37个平菇菌株的遗传差异分析
2016-09-01盛春鸽潘春磊王延锋磊王金贺刘姿彤于海洋黑龙江省农业科学院牡丹江分院黑龙江牡丹江5704河北省农林科学院遗传生理研究所河北石家庄05005
盛春鸽 潘春磊 王延锋* 李 慧 史 磊王金贺 刘姿彤 张 鹏 于海洋(. 黑龙江省农业科学院牡丹江分院,黑龙江 牡丹江5704;. 河北省农林科学院遗传生理研究所,河北 石家庄 05005)
37个平菇菌株的遗传差异分析
盛春鸽1潘春磊1王延锋1*李慧2史磊1王金贺1刘姿彤1张鹏1于海洋1
(1. 黑龙江省农业科学院牡丹江分院,黑龙江 牡丹江157041;2. 河北省农林科学院遗传生理研究所,河北 石家庄 050051)
摘要以22个国内平菇菌株,15个外引菌株为试验材料,观测其菌丝长速、子实体颜色、菌盖及菌柄形态等14个农艺性状,对各性状进行多样性及主成分分析,同时对8个性状进行聚类分析。结果表明:不同平菇性状之间存在不同程度的多样性,多样性指数变化范围 0.393~1.961,国内群体的平均多样性指数为 1.611,高于外引材料(1.397)13.28%。各性状间变异系数范围4%~75%,这两个极端值均出现在外引群体中。多变量的主成分分析,14个性状,可以用7个主成分来概括,7个主成分的方差累积贡献率为84.44%。各因子所代表的指标总结为单菇因子、发育因子和产量因子3个因素。8个性状的聚类分析把37份材料分为3个类群,分类结果与子实体农艺性状有一定的相关性。
关键词平菇;性状;种质资源;多样性
我国平菇资源较为丰富[1],全国大部分省市都有栽培,以河南、山东栽培量最大。吴素蕊等统计2007—2011年五年的总产量为2 414.676万吨,是我国出口创汇的主要食用菌之一[2,3]。在我国,俗称平菇的食用菌包括糙皮侧耳(Pleurotus ostreatus)、白黄侧耳(P. cornucopiae)、佛罗里达侧耳(P. florida)和肺形侧耳(P. pulmonarius)4个种[4]。有关平菇的生理生化、分子方面的研究特别多,而完全基于表型性状的研究则不多。本研究选取目前国内栽培量较大的典型菌株 22个,从韩国、俄罗斯等引进菌株15个,总计37个菌株,在黑龙江省农业科学院牡丹江分院食用菌研究中心太阳能温室内进行田间栽培,观察鉴定其形态的多样性。
1 材料与方法
1.1试验材料
供试材料,平406、黑平1号、泰平19、ccef89、豫平5号、新科101、双抗黑平、灰美2号,来源于中国农业科学院;林特平来源于黑龙江省林副特产研究所;黑平19、平(新疆-阿勒泰)、美味侧耳2、海丰灰平、黑平2号、PF1、PF2、平109、平425、平838、平杂3、平2026、平150来源于黑龙江省农业科学院牡丹江分院;俄2014 平1、俄2014平2来源于俄罗斯科学院远东分院;韩CB1、韩CB2、韩CB3、韩CB4、韩CB5、韩CB43、韩CB54、韩CB67、韩CB68、韩CB69、韩CB100、韩CB101、韩CBS06来源于韩国忠清北道农业技术院。笔者首先对 37个菌株进行拮抗试验,确认其分属于不同的菌株。
