杏的自交不亲和机制研究进展
2016-08-10韩雪平王金政薛晓敏王来平
韩雪平,王金政,薛晓敏,王来平
(山东省果树研究所,山东泰安,271000)
杏的自交不亲和机制研究进展
韩雪平,王金政,薛晓敏*,王来平
(山东省果树研究所,山东泰安,271000)
摘要:被子植物的自交不亲和有孢子体型不亲和与配子体型不亲和两种类型,杏的自交不亲和属于配子体型不亲和。从生物学、细胞学和分子学角度综述杏的自交不亲和机制,对比分析研究自交亲和品种和自交不亲和品种不同的受精过程及杏的开花生物学和授粉生物学特性,为生产上花期管理、合理选配授粉树提供依据,为杏种质资源、遗传育种及生物技术等方面研究提供参考。
关键词:杏;自交不亲和;配子体自交不亲和
Kostina (1964)[1]将栽培杏(AmeniacavulgarisLam.)的700个品种划分为4个地理生态群,即中亚生态群、伊朗-高加索生态群、欧洲生态群、准噶尔-外伊犁生态群,其中欧洲生态群是杏演化历史进程中最为年轻的一个地理生态群,大多品种表现自交亲和(Self-compatibility,简称SC),华北和准噶尔-外伊犁中国杏的地理生态群品种、类型或株系总体上表现为自交不亲和[2](Self-incompatibility,简称SI。自交不亲和性可分为两大类型,一类是孢子体型SI(Sporophytic self-incompatibility,SSI),一类为配子体型SI(Gametophytic self-incompatibility,GSI)。主要是由于花粉的S基因表达时期不同[3]。中国杏自交不亲和属于配子体自交不亲和,因雌雄性器官具有相同的S等位基因,它们相互排斥,交配不亲和。孢子体自交不亲和受S位点复等位基因控制,当自花授粉时,SP11/SCR与SRK(S位点蛋白11、S位点富半胱氨酸蛋白、S位点受体激酶)特异识别,造成后者的Ser/Thr激酶的磷酸化,引发了一系列由SLG、ARC1(S位点糖蛋白、孢子体自交不亲和反应臂重复蛋白)及水孔蛋白等因子参与的SSI信号转导途径,最终产生自交不亲和[4]。
1配子体与孢子体自交不亲和机制
自交不亲和植物拒斥自我花粉和与其基因型相同的花粉,依它们的表现方式可划分为3种类型,即花粉表型的遗传控制,花的形态和所涉及遗传位点的数量。根据花粉不亲和表型遗传控制方式的不同,单因子SI分为两类,一类为配子体自交不亲和性(gametophytic self-incompatibility, GSI),花粉不亲和表型由单倍体基因型决定,配子体型的S基因在减数分裂后才有活性,这类植物自交不亲和表型取决于自身的单倍体S基因型。花粉管生长的抑制常发生在花柱传递组织中。另一类为孢子体自交不亲和性( Sporophytic self-incompatibility, SSI),即花粉不亲和表型由孢子体基因型决定,孢子体型自交不亲和性在十字花科、旋花科和菊科中发现,如油菜、白菜属于这一类。一般认为,孢子体的S基因在花药中表达,其基因产物转移到花粉上发挥生物功能,因此花粉的自交不亲和取决于花粉的二倍体亲本植物即孢子体细胞核内两个S等位基因的相互作用。配子体型自交不亲和和孢子体型自交不亲和的作用方式,配子体花粉生长抑制在花柱传递组织中,孢子体花粉停止生长在柱头表面[5](图1)。
配子体型自交不亲和遗传机制相当复杂。GSI 分子水平研究的开端是Lewis在月见草(Oenotheraorganesis)中分离了与特定S等位基因相关的蛋白质,因此,研究S-蛋白是如何识别自花和异花花粉的假说,主要有单基因和双基因控制 SI 的两类假说,但具体区分的分子机理还不清楚。单基因假说认为,S 位点上的一个基因同时控制花柱和花粉的自交不亲和性。双基因假说认为,花粉和雌蕊中分别存在不同S基因控制着 SI。这类假说以S-RNase的细胞毒素效应为基础,并又可分为两种模型,即膜受体学说和胞内抑制剂说[6]。膜受体学说是花粉S-基因产生的膜受体特与S-RNase特异性的结合。但这种模型很难解释S-位点基因片断重复的花粉自交亲和的现象和茄科的二倍体加倍。胞内抑制剂说认为,花粉S-基因编码一种特异的胞内RNase抑制剂。这种抑制剂抑制除自己的S-等位基因编码的S-RNase外的其它各种S-RNase 的活性。这种假说可以成功地解释为什么二倍体花粉是自交亲和的。体外核酸酶实验证明,花粉管吸收S-RNase没有表现出特异性[7-8]。运用花粉管的免疫组织化学标记方法证明了亲和与不亲和的花粉管的细胞质中都有 S-RNase 的积累。这为抑制剂假说提供了充分的实验证据,但是这种抑制剂为何不对自体核酸酶发生抑制的机制却很难让人理解。
