粤东6种菊科植物种子萌发的生物学零度和积温
2016-09-01陈文
陈 文
(韩山师范学院地理学系,广东潮州 521041)
粤东6种菊科植物种子萌发的生物学零度和积温
陈文
(韩山师范学院地理学系,广东潮州521041)
温度是影响种子萌发的重要环境因子.该文以积温模型分析了6种菊科植物包括小蓬草(Conyzacanadensis)、鬼针草(Bidenspilosa)、金腰箭(Synedrellanodiflora)、假臭草(Praxelisclematidea)、藿香蓟(Ageratumconyzoides)和金钮扣(Spilanthespaniculata)种子萌发对温度(15,20,25,30,35和40 ℃)的响应以及萌发所需的生物学零度和有效积温.结果表明除小蓬草外(52~55%),其他5种菊科植物种子在适宜温度范围内萌发率达到80~95%;小蓬草和假臭草种子萌发的最适温度范围较窄(20~25 ℃),鬼针草、金钮扣和藿香蓟种子萌发的最适温度范围较宽(15~30 ℃/35 ℃/40 ℃),金腰箭种子萌发的最适温度偏高(25~40 ℃);产于北美洲的小蓬草和分布广泛的鬼针草种子萌发的生物学零度较低(分别为3.94 ℃和5.48 ℃),产于热带美洲或南美洲的金腰箭、假臭草、藿香蓟其生物学零度较高(分别为16.90 ℃、12.56 ℃和11.12 ℃);6种植物种子萌发的生物学零度与原产地年均低温呈显著正相关,与积温呈显著负相关.除了金腰箭外,其余5种菊科植物均具有向更高纬度地区扩张的潜力.
菊科植物;种子萌发;生物学零度;积温;适应性
中国是生态系统高度多样化的国家,也是生物入侵较严重的国家之一,约有515 种外来入侵植物,其中260种出现在我国东南地区[1],并由东南向西北方向逐渐扩散蔓延[2],对当地生态系统和物种多样性造成了严重的影响.揭示外来植物的入侵机制及其预测和防控成为当今入侵生物学研究的核心课题[3-4].种子萌发是植物实现种群更新和物种延续的关键环节,也是生活史中最脆弱且敏感的阶段,易受到各种环境因子的影响[5-6],其中温度是影响种子萌发和幼苗生长的重要环境因子[7-8].每一种植物都需要温度达到一定值时才开始发育和生长,称为发育阈温度或生物学零度(Biological zero)[9],同时,还需要一定的总热量即总积温或者有效积温(Effective accumulated temperature)才能完成某一阶段的发育.不同物种种子萌发的起点温度、适宜温度范围以及有效积温不同[9],与物种的原产地和地理分布密切相关[10],并与有利于随后幼苗生长的环境相匹配[11-12].菊科是被子植物中最大的科,也是入侵植物种数最多的科.一些学者对菊科入侵植物种子生理生态[13]、化感作用[14]、萌发对光的需求[15]等进行了比较研究,揭示了菊科入侵植物种子萌发的生理生态特性.然而,从生物学零度和有效积温角度探讨菊科植物种子萌发对特殊环境条件适应及物种扩张预测的相关研究较少[16-18].
本文以粤东地区6种菊科植物为对象,研究种子萌发的适宜温度范围、生物学零度和有效积温,试图探查6种菊科植物种子萌发过程对环境的适应机制以及扩张潜力,为植物种群动态的研究提供实证资料,为外来植物的入侵机制、预测和防控研究提供相关基础信息.
1 材料与方法
1.1研究地概况
研究地位于粤东地区(114°53′~117°08′E,22°31′~24°15′N),该地域属于亚热带海洋性季风气候,年均降水量1 300~2 400 mm;年日照时数1 900~2 400 h;≥10 ℃的积温7 300~7 890 ℃;年均气温21.4 ℃,6~8月平均气温28~29 ℃,12~2月平均气温9~16 ℃,极端最高气温为39.6 ℃,极端最低气温为-0.5 ℃.
