天敌昆虫在诱集植物上的多样性及对温室蚜虫的防治作用
2016-09-16吴圣勇徐丽荣王登杰雷仲仁
吴圣勇,徐丽荣,李 宁,王登杰,雷仲仁,4
(1中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室,北京 100193;2湖北黄冈市农业科学院,湖北黄冈 438000;3四川省达州市达川区植保植检站,四川达州 635000;4闽台特色作物病虫生态防控协同创新中心,福州350002)
天敌昆虫在诱集植物上的多样性及对温室蚜虫的防治作用
吴圣勇1,徐丽荣2,李 宁2,王登杰3,雷仲仁1,4
(1中国农业科学院植物保护研究所植物病虫害生物学国家重点实验室,北京 100193;2湖北黄冈市农业科学院,湖北黄冈 438000;3四川省达州市达川区植保植检站,四川达州 635000;4闽台特色作物病虫生态防控协同创新中心,福州350002)
【目的】针对天敌昆虫的保护和利用,通过混合种植不同诱集植物,分析天敌昆虫的多样性特征及与其影响因子间的关系;通过筛选和贮存诱集植物上的优势天敌,并构建储蓄植物系统,应用于温室黄瓜中,评价其对目标害虫蚜虫的防治效果。【方法】2012、2013和2015年分别在河北廊坊和湖北黄冈两试验基地混合种植5种诱集植物:玉米、紫花苜蓿、向日葵、小麦和大豆。通过调查天敌昆虫类群,计算2012和2015年的天敌群落特征参数,并用冗余分析法(redundancy analysis,RDA)分析天敌群落结构与诱集植物种类、调查时间和植物花期的关系;2013年通过收集并贮存诱集植物中的优势天敌瓢虫,构建出小麦-麦蚜-瓢虫的储蓄植物系统,并在系统中接种不同密度瓢虫后应用于温室黄瓜中防治蚜虫,比较各瓢虫密度下的储蓄植物系统对蚜虫的防治效果。【结果】诱集植物上天敌昆虫的Shannon-Wiener多样性指数、Simpson优势集中性指数和Pielou均匀性指数总体上随着调查时间的变化呈现显著差异;通过对天敌的等级划分,蚜茧蜂和龟纹瓢虫为优势类群,二者的个体数量均占所有诱集天敌数量的10%以上;RDA分析结果表明,诱集植物种类、调查时间和植物花期对天敌群落结构均有显著影响,三者总和对2012和2015年天敌多样性信息的总变异可分别解释26.02%和17.20%;2012和2015年的RDA排序图均表明,小花蝽和中华草蛉与植物花期相关性较高;将小麦-麦蚜-瓢虫的储蓄植物系统应用于温室后的第2周,接种200头瓢虫的处理,蚜虫密度显著低于其他处理,防治效果为69.4%。【结论】采用协调天敌诱集和助增策略,通过混合种植不同诱集植物,诱集到了多种自然天敌类群,并将筛选出的优势天敌构建成储蓄植物系统,应用于温室黄瓜中,其中,接种高密度瓢虫的储蓄植物系统对蚜虫具有一定的控制作用。因此,以天敌昆虫诱集、保护、筛选和利用为一体的害虫生物防治思路在实践中具有可行性。
诱集植物;天敌昆虫;冗余分析;储蓄植物系统;释放应用;蚜虫
0 引言
【研究意义】天敌昆虫作为自然生态系统中的组成部分,在调控农业害虫种群方面具有重要的生态功能[1]。由于传统的化学农药在害虫防治中并不具有选择性,从而杀害了大量的天敌昆虫,破坏了生态系统,显著降低了自然天敌的控害效果[2]。通过生境管理,即在农业生态系统中建立适宜的生态基础设施,为天敌昆虫在不利的自然环境中提供食物资源(花粉、蜜露、替代猎物/寄主)或庇护场所,可提高天敌昆虫的种群和害虫生物防治能力。其中,种植天敌昆虫的诱集植物是应用较为广泛且相对成功的措施[3]。此外,助增天敌昆虫的主动调控作用,如构建储蓄植物系统(banker plant system)(基于陈学新等[4]的建议,本文统一用“储蓄植物系统”作为中文名称),可显著提高天敌昆虫控害的功效[5-6]。采用协调天敌诱集和助增策略,将天敌诱集与温室应用结合,可为利用天敌昆虫控制害虫提供理论和实践基础。【前人研究进展】THIES等[7]早在1999年就研究发现,景观作物的丰富性有助于天敌昆虫的迁移和对周围作物害虫的持续防控。此外,在保护地作物周围种植开花性植物,可为多种天敌昆虫,如瓢虫、草蛉、食蚜蝇和寄生蜂等提供补充食物,从而显著提高天敌昆虫的寿命和繁殖率[8-10]。进一步研究发现,很多捕食性和寄生性天敌对花粉和蜜露释放出的挥发性有机物表现出明显的趋性[9,11-13],LI等[14]研究发现,在作物或天敌储蓄植物中添加花粉对小花蝽和瓢虫具有明显的诱集作用。