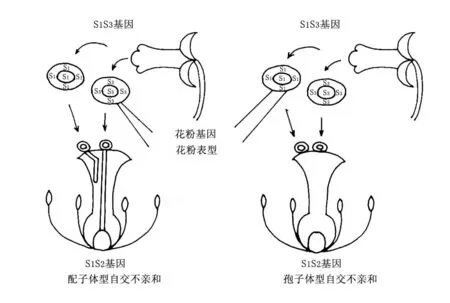
图1配子体型和孢子体型自交不亲和的作用方式
2生物学基础
果树花粉量的多少和花粉萌发率的高低直接影响其授粉、受精及坐果。因此,无论引种栽培,还是杂交育种工作,开展各个品种花粉量和生活力的研究都具有重要意义[9]。比较研究发现杏品种间花粉萌发率的差异很大,但是所有品种均为花粉可育型,并且大多数品种不完全花百分率很高,但是不完全花的多少是品种特性,受遗传因素的支配[10],因此综合考虑自然授粉结实率很低的主要原因是雌蕊败育。品种间不完全花百分率与花粉萌发率呈正相关[11]。但是,王朝凤研究发现,花粉管生长长度比花粉萌发率更能代表花粉的代谢活力,与能否完成受精关系更为密切[12],所以以花粉量和花粉管长度聚类分析的结果作为花粉萌发特性好坏的依据。
刘宝敬等[13]对孢子体不亲和甘蓝进行的研究结果表明,自交不亲和系与亲和系的柱头和花粉氨基酸组分有显著差异,柱头中苏氨酸以及花粉中甘氨酸含量可以作为评价自交不亲和性的指标。郑洲的实验证明,花粉氨基酸组分在自交亲和品种系与自交不亲和品种系上无显著差异,而花柱氨基酸的组分与花粉氨基酸组分有所不同,其中自交不亲和品种系酪氨酸、苏氨酸、甲硫氨酸和精氨酸含量明显高于自交亲和品种,含量的水平达到了差异显著或极显著。其原因可能与配子体不亲和与孢子体不亲和的花粉与花柱的识别不同有关。孢子体不亲和性的识别部位主要在花粉和柱头,而配子体不亲和性的识别部位主要在花柱[14]。
3细胞学基础
冯建荣和郑洲都认为,无论是自交亲和还是自交不亲和的品种,授粉24小时时,花粉在柱头上萌发,二者并无明显差异,48小时时,自交亲和品种已有花粉管进入子房, 而自交不亲和品种花粉管多数生长至花柱近中部,未发现有进入子房的。72小时观察, 可以看到二者有明显差异,自交亲和品种的花柱基部有大量花粉管,许多花粉管已经延伸进入子房上端, 而自交不亲和品种的花柱基部花粉管较少,只有极个别花粉管进入子房, 多数的花粉管则在花柱中下部停止生长,同时有顶端膨大现象,并在授粉后96小时进入子房[15]。彭晓莉研究72个授粉组合的花粉管伸长荧光显微观察结果看出,大多数花粉在授粉24小时后开始萌发,只有其中的6个组合授粉后96小时到达花柱基部,其余的均与郑洲、冯建荣的研究结果一致。因此认为花粉萌发率和花粉管达到花柱基部的多是花粉活力较高的品种。授粉组合中坐果率较高的授粉品种,花粉萌发率高、花粉管到达花柱基部的速度快,比率大。所以选择花粉活力高、与母本亲和性强的父本是提高坐果率的关键因素[16]。
4分子学基础
一般情况下,配置授粉品种时考虑的首要因素是杂交亲和力,一方面是因为杂交花朵坐果率30%以上才能够满足丰产栽培的要求,另一方面因为杂交坐果率数据还可为DNA分子水平上确定品种的S基因型提供依据。杂交结实率≤30%,一般认为不亲和,S基因型有可能相同;若品种间杂交坐果率超过30%,说明两个S等位基因中至少一个S等位基因不同[17]。