1.2研究材料
所研究植物为菊科(Asteraceae)白酒草属(Conyza)、鬼针草属(Bidens)、金腰箭属(Synedrella)、泽兰属(Eupatorium)、藿香蓟属(Ageratum)、金钮扣属(Spilanthes),每属选取1 种植物,分别为小蓬草(Conyzacanadensis)、鬼针草(Bidenspilosa)、金腰箭(Synedrellanodiflora)、假臭草(Praxelisclematidea)、藿香蓟(Ageratumconyzoides)和金钮扣(Spilanthespaniculata).植物名、原产地及分布区类型[19]见表1.
1.3萌发实验
植物成熟种子于2013年秋季采自广东省潮州市郊区.每种植物50粒种子均匀放在铺有两层滤纸的90 mm培养皿中,在6个温度梯度(15,20,25,30,35,40 ℃)、12 h光照/12 h黑暗条件下进行萌发测试,光照强度为800~1 000 lx,3次重复,以幼根突破种皮、肉眼看到白色幼根为萌发[7],每24 h统计萌发个数,将萌发的种子及时移去,适量补充水分,实验持续30 d.
1.4数据计算与统计
萌发力(Germination ability)表达了种子萌发能力的强弱,通过萌发势和萌发率来体现.萌发势(Germination potential)反映了种子萌发速度和整齐度,是萌发试验初期一定日期内萌发种子数占供试种子数的百分比;萌发率(Germination percentage)是萌发测试终期(30 d)种子萌发数占供试种子数的百分比[17-18].为计算6种菊科植物种子萌发的发育起始温度和有效积温,本实验以累积达到50%萌发率作为完成群体萌发的标准.
有效积温引用公式为
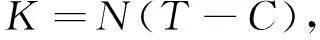
(1)
其中,K为有效积温;T为实验温度;C为生物学零度;N为萌发率达50%时的天数;
生物学零度C的计算:以两种实验温度(T1和T2),分别记录两个相应的发育时间N1和N2值:
(2)
其中1和2代表两个实验.
计算出各实验处理萌发率50%时的天数N,在亚适宜温度范围内采用多组合平均法分别求出每种植物的生物学零度(C)和有效积温(K)[20].
以Excel 和SPSS 19.0统计软件对实验数据进行统计和分析.

表1 6种菊科植物种子萌发的生物学零度和有效积温
*植物原产地年均气温:为原产地各地带最低和最高平均气温范围.北美洲:墨西哥和加拿大;热带美洲:巴西、厄瓜多尔和洪都拉斯;南美洲:阿根廷和巴西.
2 结果与分析
2.16种植物种子萌发的生物学零度与积温
6种菊科植物种子萌发的生物学零度范围是3.94~16.90 ℃,有效积温范围是10.37~65.46 ℃·d.白酒草属小蓬草和鬼针草属鬼针草种子萌发的生物学零度分别为3.94 ℃和5.48 ℃,其有效积温分别为65.46 ℃·d和50.75 ℃·d;金腰箭、假臭草、藿香蓟和金纽扣种子萌发的生物学零度分别为16.90,12.56,11.12和10.69 ℃,其有效积温分别为10.37,46.38,27和25.04 ℃·d(表1).