1.2试验方法
(1)供试培养基。马铃薯综合培养基(PDA培养基):去皮马铃薯200 g煮汁,葡萄糖20 g,磷酸二氢钾2 g,硫酸镁0.5 g,琼脂15 g,加蒸馏水定容至1 000 mL。栽培种培养基:木屑78%,麦麸10%,稻糠10%,石灰1%,石膏1%,含水量65%。
(2)试验设计与栽培场所。试验于2014—2015两年春季的4~6月在牡丹江农科院食用菌中心完成,采用随机区组设计,每个菌株3次重复,每个重复40袋。栽培场所气温14~18 ℃,空气湿度80%~95%,光照200~400 lx,通风良好。
(3)农艺性状测定。供试菌株农艺性状指标和测定方法参考李慧等使用的糙皮侧耳新品种DUS测试指南中的部分指标和方法[5]。详见表1。

表1 平菇14个表型性状的测定方法
(4)数据处理和统计分析。对子实体采收期颜色、菌柄形态、菌柄质地、菌盖质地等4个非数值性状按照表2所示进行分级赋值后再做数据分析,对其他10个数值性状进行质量化处理,依照王述民描述的方法进行统计,分为10级,1 级Xi<<—2σ,10级Xi><+2σ,中间每级间隔0.5σ[6]。使用DPS7.05软件进行聚类及主成分分析,聚类时种质间距离采用欧氏距离,类平均(UPGMA)法。多样性指数的计算采用Shannon-Weaver信息指数。

表2 平菇农艺性状基本统计分析
2 结果与讨论
2.1平菇农艺性状基本参数统计分析
37个菌株的农艺性状研究结果(表 2),其14个农艺性状在不同菌株间存在不同程度的遗传变异,同一性状在两个群体上的变异系数和遗传多样性指数也有所不同,各性状间的变异系数范围为4%~75%。这两个极端值均出现在外引群体中,表明外引群体的变异程度大。遗传多样性指数范围为 0.393~1.961。两群体间各性状平均值的T测验结果:国内菌株和外引菌株在菌丝长速和菌柄直径两个性状中存在极显著差异(p<0.01)。国内群体的平均多样性指数为 1.611,高于外引材料(1.397)13.28%。
综合两群体的数据可见,变异系数最大的性状为菌柄形态,最小的为菌丝长速。多样性指数由高到低依次是菌丝长速(1.961)>产量、菌盖短径(1.945)>菌盖长径(1.925)>发菌速度(1.909)>菌柄直径(1.859)。
2.2农艺性状聚类分析
在调查的 14个农艺性状中,笔者选取出菇期间子实体采收期颜色、菌柄形态、菌盖长径、菌盖短径、菌盖长宽比、菌盖厚度、菌柄长度、菌柄直径等8个农艺性状作为基本依据,进行聚类分析,结果如图1。
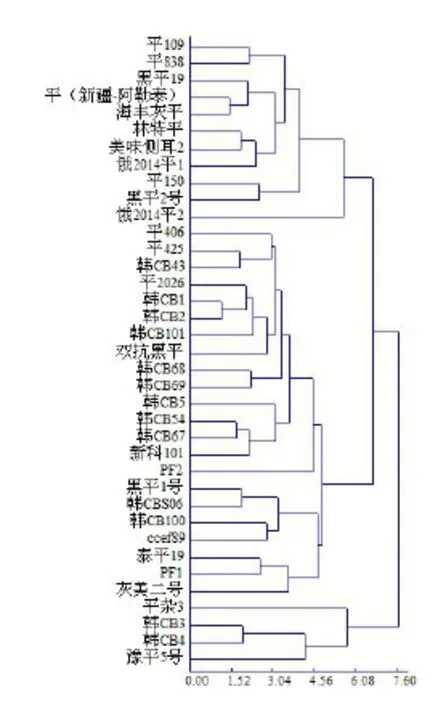
图1 37个平菇种质农艺性状树状图
