云烟87大小孢子形成和雌雄配子体发育观察
2016-08-16吉志超李祖任高凯悦张依楠廖海民
吉志超,李祖任,高凯悦,张依楠,廖海民
(贵州大学 生命科学学院,贵州 贵阳550025)
云烟87大小孢子形成和雌雄配子体发育观察
吉志超,李祖任,高凯悦,张依楠,廖海民*
(贵州大学 生命科学学院,贵州 贵阳550025)
采取不同发育阶段烤烟云烟87的花器官,采用常规石蜡切片法对其解剖结构进行研究。结果表明:烤烟云烟87花药壁发育类型为基本型,分化完全的花药壁由表皮、纤维层、中层和绒毡层构成,后者为腺质型绒毡层类型。小孢子母细胞减数分裂类型为同时型,四分体呈四面体型和十字交叉型,成熟花粉粒为二细胞型。烤烟云烟87的胚珠为倒生胚珠,薄珠心,胚珠多数;大孢子母细胞由孢原细胞直接生成,胚囊发育类型为蓼型。
云烟87;大孢子;小孢子;雄配子体;雌配子体
云烟87是以云烟2号(母本)和K326(父本)杂交选育而成,2000年12月经国家品种审定委员会审定[1],该品种结合了其母本和父本的优点,具有抗病性强、品质高、烘烤较易且适宜广泛种植等特点[2]。根据贵州省烟草公司数据资料,2015年贵州省烤烟主栽品种主要有四种:云烟87、K326、云烟85、南江3号,其中云烟87占40%,约80万亩。目前,对云烟87的研究主要集中在栽培、烘烤和品种特征等方面[1-6],廖菊够等[7]对迪勃纳氏烟草大小孢子发生及雌雄配子发育过程进行观察研究,尚未见有关云烟87的胚胎学方面的研究报道。
大小孢子形成和雌雄配子体发育是植物进行生殖繁育的重要阶段,研究云烟87的配子体发育规律既可以了解其生殖生长进程,同时也为研究烟草杂交育种提供理论基础。运用石蜡切片技术对云烟87大小孢子的发生和雌雄配子体的发育进行了研究,以期了解该物种的生殖和繁衍特性,研究结果将为控制花期,制定有效的田间栽培措施,以及烟草种质资源的保持和开发利用提供理论依据。
1 材料与方法
1.1实验材料
烤烟云烟87,来源为清镇市烟草公司现代烟草农业芦荻基地单元。取云烟87处在盛花期大小不同的花蕾,将雌蕊和雄蕊分离出,置于50%FAA固定液中保存备用。
1.2实验方法
本实验采用石蜡切片法[8]。经过乙醇浓度递增脱水、二甲苯透明、浸蜡、包埋、切片(厚度8~10 μm)、番红-固绿染色、中性树胶封片等过程。再用Olympus DP26显微镜观察并拍照。
2 结果与分析
2.1小孢子发生和雄配子体的形成
烤烟云烟87的花为两性花,每朵花中包含5个雄蕊,每个花药具4个花粉囊(图1-1)。小孢子发生起始于雄蕊原基,由一群具有旺盛分裂能力的细胞构成。雄蕊原基细胞不断分裂逐渐呈四棱形,在其四个角隅处分化产生核大、质浓的孢原细胞(图1-2),孢原细胞进行平周分裂形成初生壁细胞和初生造孢细胞,紧贴表皮为初生壁细胞,其内方为初生造孢细胞(图1-3)。