2.26种植物种子萌发的最适温度范围
6 种植物种子萌发的最适温度范围有较大差异,可分为2类:① 种子萌发的最适温度范围较窄(20~25 ℃)的植物,包括小蓬草和假臭草.小蓬草种子达到最大萌发率(52%~55%)和萌发势(47%~51%)的最适温度范围分别是20~30 ℃和20~25 ℃;假臭草达到最高萌发率(76%)和萌发势(74%)的最适温度范围是20~25 ℃和25 ℃;② 种子萌发的最适温度范围较宽(15~35 ℃或15~40 ℃)的植物,包括鬼针草、金腰箭、藿香蓟和金钮扣.鬼针草达到高萌发率(79%~94%)的温度域为15~35 ℃,高萌发势(65%~86%)的温度域为20~25 ℃;金腰箭达到高萌发率和高萌发势(80%~88%)的温度域为25~40 ℃;藿香蓟达到高萌发率(84%~90%)的温度域为15~30 ℃,高萌发势(73%~88%)的温度域为20~30 ℃;金钮扣达到高萌发率(89%~94%)和萌发势(84%~94%)的温度域为15~40 ℃和20~40 ℃(图1).
2.3种子萌发的生物学零度与积温之间的关系
相关性分析显示,6种植物种子萌发的生物学零度与植物原产地年平均最低气温之间呈显著正相关关系(P<0.05,图2),种子萌发的生物学零度与积温之间为显著负相关关系(P<0.05,图3)小蓬草和鬼针草种子萌发的生物学零度分别为3.94 ℃和5.48 ℃,其有效积温较高(分别为65.46 ℃·d和50.75 ℃·d);金腰箭、藿香蓟和金纽扣种子萌发的生物学零度分别为16.9,11.12和10.69 ℃,其有效积温分别为10.37,27.00和25.04 ℃·d.

图1 6种菊科植物种子萌发的温度范围
3 讨论
本研究发现,6 种植物种子萌发的最适温度范围有较大差异,小蓬草和假臭草种子萌发的最适温度范围为20~25 ℃,鬼针草、藿香蓟和金钮扣种子萌发的最适温度范围分别为15~35 ℃,15~30 ℃,15~40 ℃,金腰箭种子萌发的最适温度范围为25~40 ℃.不同物种种子萌发温度幅的差异可能与物种的原产地、地理分布及其进化适应有密切关系.小蓬草原产北美洲,现广泛分布于我国东北、华北平原、西南和华南地区,其种子萌发的温度幅较窄20~25 ℃,并且只有部分种子萌发(52%~55%),部分种子可能进入种子库,增大物种适合度[21].鬼针草是一种危害极其严重的世界广布种,广泛分布于我国南北各省区的村旁、路边及荒地,达到高萌发率(79%~94%)的温度范围较宽泛,为15~35 ℃,显示鬼针草具有强大的种子繁殖能力和适应环境能力[21-24],因而种群能快速繁殖并具有向高纬度地域扩张的较大潜力.本土植物金钮扣主要分布在我国华南和西南等地,达到高萌发率(89%~94%)的温度域较宽(15~40 ℃),种群也具有向高纬度地带迁移的较大潜力.外来种金腰箭达到高萌发率(80%~88%)的温度域为25~40 ℃,种群的扩张可能受到低温限制,不易向高纬度地带扩展.
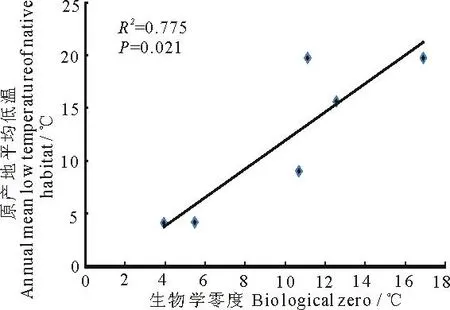
图2 6种植物种子萌发的生物学零度与物种原产地年均低温的相关性

图3 6种植物种子萌发的生物学零度与积温的相关性
研究还发现,6种植物种子萌发的生物学零度与植物原产地年平均最低气温显著正相关、与有效积温显著负相关.产于北美洲的小蓬草和广布种鬼针草种子萌发的生物学零度较低,分别为3.94 ℃和5.48 ℃,其有效积温较高;产于热带美洲和南美洲的金腰箭种子萌发的生物学零度最高(16.90 ℃),其次是假臭草(12.56 ℃)和藿香蓟(11.12 ℃),其有效积温较低.说明适应于低温环境的物种其种子在较低温度下即开始萌发,但只有达到一定总热量后幼苗才能正常建植,减少了多变环境下大量幼苗死亡的风险,增大物种适合度;而适应于温暖环境的物种其种子在较高温度下开始萌发,能够在较短时间内完成萌发,快速建植.种子萌发的生物学零度与植物原产地年均最低气温之间以及与有效积温之间的关系是物种在长期生存中对气候条件适应而逐渐形成的趋势[25-26].