从图1可以看出,供试菌株在欧氏距离6.08处,可以分为三大类,第一类包括“平109”在内的 11个个体,结合子实体形态观察,第一类子实体的共同特点是菌盖颜色较浅,菌盖大小适中,菌肉较薄,个别品种整齐度较差。第二类个体数目比较多,有 22个菌株,包括大部分的韩引菌株,比照第一类菌株,此类菌株子实体颜色深,菌盖大,质软,肉质肥厚,子实体发生量大且出菇整齐。第三类个体数较少,包括“平杂 3”等4个菌株,除“韩CB4”外,菇体颜色浅,菌盖小。在同一栽培条件下,外引菌株和国内菌株在形态上没有明显差别。
2.337个菌株的主成分分析
对37个菌株14个农艺性状的主成分分析结果(表3)显示,在新得的14个主成分中,所有性状指标提供的信息主要集中在前7个主成分,累计贡献率达84.44%,表明这7个主成分最大程度地反映了本研究中的各项指标。从方差的贡献率来看,前3个主成分的方差贡献率较高,都大于10%。第一个主成分的贡献率达到30%,涵盖了近1/3的信息总量。综合因子在某一性状上的载荷值,以及此载荷值在其他各因子中的大小分析可知:第一主成分主要包括菌盖长径、菌盖短径、菌盖长宽比、菌盖厚度等性状,第二主成分为颜色性状,第三主成分包括菌丝长速、发菌速度性状,第四主成分包括菌柄形态、菌柄质地性状,第五主成分包括菌盖质地、菌龄性状,第六主成分为菌柄质地性状,第七主成分包括菌柄长度、产量性状。重新归纳整理这7个主成分,可得如下三类,第一类包括第1、2、4、5、6主成分,此类主成分反映的性状有菌盖长径、菌盖短径、菌盖长宽比、菌盖厚度、子实体采收期颜色、菌柄形态、菌柄质地、菌盖质地等与单菇相关的性状指标,称之为“单菇因子”[7]。第二类为第 3个主成分,含菌丝长速、发菌速度性状,这些指标与发育期间的营养生长有关,可谓“发育因子”。第三类为第7个主成分,反映的是产量性状,可称为“产量因子”。
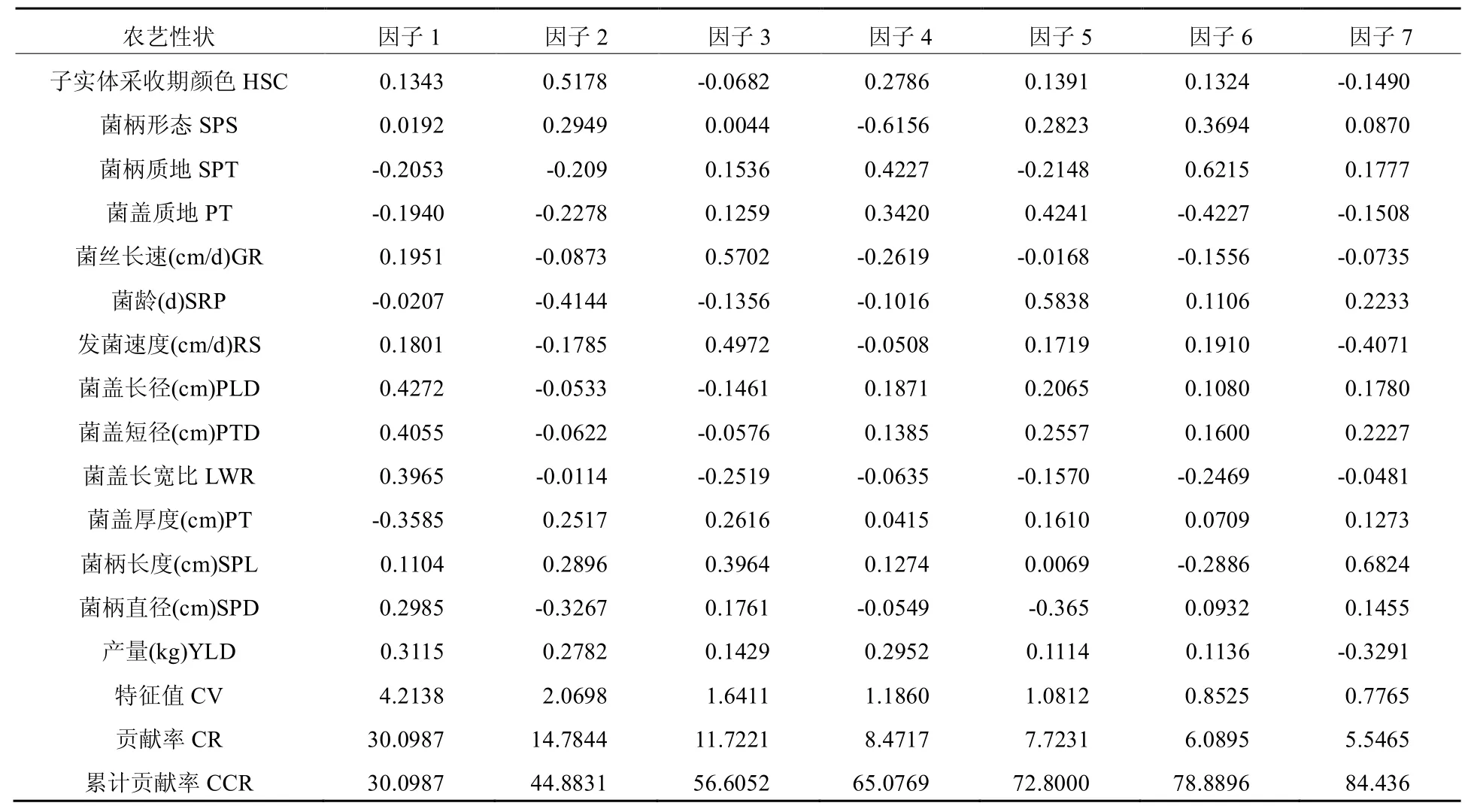
表3 37个菌株的主成分分析
3 讨 论
种质资源多样性一直是遗传学研究的重点,食用菌上有关遗传多样性的研究多集中在分子标记上[8,9]。以形态学特征为基础的现代分类学研究在植物上得到了广泛应用[10~12],管志勇等以切花菊为材料,利用 DUS性状对其相似性和亲缘关系进行分析,取得了较好的结果[13]。张向前等对燕麦的农艺性状进行分析,为燕麦种质资源的研究及燕麦育种打下基础[14]。国外在李[15]、黑加仑[16]、甜瓜[17]上也有类似研究,国内食用菌基于农艺性状做遗传多样性分析的研究比较少,林范学对香菇的 16个数量性状进行表型、遗传和环境相关性分析及主成分分析,表明各个性状和指标之间存在一定程度的相互关系,16个性状缩减为酶活、发育、转色等6个主成分[18]。常艳等对16个糙皮侧耳菌株的10个数量性状进行主成分及聚类分析,聚类结果与农艺性状有一定的联系[19]。此外,陈影等对木耳进行以农艺性状为主的多样性分析[20]。国外在食用菌上的相关研究鲜有报道。
