.不同侧别丘脑前核群卒中所致认知损害的特征[J].脑与神经疾病杂志,2010,18(6):401-404.
[20]Hampstead BM,Koffler SP.Thalamic contributions to anterograde,retro-grade,and implicit memory:a case study[J].Clin Neuropsychol,2009,23(7):1232-1249.
1003-2754(2016)08-0766-03
R742
教学园地
2016-04-11;
2016-05-29
国家自然科学基金项目(No.81301095);合肥市第四周期医学重点学科项目(卫科2013-206号)
(安徽医科大学第三附属医院,合肥市第一人民医院神经内科,安徽 合肥 230061)
朱幼玲,E-mail:zhuyoulingmail@126.com