关于用储蓄植物增殖和释放天敌的措施防治害虫已有 40年的历史[4],多种天敌昆虫,如蚜茧蜂(Aphidiid)、丽蚜小蜂(Encarsia formosa)、食蚜瘿蚊(Aphidoletes aphidimyza)、食蚜蝇 (Scaeva pyrastri)和植绥螨(Phytoseiidae)等都有应用,防治对象包括蚜虫、粉虱、蓟马和叶螨等[5,15]。【本研究切入点】作物花期对天敌昆虫具有诱集性[11],且增加作物种类有助于天敌迁移和栖息以及提高天敌多样性[16],因此,本研究通过3年在不同地点混合种植5种诱集植物,分析所诱集天敌的多样性特征;此外,将诱集到的优势天敌昆虫通过储蓄植物繁育后,应用于温室黄瓜中防治瓜蚜。【拟解决的关键问题】分析诱集植物中的天敌多样性特征及与诱集植物种类、调查时间和植物花期的关系,筛选出优势天敌种类,并在温室中应用,评价其对黄瓜蚜虫的防治效果。
1 材料与方法
天敌昆虫的诱集试验于2012和2013年在河北省廊坊市的中国农业科学院植物保护研究所试验基地(116°69′ E,39°52′ N)开展,2015在湖北省黄冈市农业科学院梅家墩试验基地(114°55′ E,30°34′ N)开展。储蓄植物系统的温室应用于2013年在河北省廊坊市中国农业科学院植物保护研究所试验基地进行。
1.1供试植物
诱集植物包括玉米、紫花苜蓿、向日葵、小麦和大豆。其中,玉米、紫花苜蓿和向日葵种子购自北京鑫农丰农业技术研究所;小麦和大豆分别为当地试验基地提供。
1.2天敌昆虫诱集试验
在两个试验基地中,每种作物设置一个小区,小区面积为3 m×3 m,小区重复3次,5种作物共15个小区,按照随机区组排列,每个小区间隔1 m(图1),试验区周围为常规作物。根据诱集植物的生长周期差异,先后安排种植不同作物。试验期间,小麦处于苗期,玉米处于抽雄吐丝期,紫花苜蓿和向日葵经历现蕾期和花期,大豆经历出枝期、开花期和结荚期。调查采用5点取样法,每点选择0.5 m×0.5 m的面积,其中,玉米、向日葵和大豆每点取样6株,用直接观察法调查取样点内的所有天敌种类。调查时同时记录作物的生长时期。每周调查一次,若遇到阴雨天气,提前或延后1 d调查。廊坊调查时间为2012年5月30日至7月3日(由于收集天敌会影响调查结果,所以2013年只收集天敌而不做调查,并构建储蓄植物系统和开展温室应用试验),黄冈调查时间为2015年7月18日至8月21日。试验调查期间,诱集植物的栽培管理措施按照常规农事操作进行,每天用滴灌浇水,全苗生育阶段,不定期锄草,不使用除草剂和任何药剂。
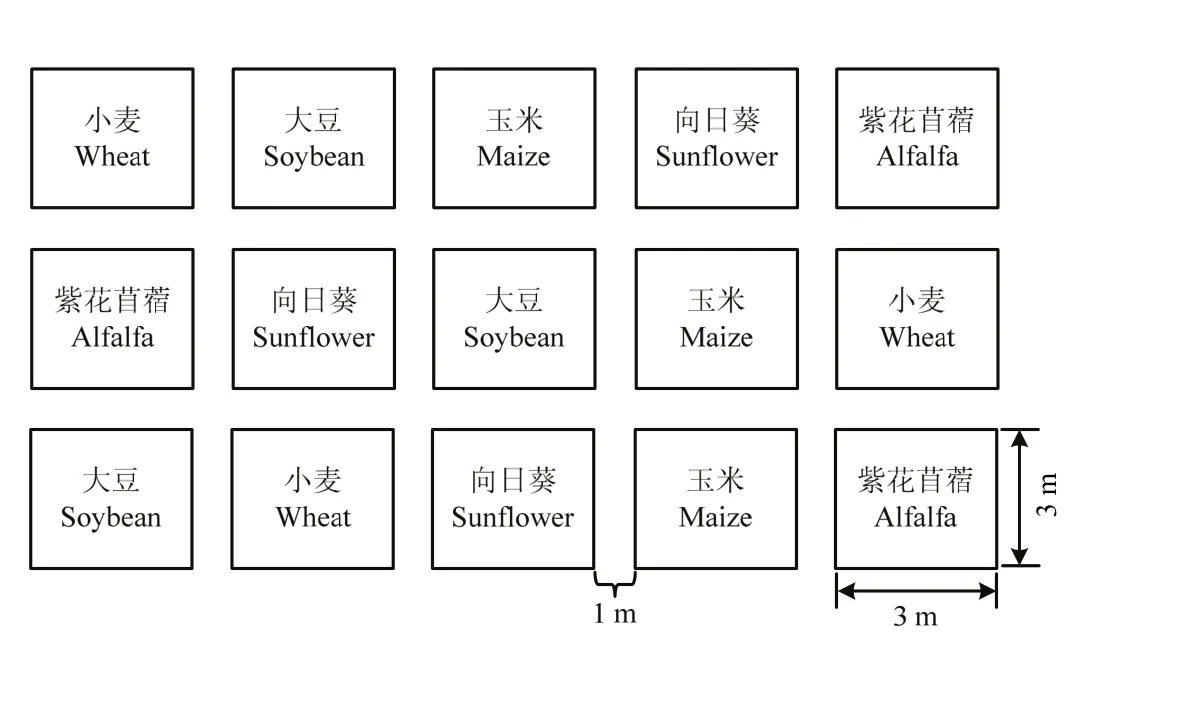
图1 2012、2013和2015年诱集植物小区分布Fig. 1 Distribution of plots for trapping crops in 2012, 2013 and 2015
1.3储蓄植物系统在温室中的应用
基于2012年廊坊试验基地天敌昆虫的诱集效果,2013年将诱集植物中的瓢虫卵块(包括龟纹瓢虫(Propylaea japonica)、异色瓢虫(Harmonia axyridis)、七星瓢虫(Coccinella septempunctata)和二星瓢虫(Adalia bipunctata)收集起来,放入透明塑料杯(上口直径10 cm,下口直径6 cm,高16 cm)中,上口用纱布封盖,然后置于温度为(10±1)℃,相对湿度(70±5)%,光周期L∶D=16 h∶8 h的人工气候箱中贮存备用。
温室应用前,将培养箱温度调至(26±1)℃,待卵孵化后,将1—2龄期的幼虫接入带有麦蚜的小麦上(小麦提前移栽到盆口直径为20 cm的花盆中),并用100目的纱网笼罩,即构建出了以麦蚜为替代猎物,小麦为储蓄植物的小麦-麦蚜-瓢虫储蓄植物系统,并应用于温室黄瓜中防治蚜虫,即瓜蚜(Aphis gossypii)。试验设置3个瓢虫密度梯度,A:50头;B:100头;C:200头;D:空白对照。将蚜虫发生密度相对均匀的区域划分为12个小区,每区为一畦,每畦25颗黄瓜,小区之间用100目的防虫网隔开。小区随机排列。每个小区中间安排一个处理,每个处理重复3次。在每个小区内定点调查5株黄瓜苗,每株选取上、中、下相对位置上各一片叶子,调查整张叶片正反面上的所有活蚜虫数量,每周调查一次,连续调查两周。储蓄植物系统应用前,先调查各处理中的蚜虫基数。