初生造孢细胞经多次有丝分裂形成小孢子母细胞,小孢子母细胞呈多边形、体积较大、细胞核较为明显(图1-4)。小孢子母细胞经减数分裂形成小孢子,减数分裂的第一次分裂不产生细胞壁,其胞质分裂类型为同时型(图1-6)。第二次分裂产生四个子细胞,在四个子核之间产生细胞壁,细胞质彼此分离。形成的4个小孢子聚集在一起,为四分体时期,四分体呈四面体型和十字交叉型,其中四面体型较多(图1-7)。以后,四分体的胼胝质壁溶解,分离为四个小孢子(图1-8)。刚被释放出小孢子形状不规则,体积较小,着色比较浅。之后小孢子的体积逐渐增大,呈圆形,表面具3个萌发沟(图1-9)。刚开始小孢子早期细胞核位于细胞中央,为单核中央期(图1-9),随后由于中央大液泡形成,液泡把细胞核挤到了紧贴细胞壁的一侧(图1-10)。之后小孢子细胞发生一次不对称的有丝分裂,进入双核期,位于花粉粒中央的核为营养核,位于花粉粒贴近细胞壁位置的核为生殖核,因此烤烟云烟87的花粉粒属于二细胞型花粉(图1-11)。
初生壁细胞继续进行多次垂周分裂和平周分裂形成多层细胞,这些细胞和位于最外层的表皮构成花药壁。早期的花药壁由外向内依次是:表皮、药室内壁、中层、绒毡层(图1-5)。
表皮位于最外层,为一层排列紧密的细胞构成,其外壁上具有角质膜,具有保护功能(图1-5)。药室内壁紧贴表皮,在中层之外,细胞较大,为一层细胞(图1-5)。在花药成熟时,药室内壁细胞的内切向壁和径向壁出现加厚,分化成纤维层(图1-11)。在花粉囊的连接处则不形成纤维层,仍保持薄壁状态,以后此处开裂,花粉粒从此散出(图1-12)。中层位于绒毡层和药室内壁之间,由2层细胞构成,中层在小孢子发育过程中会逐渐解体(图1-5)。位于花药壁最内层的细胞为绒毡层,由一层体积较大且质浓厚的细胞构成。绒毡层的细胞具有分泌功能,对小孢子的发生有重要的作用。早期的绒毡层细胞是单核的,小孢子母细胞减数分裂时,绒毡层细胞活动开始活跃,细胞由单核变成两核或多核(图1-7),当减数分裂快结束时,也就是快到小孢子时期时,绒毡层开始出现退化(图1-9),在小孢子发育的晚期,也就是花粉成熟时,绒毡层完全退化(图1-12)。小孢子发生进程中,绒毡层细胞的原生质一直在其原位置,到花粉成熟时,细胞壁解体,细胞自溶,属于腺质绒毡层类型。
2.2大孢子发生和雌配子的形成
烤烟云烟87子房由2个心皮合生成,2室,中轴胎座,倒生胚珠,多数胚珠(图2-1,2)。在胚珠的珠孔端,表皮下面的一个细胞分化形成孢原细胞,其细胞的体积比周围细胞大,质浓厚,核明显,这个细胞不经过分裂直接成为大孢子母细胞(图2-3,4)。大孢子母细胞在进行减数分裂形成大孢子前,细胞体积变大,细胞周围被胼胝质包围。
当大孢子母细胞体积增大到一定阶段,即进行减数分裂,减数第一次分裂形成二分体(图2-5),二分体继续分裂,经减数第二次分裂形成四分体,四分体呈一字线排列(图2-6,7)。4个大孢子中有3个退化,近合点端大孢子继续发育为功能大孢子,此种胚囊发育类型为单孢子胚囊(图2-8)。