当将SI等位基因通过杂交引入自交亲和(Self-compatibility,SC)植株时,常发现还有其它因素影响SI反应,这在早期和现在的实验中都有证实[18-19]。Berantzky 等更进一步提出了遗传证据,证明仅有 S-RNase 不足以决定雌蕊S表型。将S-RNase转入同种SC植株并未使其获得S基因特异性花粉拒斥作用,如巴旦水杏的自交不亲和就是由于花柱中的S-核酸酶(S-RNase)进入花粉管细胞器,导致自交授粉的花粉管的mRNA和rRNA降解,使细胞溶解,导致花粉管生长受到抑制[20]。然而当S-RNase在SI和SC植株杂交的F1代表达时,却可拒斥对应的S基因型花粉,显然存在非S-RNase参与柱头中的SI反应。自交亲和Nicotiana sylvestris 品种中残存的S-RNase 似乎也与此有关[21]。通过差异性cDNA克隆的方法从烟草花柱中分离到一个 SI 相关因子即天冬氨酸丰富蛋白HT,反义转基因实验证明,降低该蛋白表达可使植株接受原本不亲和的S基因型花粉(McClure et al,1999)[22]。凯特和新世纪品种的3个S-RNase 基因DNA、RNA(cDNA)片段序列显示出P.aS8、P.aS9、P.aS10 在C1-C5 保守区间, 内含子长度、碱基排列顺序均有差异, 且在高变区(RHV) 和保守区(RC4)也有明显的差异。据已有研究认为S-RNase 基因的高变区(RHV)在自交不亲和反应中可能参与花粉S-蛋白的专一性识别[23-25],而位于高变区内的内含子虽然没有编码形成S-RNase基因的氨基酸, 但是在雌蕊与花粉S-基因的识别及亲和反应中是否为多余的“垃圾序列”,或者具有某些尚未阐明的功能[26]。HT的功能还有待进一步证实,可能与S-RNase形成复合物,或者与S-RNase进入花粉管有关。
5小结与展望
分别从生物学、细胞学和分子学角度阐述杏的自交不亲和机制,花粉管的生长长度比花粉萌发率和完全花的比例更能代表自花坐果率的高低,从冯建荣、郑洲、彭晓莉试验研究证明,SC和SI的花粉管在花柱中生长的荧光显微镜观察48小时时已出现差异,72小时时差异明显,SC的花粉管已进入子房上端,而SI则是在花柱中下部停止生长;杏花柱S-RNase 的活性与杏自交不亲和性基因有关,S-RNase 可能是S 基因的产物,其中SC的花柱S-RNase 也出现了两个带[27];对杏还确定了S1~S23 的23个S 基因及70个品种的S 基因型[28]。研究杏的自交不亲和机制为欧洲杏中的自交亲和品种和中国杏中的自交不亲和品种的杂交育种工作提供理论基础。
参考文献:
[1]Kost ina K F. 1964. Application of phytogeographicalm ethod to apricot classification. TrudNik it Bot Sad. 37, Kolos, Moscow.
[2]何天明,陈学森,张大海,等.中国普通杏种质资源若干生物学性状的频度分布[J].园艺学报, 2007,34 (1):17-22.
[3]王爱华,戴洪义.果树自交不亲和性的研究进展[J].莱阳农学院学报, 2002,19(3): 202-209.
[4]赵永斌,朱利泉,王小佳.植物孢子体自交不亲和信号转导功能分子研究进展[J].生物技术通讯,2004,02:176-178.
[5]Kao T-h and Huang S.Gametophytic self-incompatibility: A Mechanism for self/ non-self discrimination during sexual reproduction[J]. Plant Physiol, 1994, 105: 461-466.
[6]Kao TH, McCubbin AG. How flowering plants discriminate between self and non-self pollen to prevent inbreeding[C]. Proc Nat Acad Sci USA, 1996, 93: 12059-12065.
[7]Gray JE, Glavin TL, Schafer U, et al. Action of the style product of theself-incompatility gene Nicotiana alata (S-RNase) on in vitro grown pollen tubes [J]. Plant Cell, 1991, 3: 271-286.