1.4数据处理
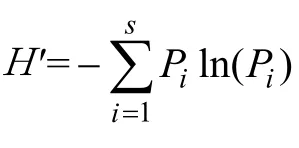
采用冗余分析(redundancy analysis,RDA)方法分析不同寄主作物上的天敌群落与其影响因子(诱集植物种类、调查时间和植物花期)间的关系[19]。冗余分析是约束排序中的重要类型,能够很好地分析物种多样性特征(群落结构和分布)与其影响因子的关系,而且可以降低个别优势类群对群落结构的影响权重。在采用冗余分析前,先利用除趋势对应分析(detrended correspondence analysis,DCA)对不同日期下不同诱集植物上的天敌昆虫群落组成进行调查,根据得到的排序轴的梯度长度(lengths of gradient)来选择排序的类型(线性模型(RDA)或者单峰模型(CCA))。如果DCA 排序前4个轴中的最大值超过4,选择单峰模型合适,如果<3选择线性模型比较合适;如果介于3—4两种模型都可行[20]。在本研究中,2012年和 2015年寄主作物上的天敌分布的梯度长度最大值分别为1.17和1.42,均小于3。利用R语言中vegan 软件包的‘diversity’函数计算H′、D和J。利用vegan软件包的‘rda’函数实现RDA分析[21]。所有原始数据均经过lg(x+1)转化。
不同调查日期间的天敌群落特征参数用单因素方差分析(One-way ANOVA,SPSS13.0),方差若有显著性,采用Turky-HSD法比较,显著性差异为P=0.05水平。
储蓄植物系统防治温室蚜虫的效果,根据应用前后各小区的活蚜数进行统计,并按下列公式计算其虫口减退率(%)及防治效果(%):
虫口减退率(%)= [(防治前虫口数-防治后虫口数)/防治前虫口数]×100
防治效果(%)= [(处理区虫口减退率-对照区虫口减退率)/(1-对照区虫口减退率)]×100
不同处理下的蚜虫防治效果用单因素方差分析(One-way ANOVA,SPSS13.0),方差若有显著性,采用Turky-HSD法比较,显著性差异为P=0.05水平。
2 结果
2.1天敌昆虫多样性特征及与其影响因子间的关系
诱集植物上天敌昆虫的群落特征参数,Shannon-Wiener多样性指数、Simpson优势集中性指数和Pielou均匀性指数随着调查时间的变化,总体上呈现显著差异(图2)。按照个体数量占到所有天敌数量的10%以上为优势类群的划分依据[22],2012年的优势类群为蚜茧蜂、小花蝽和龟纹瓢虫;2015年的优势类群为蚜茧蜂和龟纹瓢虫。其中,两年的蚜茧蜂类群分别占22.5%和14.7%。
图3为RDA排序图,箭头表示影响因子,箭头连线在两个轴上的投影长度代表某个影响因子对研究对象影响的大小,连线在轴上的投影越长代表该影响因子对研究对象的分布影响越大,箭头连线与排序轴的夹角代表该影响因子与排序轴的相关性大小,夹角越小则相关性越高。2012和2015年的天敌昆虫RDA分析结果表明,2012年,不同诱集植物上的天敌昆虫多样性信息总变异中的26.02%能够被诱集植物种类、调查时间和植物花期这 3个变量解释(F=3.048,P=0.001),其中第一轴解释了约 15.00%变异信息,第二轴解释了约 7.07%变异信息,第三轴解释了约3.95%变异信息。2015年,诱集植物种类、调查时间和植物花期能够解释 17.20%的天敌昆虫多样性信息的总变异(F=1.801,P=0.027),其中第一轴解释了约11.50%变异信息,第二轴解释了约4.20%变异信息,第三轴解释了约1.50%的变异信息。
诱集植物种类、调查时间和植物花期对天敌群落结构均有显著影响(2012年,诱集植物:r2=0.476,P <0.001;调查时间:r2=0.484,P<0.001;植物花期:r2=0.455,P<0.001;2015年:诱集植物:r2=0.604,P=0.001;调查时间:r2=0.218,P=0.029;植物花期:r2=0.215,P=0.044)。两年的PDA排序图反映出一个共同的信息,即小花蝽和中华草蛉(Chrysoperla sinica)与植物花期的相关性较高。
2.2储蓄植物系统对温室蚜虫的防治效果