4 结论
1)小蓬草和假臭草达到高萌发率(52%~55%和76%)的最适温度范围为20~25 ℃;鬼针草、藿香蓟和金钮扣达到最高萌发率(80%~95%)的最适温度范围分别为15~35,15~30,15~40 ℃,金腰箭为25~40 ℃,与物种分布区有一定关联.
2)产于北美洲的小蓬草和分布广泛的鬼针草种子萌发的生物学零度较低(分别为3.94和5.48 ℃),产于热带美洲和南美洲的金腰箭种子萌发的生物学零度最高(16.90 ℃),其次是假臭草(12.56 ℃)和藿香蓟(11.12 ℃),与物种原产地有密切关系;6种植物种子萌发的生物学零度与原产地年均低温呈显著正相关,与积温为显著负相关.
3)除金腰箭外,其余5种菊科植物均具有向更高纬度地区扩张的潜力.
[1]闫小玲,刘全儒,寿海洋,等.中国外来入侵植物的等级划分与地理分布格局分析[J].生物多样性,2014,22(5):667.
[2]冯建孟,董晓东,徐成东.中国外来入侵植物物种多样性的空间分布格局及与本土植物之间的关系[J].西南大学学报(自然科学版),2010,32(6):50.
[3]ALPERT P,BONE E,HOZAPFEL C.Invasiveness,invisibility and the role of environmental stress in the spread of non-native plants[J].PerspectivesinPlantEcology,EvolutionandSystematics,2000,3(1):52.
[4]PRENTIS P J,WILSON J R U,DORMONTT E E,et al.Adaptive evolution in invasive species[J].TrendsinPlantScience,2008,13(6):288.
[5]MILLER T.Effects of emergence time on survival and growth in an early old-field plant community[J].Oecologia,1987,72(1):272.
[6]FENNER M,THOMPSON K.TheEcologyofSeeds[M].Cambridge:Cambridge University Press,2005.
[7]BASKIN C C,BASKIN J M.Seeds:Ecology,Biogeography,andEvolutionofDormancyandGermination[M].San Diego:Academic Press,1998.
[8]PROBERT R J.The role of temperature in the regulation of seed dormancy and germination[M]//Seed:TheEcologyofRegenerationinPlantCommunities2nd.CAB International,Wallingford.2000.
[9]STEADMAN K J,PRITCHARD H W.Germination of Aesculus hippocastanum seeds following cold-induced dormancy loss can be described in relation to a temperature dependent reduction in base temperature(Tb) and thermal time[J].NewPhytologist,2004,161(2):415.
[10]TRUDGILLl D L.Why do tropical poikilothermic organisms tend to have higher threshold temperatures for development than temperate ones[J].FunctionalEcology,1995,9(3):136.
[11]ALVARADO V,BRADFORD K J.A hydrothermal time model explains the cardinal temperatures for seed germination[J].PlantCellandEnvironment,2002,25(8):1061.
[12]赵斌斌,张全星,王文慧,等.华北平原11 种植物种子萌发的生物学零点与积温探究[J].中国野生植物资源,2014,33(2):20.
[13]杨逢建,张衷华,王文杰,等.八种菊科外来植物种子形态与生理生化特征的差异[J].生态学报,2007,27(2):442.
[14]李富荣,黄莹,梁士楚,等.几种菊科入侵植物和非入侵植物的化感作用比较[J].生态环境学报,2011,20(5):813.