本研究37个菌株中,15株材料为外引材料(引自韩国、俄罗斯),两群体各性状间除菌丝长速和菌柄直径外,其他指标虽略有差异但不明显。外引菌株的变异系数变化范围大于国内菌株,表明外引资源同一性状内存在较大的遗传变异,是良好的育种材料。但遗传多样性指数低于国内菌株,这可能与外引样本量少有一定的关系。从聚类情况看,外引菌株和国内菌株没有完全分开,结果与数值分析相吻合。表明平菇菌株的表型性状与地域分布没有太密切的联系。在韩国,平菇多用850 mL的菌瓶实行工厂化栽培,采收时菌盖直径只有2~3 cm,菌柄几近中生,普遍较长(7 cm左右),便于统一包装。按照我们国内的栽培模式,在改变韩国平菇的栽培管理方式之后,实际栽培鉴定发现:韩引平菇与国内菌株在外观形态上无太大差别。可见,平菇的形态建成与外界条件和栽培管理模式有很大关系,此观点同样适用于金针菇[21,22]。韩国平菇在当地之所以表现不同主要是人工选择的结果,这种选择还未达成平菇种性的变异。所以在对平菇的种性界定上要严格考虑栽培环境(光、温、水、气)和栽培方式(瓶栽、袋栽)等对平菇表型的影响。
本文共调查了37个菌株的14个农艺性状,在聚类分析时作者选择了与子实体形态相关的 8个性状,主要以子实体的宏观形态(菌盖、菌柄等)为基础进行聚类分析,排除菌丝长速、发菌速度等其他性状对聚类效果的干扰,得到的聚类结果与子实体性状有一定程度的相关性,聚类结果比较可靠。
参考文献
[1]图力古尔,李玉. 我国侧耳属真菌的种类资源及其生态地理分布[J]. 中国食用菌,2001,20(5): 8-10.
[2]吴素蕊,赵春艳,侯波,等. 近5年我国食用菌生产区域布局情况分析[J]. 中国食用菌,2013,32(1): 51-53.
[3]关景奎. 小蘑菇成为我国创汇大产业[J]. 农产品加工(创新版),2011,2: 15-16.
[4]张金霞,黄晨阳,郑素月. 平菇新品种-秀珍菇的特征特性[J]. 中国食用菌,2005,24(4): 26-25.
[5]李慧. 我国栽培平菇种质资源评价及核心种质构建[D].福州: 福建农林大学,2012: 25-26.
[6]王述民,曹永生,Redden RJ,等. 我国小豆种质资源形态多样性鉴定与分类研究[J]. 作物学报,2002,28(6): 727-733.
[7]林范学,程水明,潘迎捷. 香菇数量性状的因子分析[J].菌物学报,2004,23(4): 502-507.
[8]宫志远,任海霞,姚强,等. 35个山东主栽平菇菌株的ISSR遗传差异分析[J]. 基因组学与应用生物学,2010,29(3): 507-512.
[9]王卓仁,刘启燕,边银丙,等. 香菇单孢杂交子代群体灰色关联度和 ISSR分析[J]. 菌物学报,2010,29(2): 267-272.
[10]Podgornika M,Vuk I,Vrhovnik I,et al. A survey and morphological evaluation of fig (Ficus carica L.) genetic resources from Slovenia. Scientia Horticulturae,2010,125: 380-389.