2013年温室试验之前,各处理中的蚜虫初始密度没有显著差异(F3,8=1.79,P=0.23),平均每片叶子上蚜虫为102头(图4)。构建的小麦-麦蚜-瓢虫的储蓄植物系统,在应用后的第一周,接种200头瓢虫的处理对蚜虫种群具有压制作用,但各处理间的蚜虫密度没有显著差异(F3,8=2.65,P=0.12),两周后,接种200头瓢虫的处理中的蚜虫密度显著低于其他处理(F3,8=5.18,P=0.03),防治效果为69.4%(表1)。
3 讨论
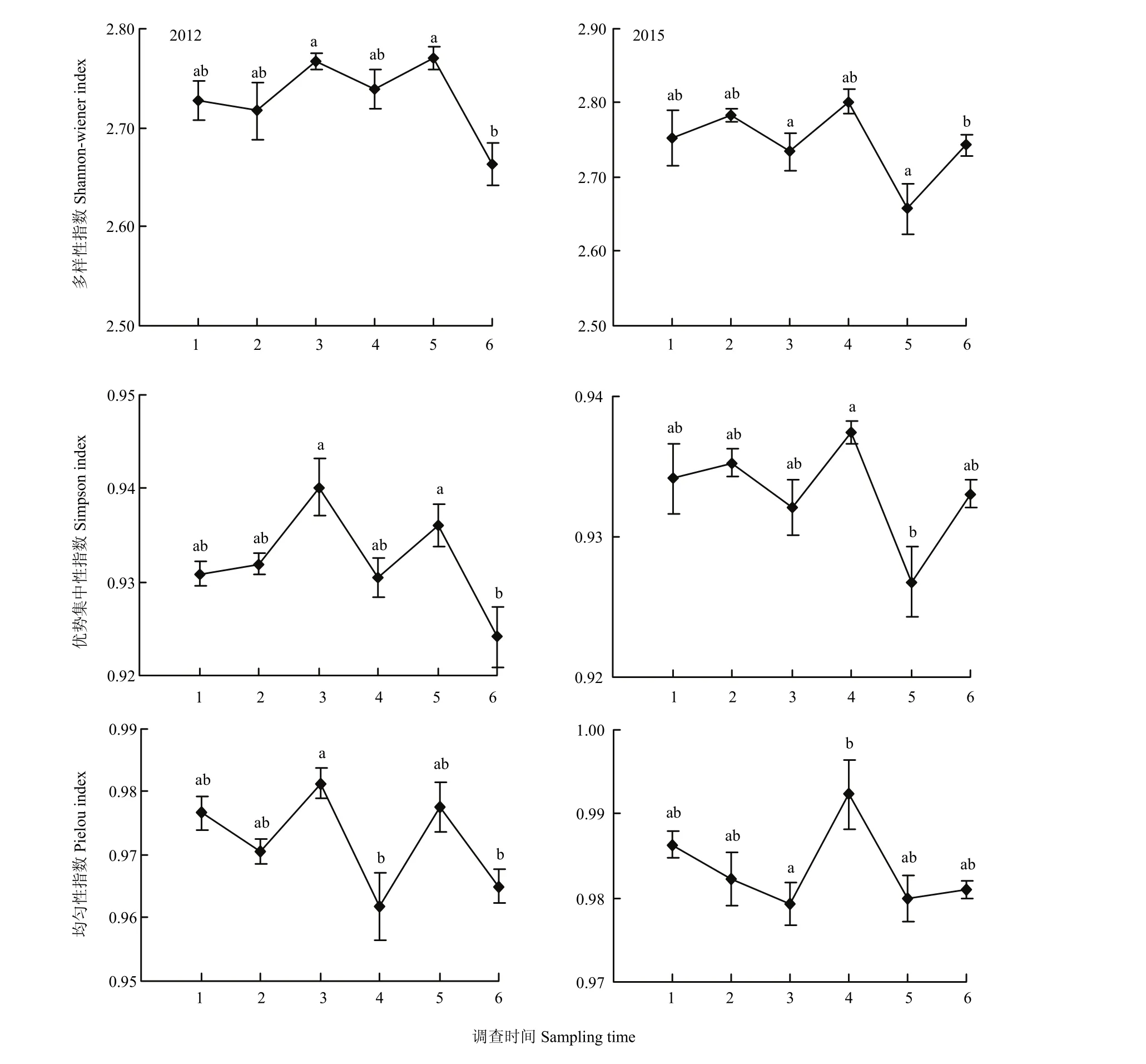
图2 2012和2015年诱集植物上天敌多样性指数的时间动态Fig. 2 Temporal dynamics of natural enemy diversity on trapping crops in 2012 and 2015

表1 接种不同瓢虫密度的储蓄植物系统对蚜虫的防治效果Table 1 Control efficacy of banker plant system inoculated at different densities of ladybird beetle against A. gossypii

图3 诱集植物上天敌群落结构与其影响因子间RDA排序图Fig. 3 Graph of RDA for natural enemy community structure and its factors
近年来,基于害虫生物防治的天敌昆虫控害机理和可持续利用方面,国内外开展了大量的工作[2,23-24]。其中,利用天敌昆虫对害虫的自然调控是今后研究的热点方向[10]。相比于害虫,天敌昆虫的不同种群对地理分布、生态环境和人为干扰等方面存在不对称性或不同步行,从而导致天敌昆虫对害虫的防控效应存在“时空错位”和“控制乏力”[24]。由于天敌昆虫对害虫的“跟随效应”以及化学农药对自然天敌的杀伤作用,因此,通过人为诱集、保育和助增等方式,对天敌昆虫种群进行主动调节,并通过促进天敌昆虫与害虫的时空同步性,保持对害虫的“高压”态势,是害虫可持续控制的重要措施。
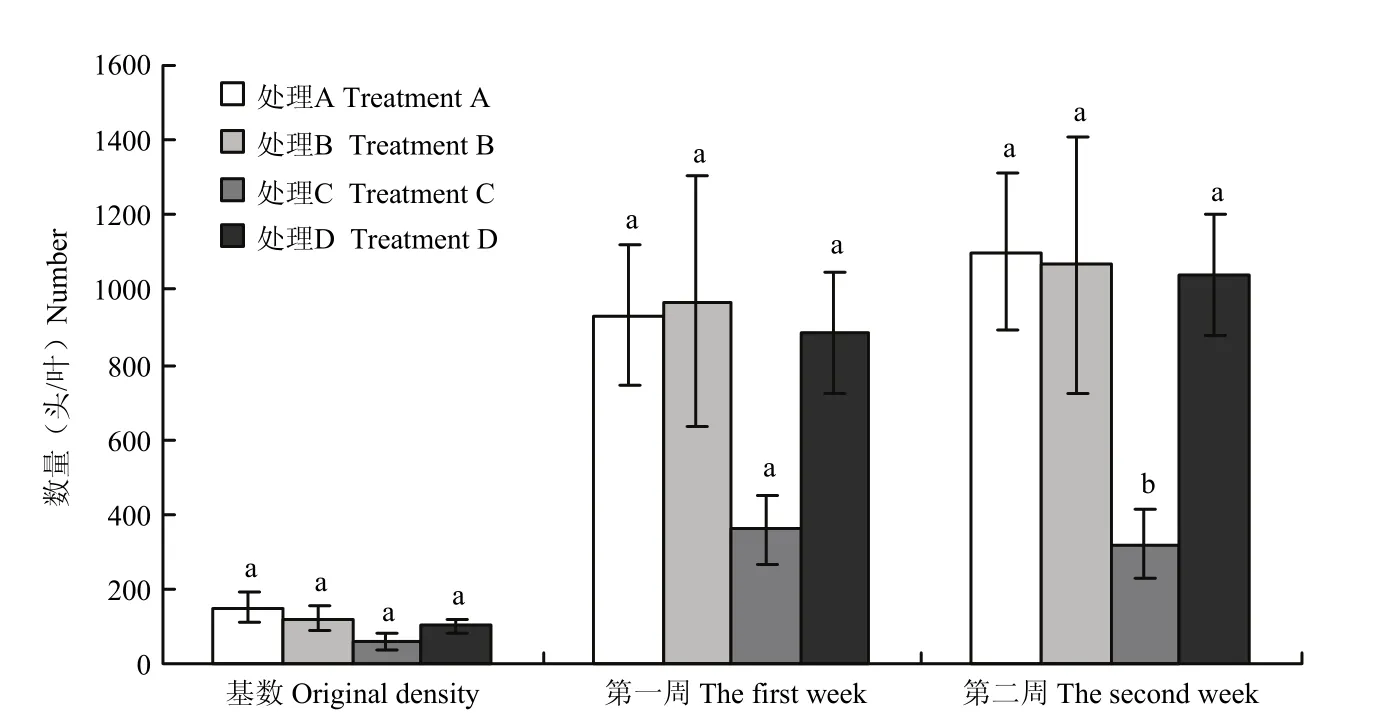
图4 接种不同瓢虫密度的储蓄植物系统处理中蚜虫种群数量Fig. 4 A. gossypii population in treatments of banker plant system inoculated at different densities of ladybird beetle
开花作物、蜜露可为天敌昆虫提供补充食物,从而有助于天敌的诱集和定殖[11,25-26],作物多样性影响天敌昆虫的食物网[27-28]和在作物上的天敌种类、种群结构和分布[16,29-30],本研究在不同地点混合种植的 5种诱集植物中,玉米、向日葵和紫花苜蓿均具有显著的诱集天敌作用[3,31-32],大豆和小麦对诱集害虫效果较好,从而起到丰富天敌食物网的作用[33]。相对于传统的生态系统中生物多样性的分析,本研究采用的冗余分析(RDA)是一种直接梯度分析方法,该方法能够在不同变量因子组合下对单个变量进行统计学特征描述,从而揭示各变量因子对生物多样性的贡献率[34]。本研究不仅将作物种类和调查时间作为常规的影响因子分析其对天敌昆虫多样性的影响,并且将植物花期也作为一个重要影响因子进行研究。2012和2015年的研究结果表明,作物种类、调查时间和植物花期共对天敌昆虫多样性信息的总变异可分别解释 26.02% 和17.20%,即80%左右的变异受到其他因子的影响,如作物的种植面积和周围植被[33]、农业景观格局[35-36]等对昆虫群落均有影响。此外,气候条件、地理位置、作物生长阶段和季节动态等也对诱集植物上的天敌多样性产生影响。因此,作物对天敌昆虫的诱集效果应在大尺度空间范围内综合考虑。本研究所在的廊坊和黄冈试验基地分别占地33和60 hm2,两个基地主要种植的作物均为棉花、小麦、玉米、花生、大豆和蔬菜等作物,种植结构相似,所以诱集的天敌昆虫类群相近,如试验过程中,两基地的小麦中蚜茧蜂数量较多,成为诱集植物上的优势群体,此外,廊坊和黄冈分别在试验的第6周和第5周降雨,对天敌种群的影响较大,群落参数均明显下降。