[15]许慧男,王文杰,于兴洋,等.菊科几种入侵和非入侵植物种子需光发芽特性差异[J].生态学报,2010,30(13):3433.
[16]李丽伟,范芳,张勇,等.菊花地上部干物质增长过程与有效积温的关系[J].中国农学通报,2007,23(7):543.
[17]王梅英,刘文,刘坤,等.青藏高原东缘10种禾本科植物种子萌发的生物学零度和积温[J].草业科学,2011,28(6):983.
[18]刘文,刘坤,张春辉,等.种子萌发的积温效应——以青藏高原东缘的12种菊科植物为例[J].植物生态学报,2011,35(7):751.
[19]吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991,4:1.
[20]云南大学生物系.植物生态学[M].北京:人民教育出版社,1980.75.
[21]肖春萍,刘博,杨莉,等.加拿大蓬种子生态适应性及其化感作用[J].吉林农业大学学报,2011,33(5):527.
[22]郝建华,刘倩倩,强胜.菊科入侵植物三叶鬼针草的繁殖特征及其与入侵性的关系[J].植物学报,2009,44(6):656.
[23]陈文,王桔红,陈丹生,等.五种菊科植物种子萌发对温度的响应及其入侵性[J].生态学杂志,2015,34(2):420.
[24]洪岚,沈浩,杨期和,等.外来入侵植物三叶鬼针草种子萌发与贮藏特性研究[J].武汉植物学研究,2004,22(5):433.
[25]ANGUS J F,Cunningham R B,Moncur M W,et al.Phasic development in field crops:Ⅰ.thermal response in the seedling phase[J].FieldCropsResearch,1981,3(3):365.
[26]TRUDGILL D L,SQUIRE G R,THOMPSON K.A thermal time basis for comparing the germination requirements of some British herbaceous plants[J].NewPhytologist,2000,145(1):107.
(责任编辑俞诗源)
The biological zero and accumulated temperature for seed germination of six Asteraceae species in eastern Guangdong
CHEN Wen
(Department of Geography,Hanshan Normal University,Chaozhou 521041,Guangdong,China)
Temperature is an important environmental factor influencing on seed germination of plants.In this study,we use the thermal time model to analyze accumulated temperature and biological zero for seed germination of six Asteraceae species includingConyzacanadensis,Synedrellanodiflora,Praxelisclematidea,Spilanthespaniculata,Bidenspilosa,Ageratumconyzoidesincubated five temperatures(15,20,25,30,35 and 40 ℃).The result shows that the seed of six Asteraceae species display a 80%~95%germination at the optimum temperature range,exception withCcanadensis(52~55%).The optimum temperature range for germination ofCcanadensisandPclematideais 20~25 ℃,and that of the other species is from 15 to 30 ℃,and to 35 ℃,or to 40 ℃,that ofSnodifloraseed is from 25 to 40 ℃.Biological zero for germination ofCcanadensisnative to north America,andBpilosawidely distribution in world is the lower(3.94 and 5.48 ℃, respectively),and that ofSnodiflora,Pclematidea,Aconyzoidesis higher(16.90,12.56 and 11.12 ℃, respectively).There is a significant positive correlation between biological zero and mean annual low temperature of native habitat of species,and a significant negative correlation between biological zero and accumulated temperature for germination(P<0.05).Exception with S.nodiflora,the population of the other five Asteraceae species may have stronger expansion potential to high latitude zone.
Asteraceae species;seed germination;biological zero;effective accumulated temperature;adaptation
10.16783/j.cnki.nwnuz.2016.04.020
2016-02-06;修改稿收到日期:2016-04-05
国家自然科学基金项目(31470021);广东韩山师范学院科研团队项目(LT201204)
陈文(1963—),男,甘肃张掖人,副教授.主要研究方向为环境生态学.
E-mail:cyw1018@sina.com
Q 985
A
1001-988Ⅹ(2016)04-0093-06
