无齿相手蟹体表刚毛的结构及其功能
2016-09-15章赛赛吕晓露孔祥东
赖 文,陈 岑,叶 婷,章赛赛,吕晓露,孔祥东
(浙江理工大学,a.材料与纺织学院,b.生命科学学院,c.启新学院,杭州310018)
无齿相手蟹体表刚毛的结构及其功能
赖文a,陈岑b,叶婷b,章赛赛c,吕晓露c,孔祥东b
(浙江理工大学,a.材料与纺织学院,b.生命科学学院,c.启新学院,杭州310018)
对源自钱塘江的无齿相手蟹体表刚毛的结构及其功能进行研究。利用场发射扫描电子显微镜(FE-SEM)对无齿相手蟹足部、颊部、脐部、螯部刚毛结构与形貌进行观察,并对其不同部位刚毛的结构及其对应的功能进行了分析。通过红外吸收光谱(FT-IR)和X射线粉末衍射(XRD)对无齿相手蟹刚毛的成分和晶相进行表征,结果表明无齿相手蟹刚毛的主要成分均为无定型结构的甲壳素,但其形态结构差异显著,这与刚毛的感知、防御、清洁等作用密切关联。
无齿相手蟹;刚毛;结构;功能
0 引 言
钱塘江流域湿地生态系统中生存着多种蟹类,经实地考察,发现其中包括无齿相手蟹、红螯相手蟹、中华绒螯蟹、平背蜞、弧边招潮蟹等不同种类的螃蟹。其中无齿相手蟹种群数量最高、分布最广,是最主要的蟹类。无齿相手蟹属于动物界、节肢动物门、甲壳动物亚门、软甲纲、软甲亚纲、十足目、爬行亚目、方蟹总科、相手蟹科、相手蟹属,从北到南主要分布在辽东半岛、江苏、浙江、福建、台湾、广东等地[1]。在各沿海河流的湿地泥岸,无齿相手蟹大部分穴居于河流泥滩、田埂、石块间、土壁洞穴或栖息于芦苇沼泽地[2-3],主要摄食底栖藻类、有机碎屑、软泥、芦苇等湿地植物的叶片等。无齿相手蟹是入海河口广泛分布的优势底栖动物,与其它底栖动物、滩涂植被之间生态关系密切,在整个湿地生态系统中发挥着重要的生态平衡作用[4-6]。
许多动物体表都长有形态各异、功能多样的刚毛,不同结构的刚毛具备不同的功能。例如壁虎可在光滑的墙壁、天花板上灵活捕食,是缘于壁虎脚掌底部刚毛具有较强的粘附作用[7-8],这种粘附作用来源于其脚掌底部数百万根微米级刚毛和纳米级绒毛阵列与接触表面接触产生的范德华力[9-10]。蜘蛛能在墙壁等光滑表面上自由移动是因为其足底细密刚毛群尖端呈三角膨大,能够扩大脚底刚毛与墙壁的接触面积从而产生极强的粘附力[11-12]。