功能大孢子的细胞核发生连续的3次有丝分裂。第一次有丝分裂形成二核胚囊(图2-9),二核胚囊体积进一步增大,两个细胞核分别移向珠孔端和合点端,第二次有丝分裂形成四核胚囊(图2-10),第三次有丝分裂形成八核胚囊,此时在大孢子母细胞中的珠孔端和合点端分别具有四个核,之后这两端分别有一个核向胚囊中央移动并相互靠拢,构成含有两个核的中央细胞,剩下合点端的3个核形成3个反足细胞,珠孔端的3个细胞核形成3个卵器,包括2个助细胞和1个卵细胞,其中助细胞靠近珠孔,卵细胞靠近中央细胞。至此,一个含有7细胞8核的成熟胚囊形成(图2-11,12)。
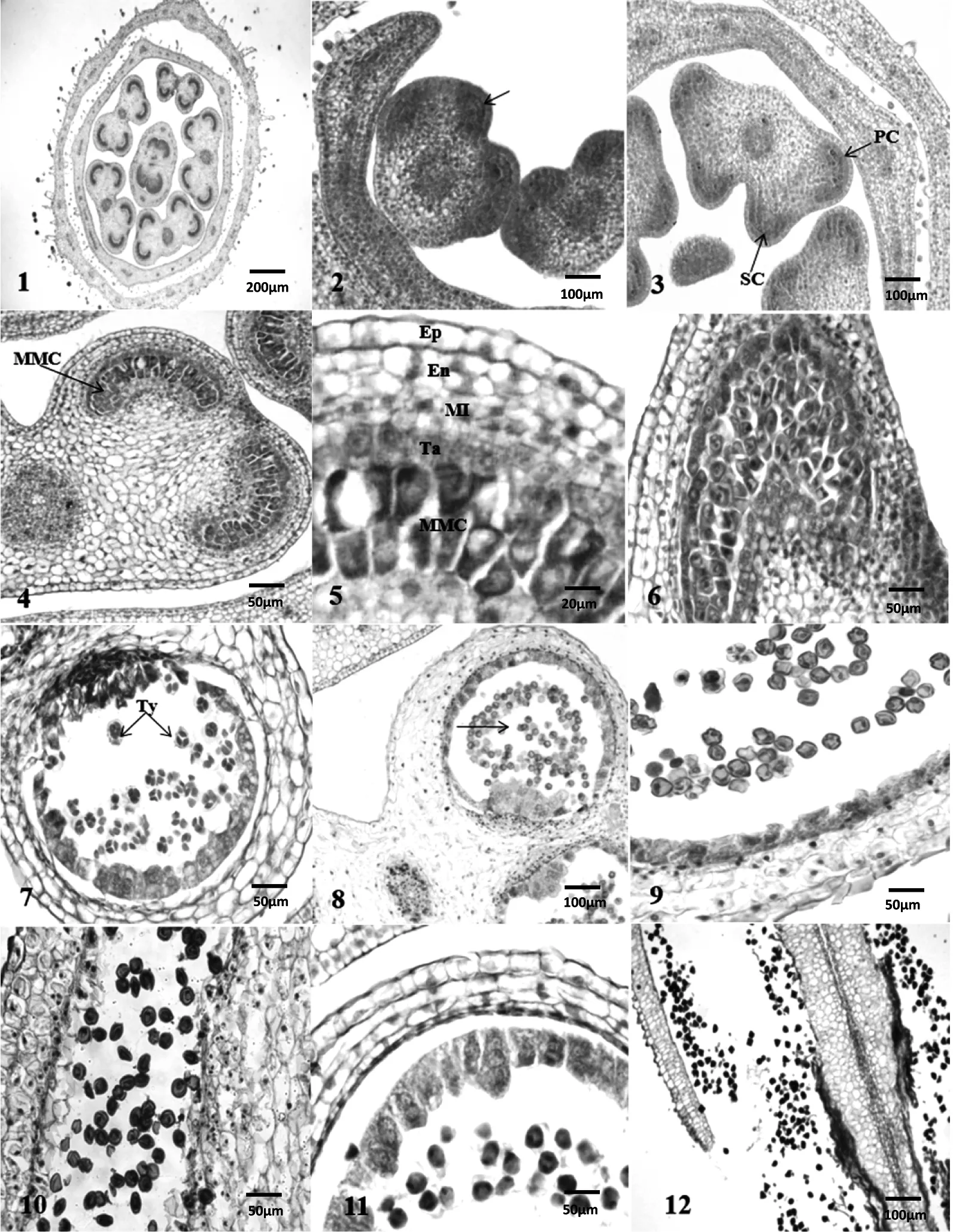
图1 云烟87小孢子发生和雄配子体发育
1.花横切;2.孢原细胞出现,箭头示孢原细胞;3.花药横切,出现初生壁细胞和初生造孢细胞;4.小孢子母细胞时期;5.分化完全的花药壁;6.小孢子母细胞减数分裂时期;7.四分体时期;8.四分体分离为四个小孢子,箭头示四分体刚分离;9.小孢子具3个萌发沟;10.双核期初始;11.二细胞型花粉;12.成熟花粉时期
PC.周缘细胞;SC.造孢细胞;MMC.小孢子母细胞;Ep.表皮;En.药室内壁;MI.中层;Ta.绒毡层细胞;Ty.四分体

图2 云烟87大孢子发生和雌配子体的形成
1.子房横切;2.心皮纵切;3.孢原细胞时期,箭头示孢原细胞;4.大孢子母细胞;5.二分体时期,箭头示二分体;6.四分体早期,箭头示四分体呈一字线排列;7.四分体晚期,箭头示近合点端一个大孢子与其余三个大孢子分离;8.功能大孢子;9.二核胚囊;10.四核胚囊,箭头示合点端和珠孔端分别具有两个核;11.成熟胚囊,箭头示卵器;12.成熟胚囊,示反足细胞、极核
Me.大孢子母细胞;OI.株被;EC.卵细胞;Sy.助细胞;PN.极核;AC.反足细胞
3 结论与讨论
云烟87在小孢子发生过程中,小孢子母细胞减数分裂类型为同时型[9]。根据Davis[10]对被子植物花药壁发育类型的划分,云烟87的花药壁发育属于基本类型。在云烟87雄配子体形成过程中,小孢子依次经历的过程为:四分体、单核中央期、单核靠边期、双核期,分化成熟的花粉为二细胞型花粉。成熟花粉粒的生殖细胞在花粉管中分裂形成2个精子, 这种花粉类型约占被子植物1/3[11]。 这与全雪丽[12]等对烟草吉烟9号品种小孢子的发育研究基本一致。这为烟草杂交育种及云烟87的单倍体育种研究提供依据。
云烟87的大孢子母细胞由珠心表皮下的孢原细胞直接形成,属于薄珠心类型[9]。大孢子母细胞为单孢子胚囊型,70%被子植物的胚囊为该类型[13]。在雌配子体的形成过程中,单孢子胚囊中的未退化的大孢子发育为功能大孢子,功能大孢子经过3次连续的有丝分裂,形成7细胞8核胚囊,因此胚囊发育属于蓼型胚囊。符合对茄科植物胚胎学特征的描述,只是与辣椒的7细胞7核胚囊[14]不同。通过对比云烟87大小孢子的发育时间,发现当雄蕊发育为四分体时,雌蕊开始发育。与廖菊够[15]等对迪勃纳氏烟草描述小孢子发育早于大孢子有所不同。
[1] 李永平,王颖宽,马文广,等. 烤烟新品种云烟87的选育及特征特性[J]. 中国烟草科学,2001(4): 38-42.