很多研究表明,提高作物多样性能为天敌提供更适合的微观环境、食物、替代寄主或猎物等资源[3],尤其是蜜源植物能够增加天敌的多样性和适合度[37-38],然而,天敌间存在兼性捕食/寄生作用(intraguild predation,IGP)[39],此外,很多植物也同时具有对害虫的诱集作用[33],因此,天敌多样性的提高并不意味着能够降低害虫种群[40-41]。鉴于诱集植物在天敌昆虫利用方面具有风险,本研究通过在自然生态系统中将几种诱集植物组合,在田间“招募”各类自然天敌,形成天敌昆虫的开放式储蓄库,之后针对目标作物上害虫的发生种类与发生密度,从天敌库中有意识地“调用”优势天敌昆虫,并临时构建出储蓄植物系统,应用于目标作物上防治害虫,探索集天敌诱集、保护、筛选与释放应用为一体的天敌昆虫利用新途径。2012和2013年在廊坊种植的诱集植物中,蚜茧蜂和瓢虫均为优势天敌类群,且在植物上以僵蚜、瓢虫卵和低龄幼虫的定殖虫态容易收集和贮存。本研究在2013年的诱集植物中,仅收集了瓢虫卵,并就地取材,将带有麦蚜的小麦作为储蓄植物,从而构建出“小麦-麦蚜-瓢虫”储蓄植物系统。2015年,在黄冈种植了同样布局的诱集植物,进一步验证了诱集结果。该研究思路不仅节约了天敌大量饲养的成本,避免了天敌利用的风险,也为生态系统中提高天敌功效所需的功能植物的筛选、搭配和布局提供参考。
储蓄植物系统综合了天敌的保护与助增的特点[5],即通过建立一个自我维持的系统以降低有害生物的种群水平[15]。储蓄植物系统的应用时间直接决定着对害虫的防治效果[42],提早预防性引入储蓄植物系统,往往会取得明显的防治效果,且成本较低。本研究在温室中引入储蓄植物时,黄瓜叶片上的蚜虫密度已经较高,所以没有取得良好的防治效果。试验中,尽管接种200头瓢虫的储蓄植物在第2周的防治效果近70%,仍压制不住蚜虫种群的暴发,后期植物由于蚜虫的严重为害而枯萎。因此,诱集植物的种植、天敌昆虫的收集和保存、储蓄植物的构建和引入应综合考虑目标作物害虫的发生时间和发生密度等信息。
目前,中国的农业害虫,尤其是入侵性害虫表现出对环境的适应性高、繁殖速度快和竞争能力强等特点[43]。通过保护和利用天敌,充分发挥天敌对害虫的持续有效控制是生物防治研究中一个始终不渝的努力方向[4]。本研究协调天敌诱集和助增策略,在小范围内探索了诱集植物上天敌多样性特征及与其影响因子间的关系,并以温室黄瓜害虫(蚜虫)为实践对象,将诱集到的优势天敌构建成储蓄植物系统,评价了其对温室蚜虫的防治效果,为天敌昆虫的保护和利用提供了理论和实践基础。
4 结论
诱集植物种类、调查时间和植物花期显著影响天敌昆虫的群落结构;构建的优势天敌储蓄植物系统,在应用时应考虑目标害虫的发生时间和密度。以天敌昆虫诱集、保护、筛选和利用为一体的害虫生物防治思路在实践中具有可行性。
References
[1] NAEEM S, BUNKER D E, HECTOR A, LOREAU M, PERRINGS C. Biodiversity, Ecosystem Functioning, and Human Wellbeing: An Ecological and Economic Perspective. Oxford: Oxford University Press, 2009.
[2] CROWDER D W, NORTHFIELD T D, STRAND M R, SNYDER W E. Organic agriculture promotes evenness and natural pest control. Nature, 2010, 466 (7302): 109-112.
[3] LANDIS D A, WRATTEN S D, GURR G M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annual Review of Entomology, 2000, 45(1): 175-201.
[4] 陈学新, 刘银泉, 任顺祥, 张帆, 张文庆, 戈峰. 害虫天敌的植物支持系统. 应用昆虫学报, 2014, 51(1): 1-12. CHEN X X, LIU Y Q, REN S X, ZHANG F, ZHANG W Q, GE F. Plant-mediated support system for natural enemies of insect pests. Chinese Journal of Applied Entomology, 2014, 51(1): 1-12. (in Chinese)