蜣螂能够生存在粘湿土壤环境中是缘于其腹面和足部刚毛方向与土壤表面垂直而形成柔性界面,通过减小两者之间接触面积使得刚毛具有优良的抗粘附功能和脱附作用[13-14]。蟑螂在遇到外界刺激逃离反应非常迅速,因为其腹部尾须刚毛连接着触觉系统而具有高度灵敏的感觉功能[15]。水黾能在波动的水面上稳定滑行是因为其腿部数千根微米级刚毛同向排列构成螺旋状的沟槽,空气被吸附在沟槽中形成空气垫,阻碍了水滴的浸润使得刚毛具有超强疏水性能[16-17]。
无齿相手蟹作为钱塘江水域湿地生态系统中重要优势物种之一,有关其刚毛结构与功能的研究目前尚未见报道。本文通过对钱塘江无齿相手蟹足部、颊部、脐部、螯部刚毛形貌及其结构的研究,来探析无齿相手蟹不同部位刚毛的结构及其功能。
1 材料与表征方法
1.1实验材料的选取
无齿相手蟹在桐庐富春江水库大坝以下至杭州湾跨海大桥一带150 km的钱塘江流域湿地内均有分布,其中从杭州六和塔附近至嘉兴海宁一带湿地中种群密度最大。课题组于杭州下沙钱塘江江东大桥泄洪水道两侧的湿地中捕获健壮、成年无齿相手蟹若干只。利用超薄刀片将无齿相手蟹足部、颊部、脐部、螯部刚毛切下,用去离子水、无水乙醇清洗干净并冷冻干燥。取无齿相手蟹体表刚毛进行FE-SEM、XRD、FT-IR材料表征以及结构与功能的研究分析。
1.2场发射扫描电镜(FE-SEM)观察
将各部位刚毛粘附在导电胶上。采用FE-SEM(ZEISS-ULTRA55,加速电压为3.0kV)对无齿相手蟹足部、颊部、脐部、螯部刚毛的结构进行观察。其中,为了研究无齿相手蟹刚毛横截面的结构,选取足部刚毛进行环氧树脂包埋,经超薄切片机切取横截面后进行电镜观察。
1.3X射线粉末衍射仪(XRD)观察
将冷冻干燥后的无齿相手蟹足部刚毛研磨成粉末状,采用X-射线粉末衍射仪(ARL-X’TRA,管电压40 kV,管电流35 mA,扫描速度为5 °/min,扫描范围2θ=10°~55°)对足部刚毛进行物相和结晶性能分析。
1.4红外吸收光谱仪(FT-IR)观察
采用KBr压片法,将冷冻干燥后研磨成粉末的无齿相手蟹足部刚毛与溴化钾混合,质量比约为1∶100,充分研磨后将混合物进行压片(30MPa,60s)。利用傅立叶变换红外光谱仪(Nicolet 5700, Thermo Electron,扫描范围为4000~400 cm-1)对样品进行分析。