[2] 张丽,姬厚伟,黄锡娟,等. 贵州烟区烤烟品种云烟87主要化学成分特征分析[J]. 安徽农业科学,2015,43(20): 288-290,399.
[3] 贾宏昉,尹贵宁,黄化刚,等. 低磷胁迫对云烟87糖代谢及营养元素吸收的影响机理初探[J]. 中国农业科技导报,2014,16,(3): 36-41.
[4] 范才银,曾惠宇,林志,等. K326和云烟87在湘南烟区农艺特性及产量差异性研究[J]. 湖南农业科学,2012,(13): 33-35.
[5] 丁雪丹,肖玉,周紫燕,等. 干旱对云烟87和K326的农艺性状影响研究[J]. 江西农业大学学报,2012,34(5): 893-898.
[6] 龚婷,查宏波,魏世强,等. 不同光质处理对云烟87生长及品质的影响[J]. 西南大学学报,2013,35(1): 29-34.
[7] 廖菊够,康洪梅,代瑾然,等. 烟草大小孢子发生和雌雄配子体发育研究[J]. 西北植物学报,2013,33(1): 11-16.
[8] 李祖仁,胡楠,杨吉刚,等. 繁缕和鹅肠菜的花维管系统比较解剖学研究及其系统学意义[J]. 植物科学学报,2013,31(6): 525-532.
[9] 胡适宜. 被子植物胚胎学[M]. 北京: 高等教育出版社,1985: 75.
[10] Davis G L. Systematic embryo logy of the angiosperms john wiley and sons[M]. Inc. New York,London: Sydney,1966:267-268.
[11] 胡适宜,杨弘远. 被子植物受精生物学[M]. 北京科学出版社,2002: 54-60.
[12] 全雪丽,李虎林,具红光,等. 烟草吉烟9号品种小孢子发育时期与花器官形态的相关性[J]. 烟草科技,2014,(10):75-78.
[13] 胡正海. 植物解剖学[M]. 北京: 高等教育出版社,2010: 289-294.
[14] 申家恒,李伟,杨虹,等. 辣椒大小孢子发生与雌雄配子体发育的研究[J]. 园艺学报,2007,34(6): 1443-1452.
[15] 廖菊够,康洪梅,代瑾然,等. 烟草大小孢子发生和雌雄配子体发育研究[J]. 西北植物学报,2013,33(1): 11-16.
The Megasporogenesis and Microsporogensis of Flue-cured Tobaccovar. Yunyan87 and Its Male and Female Gametophyte Formation
JIZhi-chao,LIZu-ren,GAOKai-yue,ZHANGYi-nan,LIAOHai-min*
(CollegeofLifeSciences,GuizhouUniversity,Guiyang,Guizhou550025,China)
Anatomical structure of the floral organ of Flue-cured tobacco var. Yunyan 87 at different development stage was studied by common plant paraffin section. The results showed that,the anther wall wasdefined as basis development type,which contained glandulartapetummiddle layer,endotheca and epidermis. The meiosis of microspore mother cells was the simultaneous type,the tetrad arranged as tetrahedral or decussate tetrad. The mature pollens belong to 2-cell type.The ovule of Flue-cured tobacco var. Yunyan 87 wasanatropous,thin nucellus and numerous. The megaspore mother cell wasdeveloped from the archesporial cell and the megaspore at the chalazal end developed into the polygonum type embryosac.
Yunyan87;Megaspore;Microspore;Femalegametophyte;Male gametophyte
1008-0457(2016)03-0039-05国际DOI编码:10.15958/j.cnki.sdnyswxb.2016.03.007
2016-03-04;修回日期:2016-04-08
贵阳市现代农业与农村科技计划(2013筑科合同[2013102]4-6号)。
廖海民(1965-),男,博士,教授,主要研究方向:植物分类及形态解剖学研究;E-mail: lhaimin@163.com。
Q944.5
A