[5] FRANK S D. Biological control of arthropod pests using banker plant systems: Past progress and future directions. Biological Control, 2010,52(1): 8-16.
[6] XIAO Y F, OSBORNE L, CHEN J , MCKENZIE C, HOUBEN K,IRIZARRY F. Evaluation of corn plant as potential banker plant for supporting predatory gall midge, Feltiella acarisuga (Diptera:Cecidomyiidae) against Tetranychus urticae (Acari: Tetranychidae) in greenhouse vegetable production. Crop Protection, 2011, 30(12):1635-1642.
[7] THIES C, TSCHARNTKE T. Landscape structure and biological control in agroecosystems. Science, 1999, 285(5429): 893-895.
[8] HICKMAN J M, WRATTEN S D. Use of Phacelia tanacetifolia strips to enhance biological control of aphids by hoverfly larvae in cereal fields. Journal of Economic Entomology, 1996, 89(4): 832-840. [9] WÄCKERS F L. Assessing the suitability of flowering herbs as parasitoid food sources: flower attractiveness and nectar accessibility. Biological Control, 2004, 29(3): 307-314.
[10] BIANCHI F J, BOOIJ C J, TSCHARNTKE T. Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proceedings of the Royal Society B: Biological Sciences, 2006, 273(1595): 1715-1727.
[11] WINKLER K, WÄCKERS F, BUKOVINSZKINE-KISS G, VAN LENTEREN J. Sugar resources are vital for Diadegma semiclausum fecundity under field conditions. Basic and Applied Ecology, 2006,7(2): 133-140.
[12] OSAWA N. Ecology of Harmonia axyridis in natural habitats within its native range. BioControl, 2011, 56(4): 613-621.
[13] WAJNBERG E, COLAZZA S. Chemical Ecology of Insect Parasitoids.John Wiley and Sons, Inc., 2013.
[14] LI S, TAN X, DESNEUX N, BENELLI G, ZHAO J, LI X H,ZHANG F, GAO X W, WANG S. Innate positive chemotaxis to pollen from crops and banker plants in predaceous biological control agents: towards new field lures? Scientific Reports, 2015, 5: Article number 12729.