2 实验结果与分析
2.1无齿相手蟹体貌的观察
钱塘江流域内的无齿相手蟹生长期主要分为蚤状幼体、大眼幼体、幼蟹和成蟹等几个阶段。无齿相手蟹的生长过程中要经历多次蜕皮过程,每次蜕皮过程后它的体重和形态都会增加和变大。图1为成年无齿相手蟹体貌外观图,从外观特征上可以明显看出,成年无齿相手蟹足部均匀分布长约0.2~1 cm的刚毛。无齿相手蟹作为土壤穴居动物,非光滑表面的螯足对土壤具有反粘附能力[18],它们利用螯足在河流泥滩中挖掘洞穴或取食时不会粘附上泥土。

图1 无齿相手蟹体貌外观图
如图2所示,无齿相手蟹有坚硬的四方形甲壳。头胸甲呈方形或梯形,呈茶褐色。额宽大,大于头胸甲宽度的1/2,前缘中部有较宽的凹陷,额后部有4个并立的突起,侧缘具光滑隆线。眼具短柄,呈土青色,可活动;外眼窝齿呈三角形,背眼窝缘光滑且凹陷。螯足1对,步足4对,步足长节、腕节、掌节、指节密生长短不等的硬刚毛。长节背缘近末端处具一刺;掌节外侧面具鳞形颗粒,背缘具1条颗粒隆线。雌雄异形,雄性脐部呈三角形,雌性脐部呈圆形,成年雄性个体显著大于雌性。
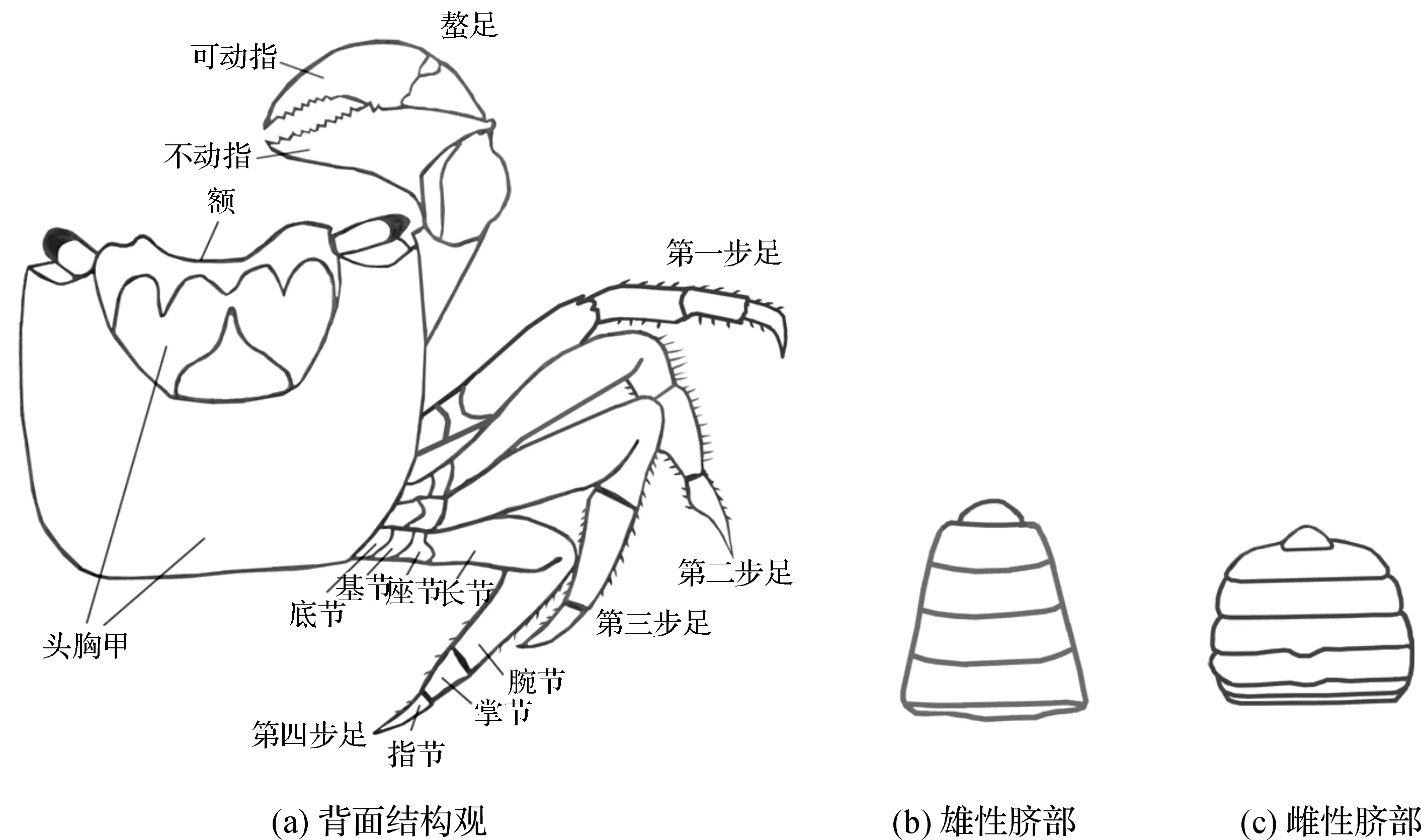
图2 无齿相手蟹形态结构示意图
2.2不同部位刚毛的场发射扫描电镜表征结果
图3(a)为成年无齿相手蟹足部刚毛在视野放大500倍的场发射扫描电镜图。通过观察,足部刚毛呈纤维状结构,刚毛表面带有排列整齐的球状突起结构,刚毛直径由底端至末端逐渐变小,长度为0.2~1 cm,直径为20~30 μm。图3(b)为成年无齿相手蟹足部刚毛在视野放大5 000倍的场发射扫描电镜图,刚毛表面球状突起结构类似于半球体,直径约为15 μm。Garm等[19]曾报道青蟹第二、第三颚足末端具有羽毛状的刚毛,上含有机械性刺激感受器,捕食时刚毛与猎物相互接触后可感知猎物相关的信息。无齿相手蟹这种球状突起结构极有可能与触觉系统具有重要关联,对其感知水流速度或方向、水波振动、获知猎物或危险信号等方面起到相关作用。

图3 无齿相手蟹足部刚毛的FE-SEM图