[8]Luu DT, Qin X, Morse D et al. S-RNase uptake by compatible pollen tubes in gametophytic self-incompatibility [J]. Nature, 2000,407:649-651.
[9]叶正文,杜纪红,苏明申,等.桃92个品种的花粉量及其萌发特性的差异[J].园艺学报,2010, 37(4):525-531.
[10]吕增仁,潘哲伟,尹铁民,等.若干杏品种的授粉生物学特性研究[J].园艺学报,1992,01:7-10.
[11]顾景梅,史修柱,李军祥.限制杏树丰产的因素及对策[J].落叶果树, 1999,(1): 51~52.
[12]王朝凤,杨途熙,魏安智,等.杏不同品种花粉量及花粉萌发特性的研究[J].北方园艺,2012, 09:1-5.
[13]刘宝敬,宋明,李成琼,等.利用氨基酸分析快速测定甘蓝自交不亲和性植物学报,1998,40(11):1028-1034.
[14]郑洲,陈学森,冯宝春,等.杏品种授粉生物学研究[J].果树学报,2004,04:324-327+395.
[15]冯建荣,陈学森,孔宁,等.杏(Prunusarmeniaca)自交不亲和强度及其授粉受精相关特性[J].果树学报,2006,05:690-694+786.
[16]彭晓莉,廖康,贾杨,等. 9个新疆杏品种间杂交亲和性研究[J].果树学报,2014,12.
[17]Hidenori Sassa , Hisashi Hirano , Hiroshi Ikehashi ,Self-incompatibility-related RNase in Styles of Japanese Pear (PyrusserotinaRehd)[J].Plant and cellPhysiol , 1992 , 33 :811-814.
[18]McClure B, Mou B, Canevascini S et al. A small asparagines-rich protein required for S-allele-specific pollen rejection in Nicotiana [J].Proc Natl Acad Sci USA, 1999, 96:13548-13553.
[19]Ai Y, Kron E, and KaoT-h. S-alleles are retained and expressed in a self-compatible cultivar of Petunia hybrida [J]. Mol. Gen. Genet. 1991,230: 353-358.
[20]齐洁.杏自交不亲和相关基因的克隆及表达分析[D].山东泰安:山东农业大学,2002.
[21]Golz JF, Clarke AE, Newbigin E, et al. Arelic S-RNase is expressed in the styles of self-compatible Nicotiana sylvestris [J]. Plant J, 1998, 16:591-599.
[22]McClure B, Mou B, Canevascini S et al. A small asparagines-rich protein required for S-allele-specific pollen rejection in Nicotiana [J].Proc Natl Acad Sci USA, 1999, 96:13548-13553.
[23]Sassa H, Nishio T, Kowyama Y, et al.1996.Self-incompatibility (S) alleles of the Rosaceae encode members of a distinct class of the T2PS ribonuclease super family[J]. Mol Gen Genet, 50: 547- 557.
[24]Sonneveld T,Robbins T P, Boskovi c R, et al. 2001.Cloning of six cherry self-incompatibility alleles and development of allele-specific PCR detection[J]. Theor ApplGenet, 102: 1046-1055.
[25]Wunsch A,Hormaz J I. 2004. Cloning and characterization of genomic DNA sequences of four self-incompatibility alleles in sweet cherry ( Prunus avium L. ) [J].TheorAppl Genet, 108: 299- 305.
[26]冯建荣,陈学森,吴燕.‘凯特’与‘新世纪’杏自交不亲和S-RNase基因的检测及克隆[J].林业科学,2006,10:129-132.
[27]Burgos L, Perez-Tornero O, Ballester J, Olmos E, 1998. Detection and inheritance of stylar ribonuleases associated with incompatibility alleles in apricot[J]. Sex. Plant Reprod, 1: 153-158.
[28]张绍铃,曹生民,吴华清.果树自交不亲和性基因型及其鉴定方法[J].果树学报, 2003,20:358-363.
收稿日期:2016-01-13
基金项目:国家“十二五”科技支撑计划资助项目(2013BAD02B03-3-3),山东省农业良种工程。
*通讯作者:薛晓敏( 1979- ),女,河北邯郸人,助理研究员,主要从事水果育种与栽培等研究工作。
作者简介:韩雪平(1985- ),女,山东菏泽人,研究实习员,主要从事核果类的育种工作。Email:hanxuepingrun@163.com
中图分类号:S662.2
文献标识码:A
文章编号:1002-2910(2016)04-0014-04