[15] 肖英方, 毛润乾, 沈国清, OSBORNE L S. 害虫生物防治新技术——载体植物系统. 中国生物防治学报, 2012, 28(1): 1-8. XIAO Y F, MAO R Q, SHEN G Q, OSBORNE L S. Banker plant system: a new approach for biological control of arthropod pests. Chinese Journal of Biological Control, 2012, 28(1): 1-8. (in Chinese)
[16] JOHNSON M T, LAJEUNESSE M J, AGRAWAL A A. Additive and interactive effects of plant genotypic diversity on arthropod communities and plant fitness. Ecology Letters, 2006, 9(1): 24-34.
[17] 赵志模, 郭依泉. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 1990. ZHAO Z M, GUO Y Q. Community Ecology Principles and Methods. Chongqing: Chongqing Bureau of Science and Technology Press,1990. (in Chinese)
[18] MAGURRAN A E. Measuring Biological Diversity. Oxford:Blackwell Publishing, 2004: 256.
[19] HOULAHAN J E, KEDDY P A, MAKKAY K, FINDLAY C S. The effects of adjacent land use on wetland species richness and community composition. Wetlands, 2006, 26(1): 79-96.
[20] LEPŠ J, ŠMILAUER P. Multivariate Analysis of Ecological Data Using CANOCO. Cambridge, UK: Cambridge University Press, 2003. [21] OKSANEN J, KINDT R, LEGENDRE P, O’HARA B, STEVENS M H H, OKSANEN M J, SUGGESTS M A. Vegan: community ecology package R package version 2.2-1[OL]. [2016-02-02]. http://vegan.rforge.r- project.org/.
[22] 许洪军, 于立忠, 黄选瑞, 朱教君, 杨晋宇, 高尚林, 王拥军. 辽东山区次生林与人工林大型地表节肢动物多样性. 生态学杂志, 2015,34(3): 727-735. XU H J, YU L Z, HUANG X R, ZHU J J, YANG J Y, GAO S L,WANG Y J. Biodiversity of macro ground-dwelling arthropods in secondary forests and plantation forests of montane region of eastern Liaoning Province. Chinese Journal of Ecology, 2015, 34(3): 727-735. (in Chinese)
[23] MARTIN E A, REINEKING B, SEO B, STEFFAN-DEWENTER I. Natural enemy interactions constrain pest control in complex agricultural landscapes. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(14): 5534-5539.
[24] 陈学新, 任顺祥, 张帆, 彩万志, 曾凡荣, 张文庆. 天敌昆虫控害机制与可持续利用. 应用昆虫学报, 2013, 50(1): 9-18. CHEN X X, REN S X, ZHANG F, CAI W Z, ZENG F R, ZHANG W Q. Mechanism of pest management by natural enemies and their sustainable utilization. Chinese Journal of Applied Entomology, 2013,50(1): 9-18. (in Chinese)
[25] REBEK E J, SADOF C S, HANKS L M. Manipulating the abundance of natural enemies in ornamental landscapes with floral resource plants. Biological Control, 2005, 33: 203-216.
[26] 汪庚伟, 田俊策, 朱平阳, 郑许松, 徐红星, 杨亚军. 蜜源食物对节肢动物天敌寿命、繁殖力和控害能力的影响. 昆虫学报, 2014,57(8): 979-990. WANG G W, TIAN J C, ZHU P Y, ZHENG X S, XU H X, YANG Y J. Effects of sugar-rich foods on the longevity, fecundity and pest control capacity of arthropod natural enemies. Acta Entomologica Sinica, 2014, 57(8): 979-990. (in Chinese)
[27] TYLIANAKIS J M, TSCHARNTKE T, LEWIS O T. Habitat modification alters the structure of tropical host-parasitoid food webs. Nature, 2007, 445(7124): 202-205.
[28] PAROLIN P, BRESCH C, PONCET C, DESNEUX N. Functional characteristics of secondary plants for increased pest management. International Journal of Pest Management, 2012, 58(4): 369-377.
[29] 尤民生, 刘雨芳, 侯有明. 农田生物多样性与害虫综合治理. 生态学报, 2004, 24(1): 117-122. YOU M S, LIU Y F, HOU Y M. Biodiversity and integrated pest management in agroecosystems. Acta Ecologica Sinica, 2004, 24(1):117-122. (in Chinese)
[30] GRIFFITHS G J, HOLLAND J M, BAILEY A, THOMAS M B. Efficacy and economics of shelter habitats for conservation biological control. Biological Control, 2008, 45(2): 200-209.
[31] KAJITA Y, EVANS E W. Alfalfa fields promote high reproductive rate of an invasive predatory lady beetle. Biological Invasions, 2010,12(7): 2293-2302.
[32] 雒珺瑜, 张帅, 王春义, 吕丽敏, 李春花, 崔金杰. 不同诱集作物对棉田刺吸性害虫及其天敌的生态作用比较. 中国棉花, 2014,41(8): 14-16. LUO J Y, ZHANG S, WANG C Y, LÜ L M, LI C H, CUI J J. Ecological effects of different trap to sucking pests and natural enemies in cotton fields. China Cotton, 2014, 41(8): 14-16. (in Chinese)
[33] SHELTON A M, BADENES-PEREZ F R. Concepts and applications of trap cropping in pest management. Annual Review of Entomology,2006, 51: 285-308.
[34] JACKSON M M, TURNER M G, PEARSON S M, IVES A R. Seeingthe forest and the trees: multilevel models reveal both species and community patterns. Ecosphere, 2012, 3(9): article 79.