图4(a)为成年无齿相手蟹足部刚毛横截面在视野放大800倍的场发射扫描电镜图,图中足部刚毛横截面外端包覆层为环氧树脂包埋剂。足部刚毛横截面为中空不规则圆形环状结构,圆形环状结构外径约为80 μm,内径约为30 μm。图4(b)为成年无齿相手蟹足部刚毛横截面在视野放大3 000倍的场发射扫描电镜图,其中足部刚毛壁厚约为25 μm,内径表面非光滑。Wolcott等[20]曾报道了沙蟹的第二步足和第三步足的毛细管刚毛簇能够从土壤中收集大量水分运输至鳃室以维持体内水平衡。类似的,无齿相手蟹刚毛的中空环状结构,对水分与营养物质的运输及吸收是有利的。此外,无齿相手蟹在蜕壳以后,新生刚毛作为体表附属物能够从原刚毛中“金蝉脱壳”,此过程的完成与刚毛所具有的中空环状结构密切关联。

图4 无齿相手蟹足部刚毛横截面结构FE-SEM图
图5(a)-(d)分别为成年无齿相手蟹颊部底端刚毛在视野放大200、500、2 000、6 000倍的场发射扫描电镜图。无齿相手蟹颊部底端刚毛呈纤维状结构,刚毛直径由底端至末端逐渐变小。刚毛长度为2~3 mm,直径为20~25 μm。颊部底端刚毛之间排列整齐且密集,间隔均匀,方向一致。颊部底端刚毛上长有分支刚毛,分支刚毛之间排列密集、方向规则,长度为30~60 μm,直径为2~4 μm。颊部底端刚毛与分支刚毛共同组成紧密的网状结构,这种结构有助于阻挡水中微小颗粒物,防止其透过颊部与胸部交界处进入胸腔,造成危害。颊部底端的刚毛具有网状分支结构,考虑到其位置,这类刚毛可能在减少水阻、保持颊部甲壳湿润等方面具有特殊功能。

图5 无齿相手蟹颊部刚毛的FE-SEM图
图6(a)、(b)分别为成年无齿相手蟹脐部边缘刚毛在视野放大500、2 000倍的场发射扫面电镜图。无齿相手蟹脐部刚毛呈纤维状结构,刚毛直径由底端至末端逐渐变小。刚毛长度为200~1 000 μm,直径为10~15 μm。刚毛之间排列紧密,间隔均匀。脐部刚毛上长有分支刚毛,长度为30~60 μm,直径为2~4 μm,上面粘有大量细小颗粒附着物。脐部刚毛及其分支刚毛所共同组成的致密结构,可有效吸附或阻挡水中颗粒物质进入脐内。

图6 无齿相手蟹脐部刚毛的FE-SEM图
图7(a)、(b)分别为成年无齿相手蟹螯部长节刚毛在视野放大500、1 000倍的场发射扫面电镜图。如图7所示,螯部长节刚毛呈现出管状和锯齿状两种典型纤维状结构,刚毛长度为1000~2000 μm,直径为30~60 μm。刚毛表面有一层薄壳层且内部有许多小孔洞,刚毛直径由底端至末端逐渐变小。由于螯部长节部位在蟹类休息、洞栖时均与蟹颊部长期接触,螯部长节表面管状和锯齿状两种刚毛的混合存在,尤其是锯齿状的刚毛可有效清洁梳理蟹颊部致密的网状刚毛(如图5所示),从而保持蟹颊部体表刚毛的清洁。