[11]王育青,秦艳,王晓晶,等. 内蒙古野生马蔺种质农艺性状遗传多样性研究[J]. 植物遗传资源学报,2014,15(4): 772-778.
[12]冯国郡,李宏琪,叶凯,等. 甜高粱种质资源在新疆的多样性表现及聚类分析[J]. 植物遗传资源学报,2012,13(3): 398-405.
[13]管志勇,王江民,陈发棣,等. 基于DUS测试性状的切花菊品种亲缘关系研究[J]. 园艺学报,2013,40(7): 1399-1406.
[14]张向前,刘景辉,齐冰洁,等. 燕麦种质资源主要农艺性状的遗传多样性分析[J]. 植物遗传资源学报,2010,11(2): 168-174.
[15]Horvath A,Balsemin E,Barbot J C,et al. Phenotypic variability and genetic structure in plum (Prunus domestica L.),cherry plum (P. cerasifera Ehrh.) and sloe (P. spinosa L.)[J]. Scientia Horticulturae,2011,129: 283-293.
[16]Pluta S,Madry W,Sieczko L. Phenotypic diversity for agronomic traits in a collection of blackcurrant (Ribes nigrum L.) cultivars evaluated in Poland[J]. Scientia Horticulturae,2012,145: 136-144.
[17]Szamosi C,Solmaz I,Sari N,et al. Morphological evaluation and comparison of Hungarian and Turkish melon (Cucumis melo L.)germplasm[J]. Scientia Horticulturae,2010,124: 170-182.
[18]林范学,程水明,李安政,等. 香菇数量性状的相关性分析和主成分分析[J]. 菌物学报,2006,25(4): 579-586.
[19]常艳,于娟娟,韩芹芹,等. 糙皮侧耳品种的主成分及聚类分析[J]. 食用菌学报,2011,18(3): 12-16.
[20]陈影,姚方杰,张友民,等. 木耳栽培种质资源的数量分类研究[J]. 菌物学报,2014,33(5): 984-996.
[21]佟希丹. 不同光质LED对黄白品种金针菇商品性状与产量的影响[D]. 长春: 吉林农业大学,2012.
[22]Yuichi Sakamoto. Protein expression during Flammulina velutipes fruiting body formation[J]. Mycoscience,2010(51): 163-169.
*为通讯作者,E-mail:mdjnks@126.com
中图分类号:S646
文献标识码:A
文章编号:2095-0934(2016)04-247-06
基金项目:国家食用菌产业技术体系(CARS-24)、公益性行业(农业)科研专项经费项目(201303080)、黑龙江省应用技术研究与开发计划项目(WB13B10602)
作者简介:盛春鸽(1986—),女,硕士,研究实习员,研究方向为食用菌遗传育种。E-mail:shengchunge@163.com
Analysis on genetic differences of 37 Pleurotus ostreatus strains
Sheng Chunge1Pan Chunlei1Wang Yanfeng1Li Hui2Shi Lei1Wang Jinhe1Liu Zitong1Zhang Peng1Yu Haiyang1
(1. Mudanjiang branch of Heilongjiang Academy of Agricultural Sciences,Mudanjiang,Heilongjiang 157041,China;2. Institute of Genetics and Physiology of Hebei Academy of Agriculture and Forestry Sciences,Shijiazhuang,Hebei 050051,China)
AbstractThis paper presents an assessment of phenotypic diversity for 37 strains of Pleurotus ostreatus. The data of 14 traits describing mycelial growth,fruitbody color,pileus and stipe morphology ect. were recorded. Diversity analysis and principal component analysis (PCA) of 14 traits,cluster analysis (CA) of 8 traits were conducted for learning its biological characteristics on 37 strains. It can also provide the basis for germplasm resource research. The results show that: the characters of different oyster mushroom showed different degree of diversity,which range of 0.393~1.961. The average diversity index(1.611) in domestic germplasm was 13.28% higher than that of introduced group(1.397). coefficient of variation(CV) range of 4%~75% occurred in introduced group,which showed that cited group has a great variation. In addition,7 principal components (PCs) were obtained from 14 tested traits through the principal component analysis (PCA),and the percentage of cumulative contributive rate is 84.44%,these PCs were designated as single fruitbody,development factors,yield respectively. 37strains were classified into 3 sections according to the clustering analysis (CA) of 8 traits,which have a certain correlation with fruitbody agronomic traits.
Key wordsPleurotus ostreatus; traits; germplasm resources; diversity