[35] 郑云开, 尤民生. 农业景观生物多样性与害虫生态控制. 生态学报,2009, 29(3): 1508-1518. ZHENG Y K, YOU M S. Biological diversity in support of ecologically-based pest management at landscape level. Acta Ecologica Sinica, 2009, 29(3): 1508-1518. (in Chinese)
[36] 张鑫, 王艳辉, 刘云慧, 戴漂漂, 董杰, 宇振荣. 害虫生物防治的景观调节途径: 原理与方法. 生态与农村环境学报, 2015, 31(5):617-624. ZHANG X, WANG Y H, LIU Y H, DAI P P, DONG J, YU Z R. Approaches biological control of pests of through landscape regulation: theory and practice. Journal of Ecology and Rural Environment, 2015, 31(5): 617-624. (in Chinese)
[37] HOGG B N, BUGG R L, DAANE K M. Attractiveness of common insectary and harvestable floral resources to beneficial insects. Biological Control, 2011, 56(1): 76-84.
[38] BALZAN M V, WÄCKERS F L. Flowers to selectively enhance the fitness of a host-feeding parasitoid: Adult feeding by Tuta absoluta and its parasitoid Necremnus artynes. Biological Control, 2013, 67(1):21-31.
[39] FINKE D L, DENNO R F. Predator diversity and the functioning of ecosystems: the role of intraguild predation in dampening trophic cascades. Ecology Letters, 2005, 8(12): 1299-1306.
[40] STRAUB C S, FINKE D L, SNYDER W E. Are the conservation of natural enemy biodiversity and biological control compatiblegoals?Biological Control, 2008, 45(2): 225-237.
[41] RATNADASS A, FERNANDES P, AVELINO J, HABIB R. Plant species diversity for sustainable management of crop pests and diseases in agroecosystems: a review. Agronomy for Sustainable Development, 2012, 32(1): 273-303.
[42] HUANG N X, ENKEGAARD A, OSBORNE L S, RAMAKERS P M J, MESSELINK G J, PIJNAKKER J, MURPHY G. The banker plant method in biological control. Critical Reviews in Plant Sciences, 2011,30(3): 259-278.
[43] WAN F H, YANG N W. Invasion and management of agricultural alien insects in China. Annual Review of Entomology, 2016, 61:77-98.
(责任编辑 岳梅)
Natural Enemy Diversity on Trapping Crops and Its Application for Control of Aphids in Greenhouse Cucumber
WU Sheng-yong1, XU Li-rong2, LI Ning2, WANG Deng-jie3, LEI Zhong-ren1,4
(1State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing 100193;2Huanggang Academy of Agricultural Sciences, Huanggang 438000, Hubei;3Dachuan Plant Protection and Quarantine Station in Dazhou City of Sichuan Province, Dazhou 635000, Sichuan;4Fujian-Taiwan Joint Centre for Ecological Control of Crop Pests, Fuzhou 350002)
【Objective】Concern over protection and application of natural enemies for biological control of insect pests has increased considerably in recent years. The objective of this study is to assess the relationship between natural enemy diversity on multiple mixed trapping crops and its influence factors. Furthermore, the efficacy of banker plant system for control of the target pest aphids in cucumber greenhouse was evaluated. 【Method】Five species of trapping crops including maize, alfalfa, sunflower, wheatand soybean were mix-planted in the experiment stations of Langfang and Huanggang in 2012, 2103 and 2015, respectively. The species of natural enemies were investigated during the experimental period. The community parameters of natural enemies were calculated, and the relationship between community structure and its influence factors (plant species, sampling time and flowering phase) was analyzed by redundancy analysis. In 2013, the dominant natural enemy, i.e., ladybird beetle was collected from the trap cropping and stored in laboratory. In addition, the wheat aphid and wheat were selected as the alternative prey and banker plant of ladybird beetle with different densities, respectively, and the system for control of Aphis gossypii was introduced on greenhouse cucumber. The control efficacy among different treatments was compared.【Result】Analyses of community parameters of natural enemies, Shannon-Wiener diversity index (H’), Simpson’s diversity index (D) and Pielou’s evenness index (J) showed differences over the sampling time. Aphidius spp. and Propylaea japonica were determined as dominant natural enemies in two years. The abundances of both species accounted for over 10% in all natural species on the mixed trap cropping. Redundancy analysis results indicated that plant species, flowering phase and sampling time significantly affected the abundance of natural enemies, and in total explained 26.02% and 17.20% of the variance of the natural enemy community data in 2012 and 2015, respectively. Graph of RDA in 2012 and 2015 indicated that Orius spp. and Chrysoperla sinica displayed higher correlation with the flowering phase. When introduced the banker plant system with density of 200 ladybird beetles in greenhouse, the density of A. gossypii was significantly lower among the treatments, with a control efficacy of 69.4%. 【Conclusion】Attractiveness of natural enemies was integrated with the strategy of their enhanced abundance by means of artificial assistance. Different species of trapping crop plants were mix-planted,attracted multiple natural enemies groups. The dominant natural enemy attracted in the mixed plants was screened to develop the banker plant system, which was then introduced in greenhouse cucumber for control of aphids. The banker plant system with the highest density of ladybird beetle resulted in a certain reduction in aphids population. Therefore, it is practicable to integrated strategies involving the trapping, protection, screening and application of natural enemies for biological control of insect pests.
trap cropping; natural enemies; redundancy analysis; banker plant system; release and application; aphid
2016-02-02;接受日期:2016-04-22
国家大宗蔬菜产业技术体系(CARS-25-B-07)、国家公益性行业(农业)科研专项(201303019-02)
联系方式:吴圣勇,E-mail:sywu@ippcaas.cn。徐丽荣,E-mail:300200100xlr@163.com。吴圣勇和徐丽荣为同等贡献作者。通信作者雷仲仁,E-mail:leizhr@sina.com