图7 无齿相手蟹螯部刚毛的FE-SEM图
2.3红外吸收光谱分析结果
图8(a)-(d)分别为成年无齿相手蟹脐部、螯部、足部、颊部刚毛的FTIR图。从FTIR谱图中可以看出:3445 cm-1处的峰为缔合—OH伸缩振动吸收峰,2927 cm-1处的峰为-CH3的对称伸缩振动吸收峰,1650 cm-1的峰(酰胺I谱带)和1538 cm-1的峰(酰胺II谱带)分别对应—NHCOCH3的C=O伸缩振动吸收峰和N—H弯曲振动吸收峰,1432 cm-1处为C—H弯曲振动吸收峰,1239cm-1(酰胺III谱带) 处的峰为—NHCOCH3的C—N伸缩振动吸收峰,1158 cm-1处的峰为C—O—C伸缩振动吸收峰, 1071 cm-1、1030 cm-1处的峰分别对应二级醇羟基与一级醇羟基,877 cm-1处的峰是CH环伸缩振动吸收峰。不
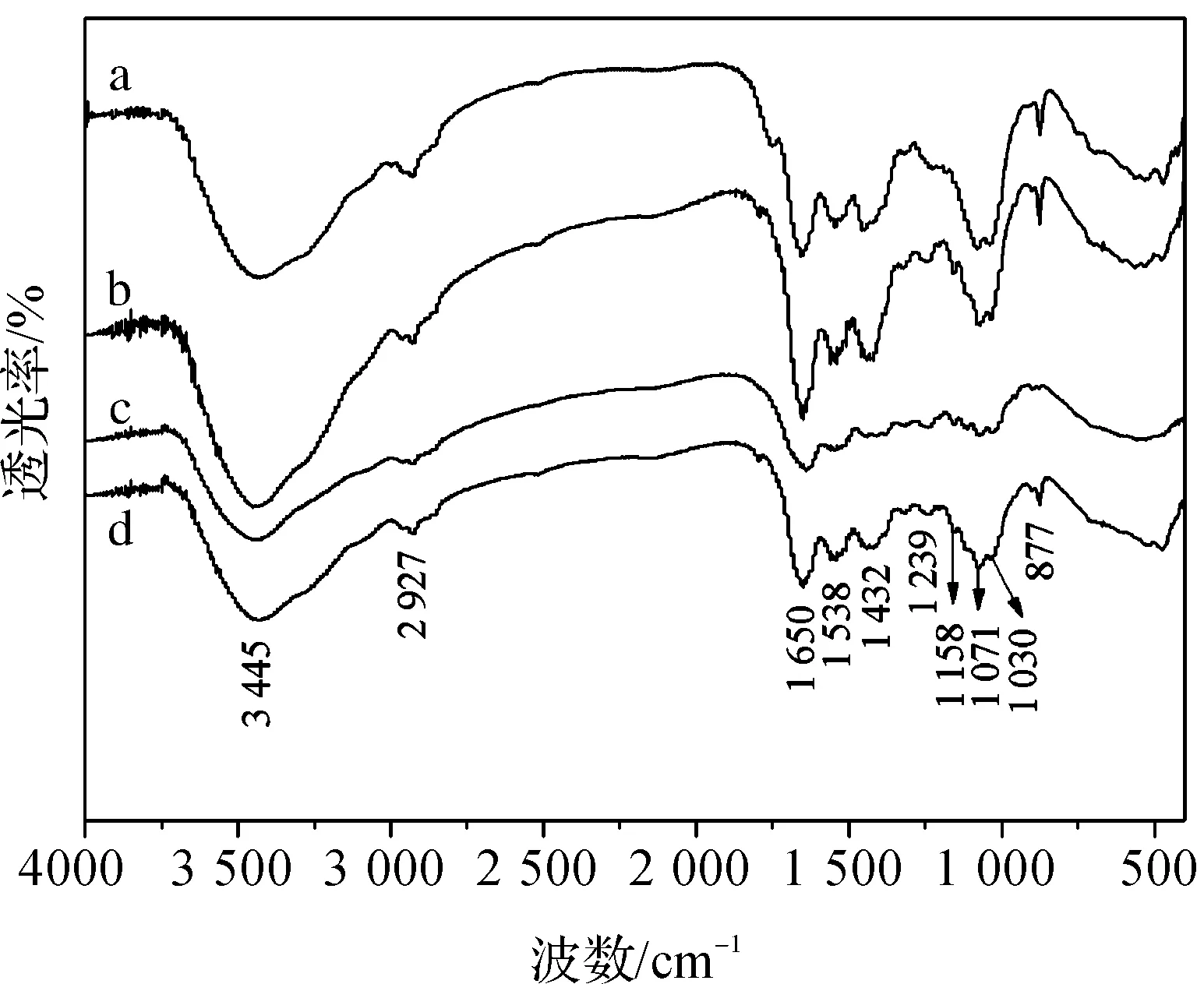
图8 无齿相手蟹不同部位刚毛的FTIR图谱
同刚毛FTIR的峰强有差异,峰的位置相近,显示了典型的甲壳素结构[21-22]。该结果证实无齿相手蟹体表不同部位刚毛的主要成分均是甲壳素。
2.4X射线粉末衍射分析结果
由于无齿相手蟹各个部位刚毛主要成分均为甲壳素,因此选择无齿相手蟹足部刚毛进行X射线粉末衍射分析。图9为成年无齿相手蟹足部刚毛的XRD图,从图中可以看出,在2θ=20°处出现了一个衍射峰,衍射峰强度较小、峰形较宽,该衍射峰显示了刚毛中的甲壳素呈无定型结构,也就是说,无齿相手蟹足部刚毛主要成分为无定型结构的甲壳素。虽然刚毛的主要组成成分接近,但是由于在结构组装上的巨大差异,它们在无齿相手蟹的体表分别发挥着感知、水分运输、防御、清洁等不同的功能。
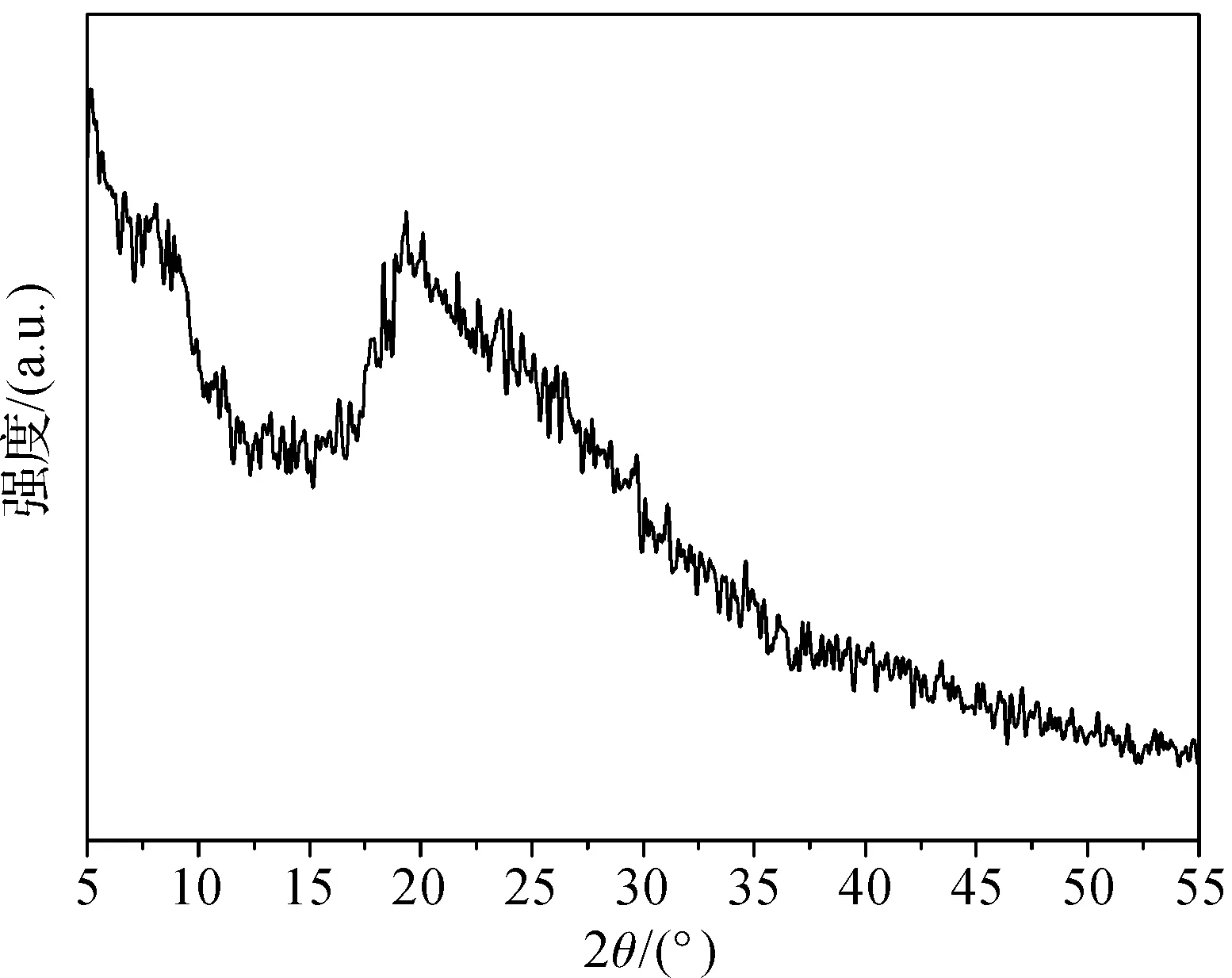
图9 无齿相手蟹足部刚毛的XRD图谱
3 结 论
无齿相手蟹足部刚毛和螯部长节刚毛均为空心结构,足部刚毛表面含有球状突起结构,螯部长节刚毛呈现管状和锯齿状两种典型结构,颊部底端刚毛和脐部刚毛均与其分支刚毛共同构成网状结构。无齿相手蟹足部、颊部、脐部、螯部纤维状刚毛的主要成分均为甲壳素,虽然成分相近,但是由于不同的结构组装及其形态,刚毛分别呈现出感知、水分运输、防御、清洁等不同功能。刚毛的结构、长度、直径各不相同,预示着其具备不同的潜在功能,不同刚毛所具有的功能十分复杂,尚需要进一步的详细研究。无齿相手蟹体表刚毛结构的分级组装受到精确的生物控制,其结构与功能的对应关系对开展材料仿生学研究具有重要的潜在意义。
[1] 戴爱云.中国海洋蟹类[M].北京:海洋出版社,1986:503-506.
[2] 熊李虎,陆健健.长江河口湿地不同植被中无齿螳臂相手蟹的分布及其洞穴利用[J].动物学杂志,2009,44(6):1-9.
[3] XIONG L H, WANG A, LU J J. The validity of burrow counting in estimating the densities of chiromantes dehaani and ilyoplax deschampsi in tidal flat at yangtze estuary[J]. Acta Ecologica Sinica, 2010, 30(1): 40-44.
[4] WANG J Q, ZHANG X D, JIANG L F, et al. Bioturbation of burrowing crabs promotes sediment turnover and carbon and nitrogen movements in an estuarine salt marsh[J]. Ecosystems, 2010, 13(4): 586-599.
[5] WANG J Q, ZHANG X D, NIE M, et al. Exotic spartina alterniflora provides compatible habitats for native estuarine crab sesarma dehaani in the yangtze river estuary[J]. Ecological Engineering, 2008, 34(1): 57-64.
[6] ZHANG X D, JIA X, CHEN Y Y, et al. Crabs mediate interactions between native and invasive salt marsh plants: A mesocosm study[J]. Plos One, 2013, 8(9): e74095-e74095.
[7] TIAN Y, PESIKA N, ZENG H B, et al. Adhesion and friction in gecko toe attachment and detachment[J]. Proceedings of the National Academy of Sciences, 2006, 103(51): 19320-19325.
[8] AUTUMN K, LIANG Y A, HSIEH S T, et al. Adhesive force of a single gecko foot-hair[J]. Nature, 2000, 405(6787): 681-685.
[9] AUTUMN K, SITTI M, LIANG Y A, et al. Evidence for van der waals adhesion in gecko setae[J]. Proceedings of the National Academy of Sciences, 2002, 99(19): 12252-12256.
[10] GAO H J, WANG X, YAO H M, et al. Mechanics of hierarchical adhesion structures of geckos[J]. Mechanics of Materials, 2005, 37(2): 275-285.
[11] KESEL A B, MARTIN A, SEIDL T. Getting a grip on spider attachment: an AFM approach to microstructure adhesion in arthropods[J]. Smart Materials and Structures, 2004, 13(3): 512-518.
[12] KESEL A B, MARTIN A, SEIDI T. Adhesion measurements on the attachment devices of the jumping spider evarcha arcuata[J]. Journal of Experimental Biology, 2003, 206(16): 2733-2738.
[13] 程红, 孙久荣, 李建桥, 等. 臭蜣螂体壁表面结构及其与减粘脱附功能的关系[J]. 昆虫学报, 2002, 45(2): 175-181.
[14] SUN J R, LI J Q, CHENG H, et al. Restudies on body surface of dung beetle and application of its bionics flexible technique[J]. Journal of Bionics Engineering, 2004, 1(1): 53-60.
[15] BOOTH D, MARIE B, DOMENICI P, et al. Transcriptional control of behavior: Engrailed knock-out changes cockroach escape trajectories[J]. The Journal of Neuroscience, 2009, 29(22): 7181-7190.
[16] GOODWYN P P, DE SOUZA E D, FUJISAKI K, et al. Moulding technique demonstrates the contribution of surface geometry to the super-hydrophobic properties of the surface of a water strider[J]. Acta Biomaterialia, 2008, 4(3): 766-770.
[17] GAO X F, JIANG L. Biophysics: water-repellent legs of water striders[J]. Nature, 2004, 432(7013): 36-36.
[18] ZHOU F, WU Z W, WANG M L, et al. Structure and mechanical properties of pincers of lobster (procambarus clarkii) and crab (eriocheir sinensis)[J]. Journal of the Mechanical Behavior of Biomedical Materials, 2010, 3(6): 454-463.
[19] GARM A. Mechanosensory properties of the mouthpart setae of the european shore crab carcinus maenas[J]. Marine Biology, 2005, 147(5): 1179-1190.
[20] WOLCOTT T G. Uptake of interstitial water from soil: Mechanisms and ecological significance in the ghost crab ocypode quadrata and two gecarcinid land crabs[J]. Physiological Zoology, 1984, 57(1): 161-184.
[21] KAYA M, SEYYAR O, BARAN T, et al. A physicochemical characterization of fully acetylated chitin structure isolated from two spider species: With new surface morphology[J]. International Journal of Biological Macromolecules, 2014, 65(1) : 553-558.
[22] KAYA M, ERDOGAN S, MOL A, et al. Comparison of chitin structures isolated from seven orthoptera species[J]. International Journal of Biological Macromolecules, 2015, 72(1) : 797-805.
(责任编辑: 许惠儿)
Structures and Functions of Setae on Crab, Sesarma Dehaani
LAIWena,CHENCenb,YETingb,ZHANGSaisaic,LÜXiaoluc,KONGXiangdongb
(a. College of Materials and Textile, b. College of Life Sciences, c. College of Qinxin, Zhejiang Sci-Tech University, Hangzhou 310018, China)
This paper mainly researched the structures and functions of setae on crabof Sesarma dehaani which was from the Qiantang River. The field emission scanning electron microscope (FE-SEM) was introduced to observe the structure as well as the morphology of different kinds of setae in Sesarma dehaani, including the setae from its foot, genal region, navel and pincers. Besides, structures and functions of different kinds of setae in Sesarma dehaani were analyzed. Besides, the constituents and crystalline phases of the setae of Sesarma dehaani were characterized by infrared absorption spectrum (FT-IR) and X-ray powder diffraction pattern (XRD). Results indicate that the main constituent of the setae of Sesarma dehaani is amorphous chitin. The significant differences in morphological structure are closely related to perception, water transmission, defense and cleaning and other functions of surface setae.
Sesarma dehaani; setae; structure; function
10.3969/j.issn.1673-3851.2016.05.019
2015-08-11
国家自然科学基金项目(50942023,51272236);浙江理工大学521人才培养计划(1610032521302)
赖文(1990-),男,江西赣州人,硕士研究生,主要从事生物材料方面的研究。
孔祥东,E-mail:Kongxiangdong@gmail.com
TS195.644
A
1673- 3851 (2016) 03- 0433- 06 引用页码: 050701
